
Neuroprotective Potential of Guiera senegalensis (Combretaceae) Leaf Hydroethanolic Extract against Cholinergic System Dysfunctions and Oxidative Stress in Scopolamine-Induced Cognitive Impairment in Zebrafish (Danio rerio)

 ,
,  ,
,  ,
, 
 ,
, 
Abstract
1. Introduction
2. Results and Discussion
2.1. Chemical Composition of Guiera senegalensis Extract
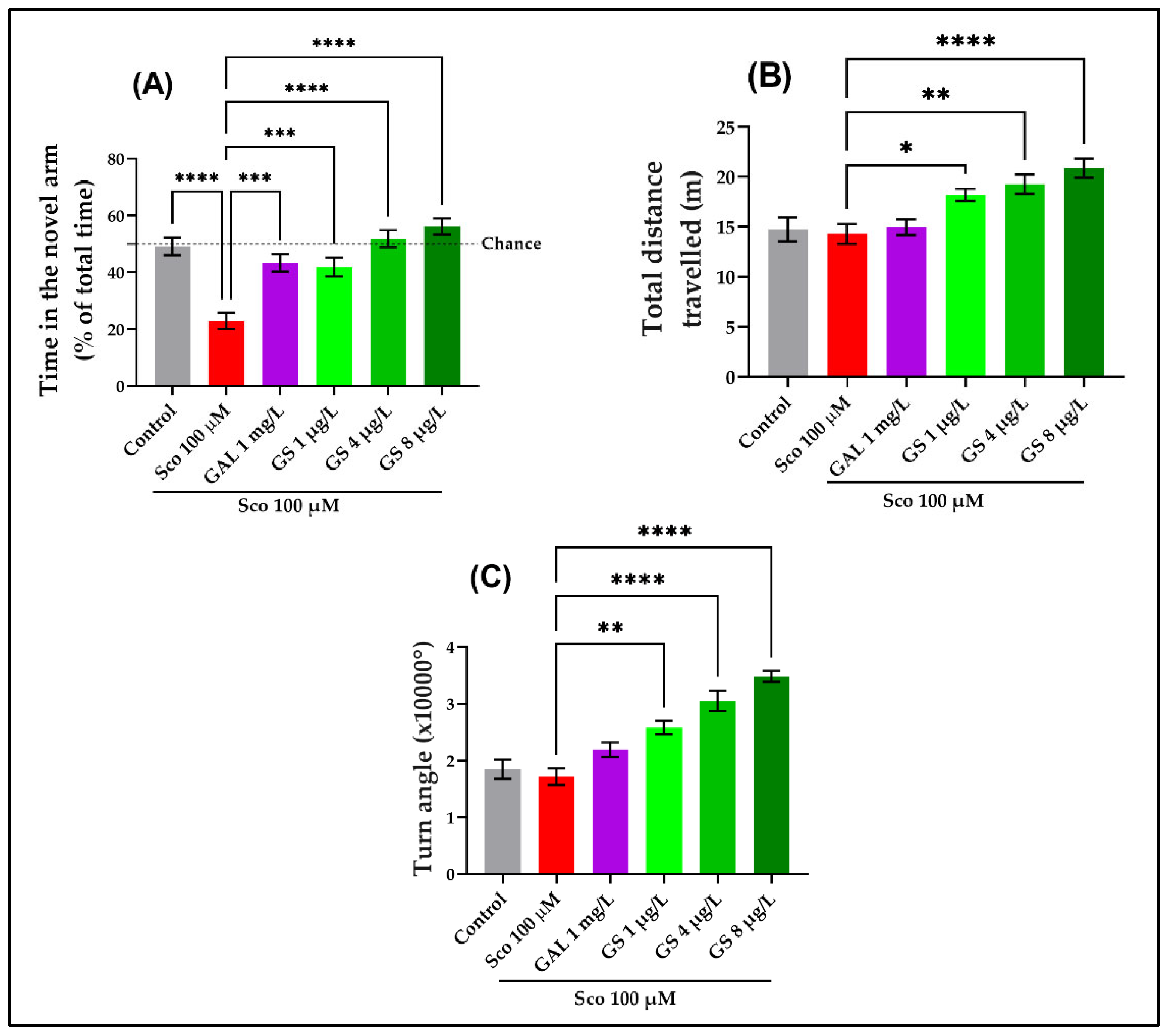
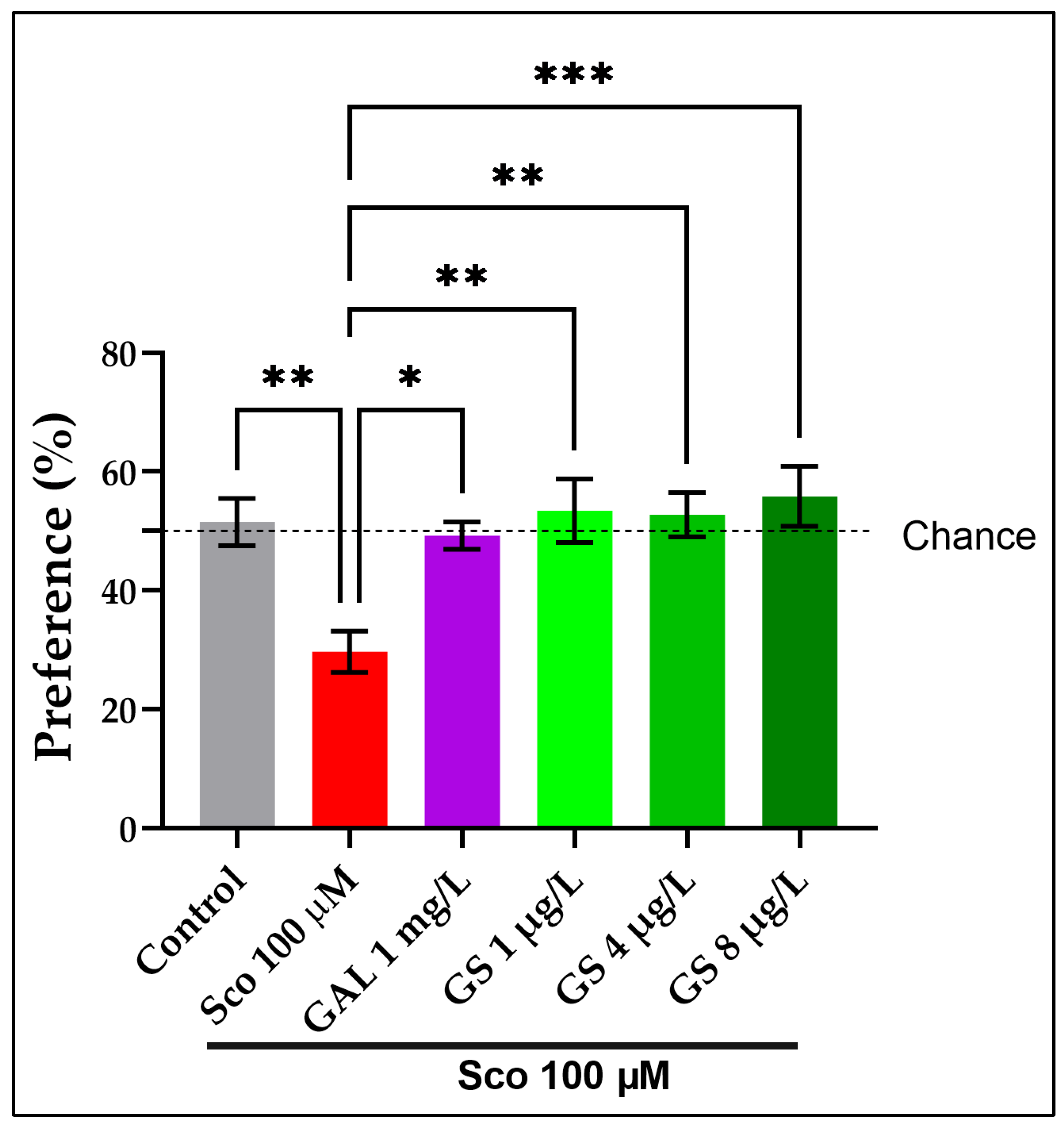
2.2. Effects on Anxious Behavior and Spatial Memory in Zebrafish in NTT, Y-Maze and NOR Tests
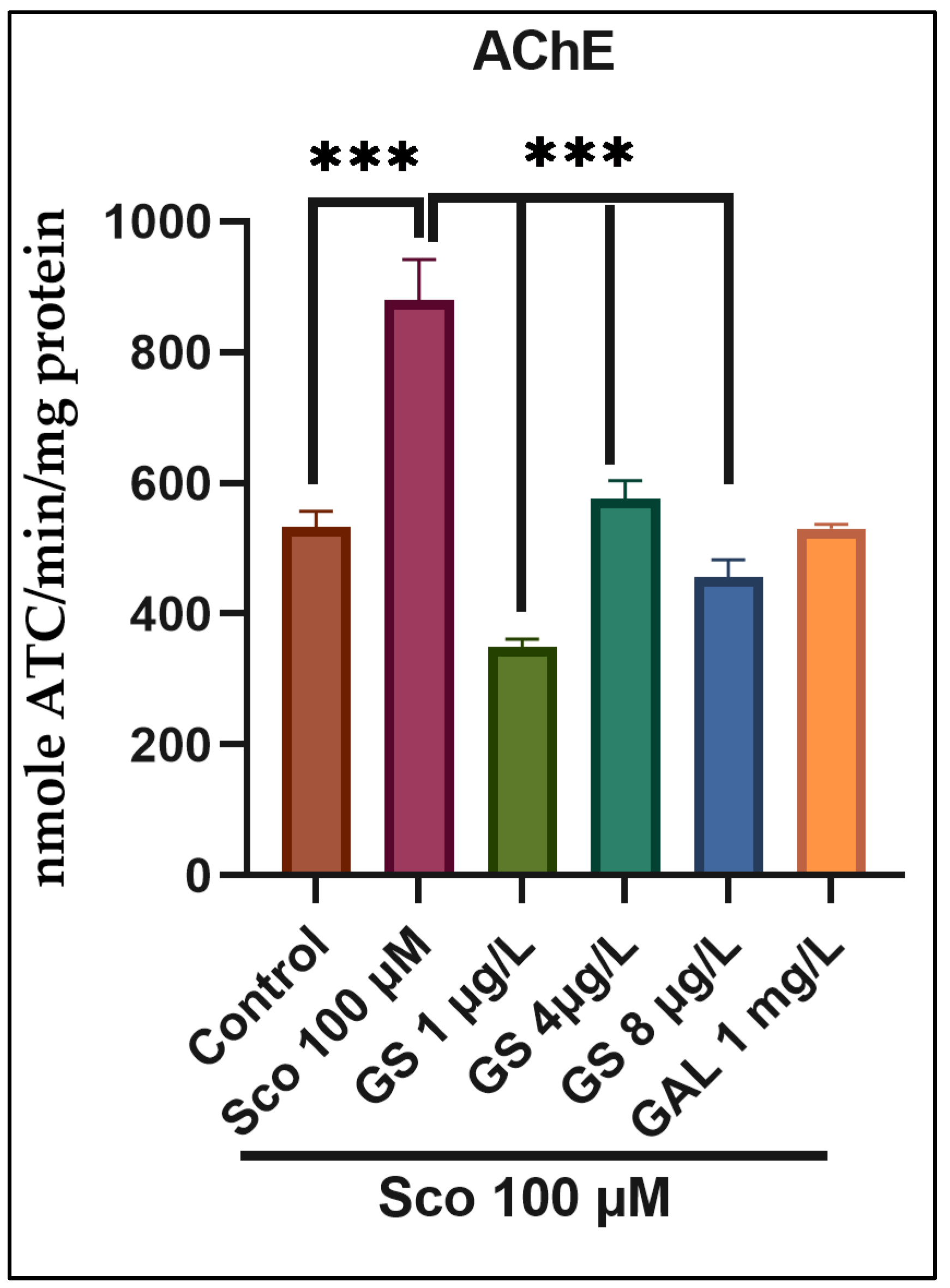
2.3. Effects on the Brain AChE Activity
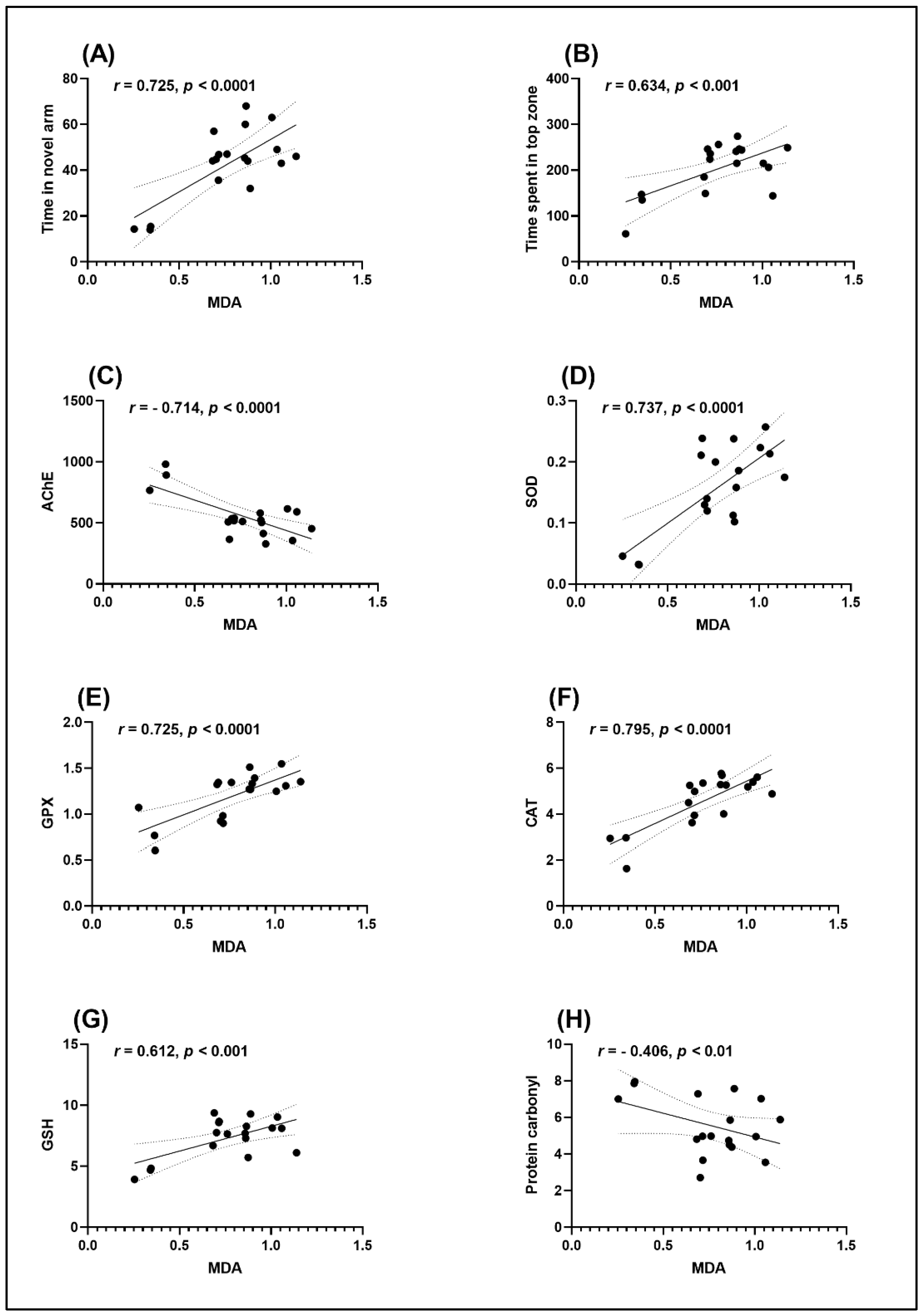
2.4. Effects on Brain Antioxidant Capacity
3. Materials and Methods
3.1. Plant Material and Extraction
3.2. High-Performance Liquid Chromatograph (HPLC-PDA)
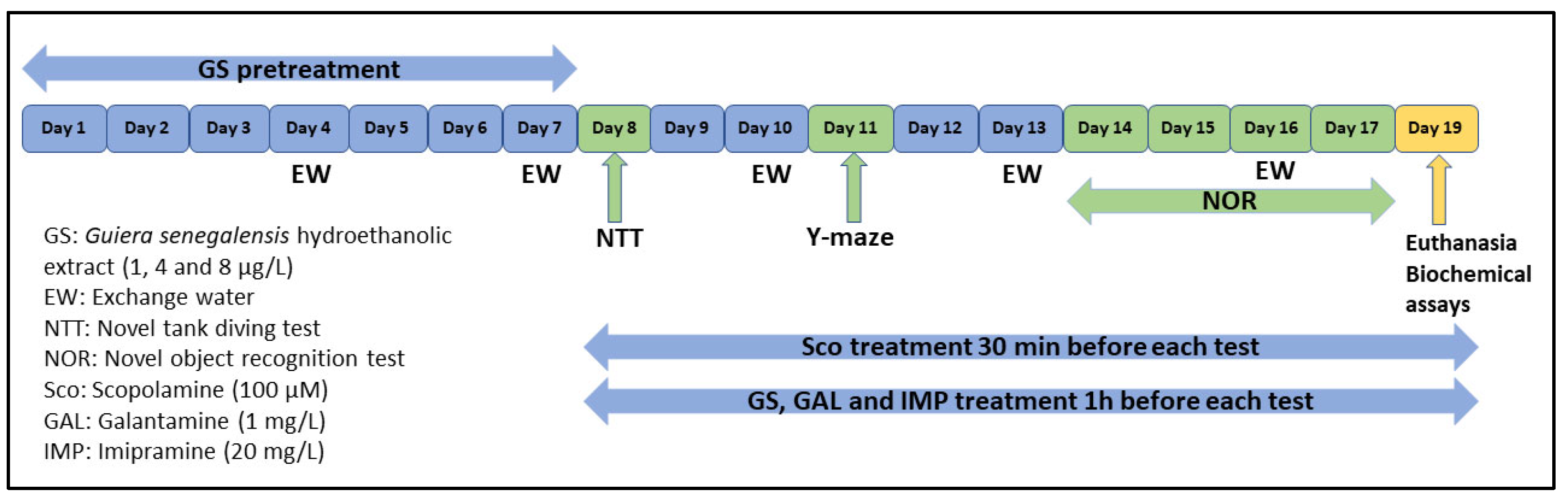
3.3. Animals and Treatment
3.4. Behavioral Analyses
3.4.1. Novel Tank Diving Test (NTT)
3.4.2. Y-Maze Test
3.4.3. Novel Object Recognition Test (NOR)
3.5. Biochemical Parameter Assay
3.5.1. Determination of the AChE Activity
3.5.2. Determination of the SOD Activity
3.5.3. Determination of the CAT Activity
3.5.4. Determination of the GPX Activity
3.5.5. Determination of the GSH Level
3.5.6. Determination of the Protein Carbonyl Level
3.5.7. Determination of the MDA Level
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hritcu, L.; Noumedem, J.A.; Cioanca, O.; Hancianu, M.; Kuete, V.; Mihasan, M. Methanolic extract of Piper nigrum fruits improves memory impairment by decreasing brain oxidative stress in amyloid beta(1-42) rat model of Alzheimer’s disease. Cell. Mol. Neurobiol. 2014, 34, 437–449. [Google Scholar] [CrossRef] [PubMed]
- Foyet, H.S.; Keugong Wado, E.; Ngatanko Abaissou, H.H.; Assongalem, E.A.; Eyong, O.K. Anticholinesterase and antioxidant potential of hydromethanolic extract of Ziziphus mucronata (Rhamnaceae) leaves on scopolamine-induced memory and cognitive dysfunctions in mice. Evid. Based Complement. Altern. Med. 2019, 2019, 4568401. [Google Scholar] [CrossRef] [PubMed]
- Ionita, R.; Postu, P.A.; Mihasan, M.; Gorgan, D.L.; Hancianu, M.; Cioanca, O.; Hritcu, L. Ameliorative effects of Matricaria chamomilla L. hydroalcoholic extract on scopolamine-induced memory impairment in rats: A behavioral and molecular study. Phytomedicine 2018, 47, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Cheignon, C.; Tomas, M.; Bonnefont-Rousselot, D.; Faller, P.; Hureau, C.; Collin, F. Oxidative stress and the amyloid beta peptide in Alzheimer’s disease. Redox Biol. 2018, 14, 450–464. [Google Scholar] [CrossRef]
- Rebe, R.; Wado, E.; Ngatanko, A.; Kamleu, B.; Kenko, L.; Damo, J.; Bahane, A.; Oben, E.; Foyet, H.S. Neuroprotective, behavioral and biochemical effects of aqueous extract of Hibiscus cannabinus L. on aluminum chloride induced Alzheimer’s disease in male Wistar rats. Acad. J. Med. Plants 2021, 9, 118–134. [Google Scholar]
- Todirascu-Ciornea, E.; El-Nashar, H.A.S.; Mostafa, N.M.; Eldahshan, O.A.; Boiangiu, R.S.; Dumitru, G.; Hritcu, L.; Singab, A.N.B. Schinus terebinthifolius Essential Oil Attenuates Scopolamine-Induced Memory Deficits via Cholinergic Modulation and Antioxidant Properties in a Zebrafish Model. Evid. Based Complement. Altern. Med. 2019, 2019, 5256781. [Google Scholar] [CrossRef]
- Kouémou, N.E.; Taiwe, G.S.; Moto, F.C.O.; Pale, S.; Ngoupaye, G.T.; Njapdounke, J.S.K.; Nkantchoua, G.C.N.; Pahaye, D.B.; Bum, E.N. Nootropic and Neuroprotective Effects of Dichrocephala integrifolia on Scopolamine Mouse Model of Alzheimer’s Disease. Front. Pharmacol. 2017, 8, 847. [Google Scholar] [CrossRef]
- Hervé, H.; Abaissou, N.; Harquin Simplice, F.; Keugong Wado, E.; Kamleu Nkwingwa, B.; Linda, J.; Kamda, D.; Blondelle, L.; Djomessie, K.; Poroch, V.; et al. Cholinergic Enhancing and Antioxidant Effect of Vigna subterranea (L.) Verdc. (Fabaceae) Landrace Aqueous Extract on Scopolamine-Induced Amnesia in Male Swiss Mice. Rev. Chim. 2019, 70, 4336. [Google Scholar]
- Kim, M.S.; Lee, D.Y.; Lee, J.; Kim, H.W.; Sung, S.H.; Han, J.S.; Jeon, W.K. Terminalia chebula extract prevents scopolamine-induced amnesia via cholinergic modulation and anti-oxidative effects in mice. BMC Complement. Altern. Med. 2018, 18, 136. [Google Scholar] [CrossRef]
- Capatina, L.; Boiangiu, R.S.; Dumitru, G.; Napoli, E.M.; Ruberto, G.; Hritcu, L.; Todirascu-Ciornea, E. Rosmarinus officinalis Essential Oil Improves Scopolamine-Induced Neurobehavioral Changes via Restoration of Cholinergic Function and Brain Antioxidant Status in Zebrafish (Danio rerio). Antioxidants 2020, 9, 62. [Google Scholar] [CrossRef]
- Balaban, H.; Nazıroğlu, M.; Demirci, K.; Övey, İ.S. The Protective Role of Selenium on Scopolamine-Induced Memory Impairment, Oxidative Stress, and Apoptosis in Aged Rats: The Involvement of TRPM2 and TRPV1 Channels. Mol. Neurobiol. 2017, 54, 2852–2868. [Google Scholar] [CrossRef] [PubMed]
- Ramandeep, K.; Shaba, P.; Sidharth, M.; Deepa, K.; Sanjeev, K. Neuroprotective Effect of Ellagic Acid Against Chronically Scopolamine Induced Alzheimer’s Type Memory and Cognitive Dysfunctions: Possible Behavioural and Biochemical Evidences. Ann. Clin. Transl. Neurol. 2015, 1, 45–64. [Google Scholar]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.J.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef] [PubMed]
- Dumitru, G.; El-Nashar, H.A.S.; Mostafa, N.M.; Eldahshan, O.A.; Boiangiu, R.S.; Todirascu-Ciornea, E.; Hritcu, L.; Singab, A.N.B. Agathisflavone isolated from Schinus polygamus (Cav.) Cabrera leaves prevents scopolamine-induced memory impairment and brain oxidative stress in zebrafish (Danio rerio). Phytomedicine 2019, 58, 152889. [Google Scholar] [CrossRef]
- Levin, E.D. Zebrafish assessment of cognitive improvement and anxiolysis: Filling the gap between in vitro and rodent models for drug development. Rev. Neurosci. 2011, 22, 75. [Google Scholar] [CrossRef]
- Brinza, I.; Abd-Alkhalek, A.M.; El-Raey, M.A.; Boiangiu, R.S.; Eldahshan, O.A.; Hritcu, L. Ameliorative effects of rhoifolin in scopolamine-induced amnesic zebrafish (Danio rerio) model. Antioxidants 2020, 9, 580. [Google Scholar] [CrossRef]
- Kinda, P.; Zerbo, P.; Guenné, S.; Compaoré, M.; Ciobica, A.; Kiendrebeogo, M. Medicinal Plants Used for Neuropsychiatric Disorders Treatment in the Hauts Bassins Region of Burkina Faso. Medicines 2017, 4, 32. [Google Scholar] [CrossRef]
- Siddig Hamad, M.; Elsafi, H.; Saeed, A.; Fadul, E. A Review on the Taxonomy, Ethnobotany, Phytochemistry and Pharmacology of Guriea senegalensis J. F. Gmel. (Combretaceae). Med. Aromat. Plants 2017, 6, 153–165. [Google Scholar] [CrossRef]
- Somboro, A.A.; Patel, K.; Diallo, D.; Sidibé, L.; Chalchat, J.C.; Figueredo, G.; Ducki, S.; Troin, Y.; Chalard, P. An ethnobotanical and phytochemical study of the African medicinal plant Guiera senegalensis J. F. Gmel. J. Med. Plants Res. 2011, 5, 1639–1651. [Google Scholar] [CrossRef]
- Sombie, P.A.E.D.; Hilou, A.; Mounier, C.; Coulibaly, A.Y.; Kiendrebeo, M.; Millogo, J.F.; Nacoulma, O.G. Antioxidant and Anti-inflammatory Activities from Galls of Guiera senegalensis J.F. Gmel (Combretaceae). Res. J. Med. Plant 2011, 5, 448–461. [Google Scholar] [CrossRef]
- Somboro, A.A.; Diallo, D.; Sidibe, L.; Traore, N.; Fofana, B.; Bouare, S.; Chalard, P.; Chalchat, J.C.; Figueredo, G.; Troin, Y. Activités anticholinestérasiques des alcaloïdes totaux extraits des feuilles, fruits, écorces de racines et écorces de tronc de Guiera senegalensis, une plante médicinale Malienne. Int. J. Biol. Chem. Sci. 2014, 7, 1723–1728. [Google Scholar] [CrossRef][Green Version]
- Abubakr, M.; Sirag, N.; Osman, I.; Osman, M.; Abakar, S.; Aboul-Enein, A. Anticancer and antioxidant activities of Guiera senegalensis. Sudan J. Med. Sci. 2014, 8, 135–140. [Google Scholar] [CrossRef]
- Hamadnalla, H.M.Y. Phytochemical Investigation, Antimicrobial, Antioxidant and Anti-Diabetic Potential of Guiera Senegalensis Leaves Extracts. Cytokines Relat. Miner. Dust Induc. Dis. 2020, 4, 1–4. [Google Scholar] [CrossRef]
- Bucar, F.; Resch, M.; Bauer, R.; Burits, M.; Knauder, E.; Schubert-Zsilavecz, M. 5-Methylflavasperone and rhamnetin from Guiera senegalensis and their antioxidative and 5-lipoxygenase inhibitory activity. Pharmazie 1998, 53, 875–878. [Google Scholar] [CrossRef]
- Alam, P.; Parvez, M.K.; Arbab, A.H.; Al-Dosari, M.S. Quantitative analysis of rutin, quercetin, naringenin, and gallic acid by validated RP- and NP-HPTLC methods for quality control of anti-HBV active extract of Guiera senegalensis. Pharm. Biol. 2017, 55, 1317–1323. [Google Scholar] [CrossRef]
- Dirar, A.I.; Devkota, H.P. Ethnopharmacological uses, phytochemistry and pharmacological activities of Guiera senegalensis J.F. Gmel. (Combretaceae). J. Ethnopharmacol. 2021, 267, 113433. [Google Scholar] [CrossRef]
- Sombie, P.A.E.D.; Hilou, A.; Coulibaly, A.Y.; Tibiri, A.; Kiendrebeogo, M.; Nacoulma, O.G. Brain protective and erythrocytes hemolysis inhibition potentials from galls of Guiera senegalensis J.F. Gmel (Combretaceae). J. Pharmacol. Toxicol. 2011, 6, 361–370. [Google Scholar] [CrossRef][Green Version]
- Rinne, J.O.; Kaasinen, V.; Järvenpää, T.; Någren, K.; Roivainen, A.; Yu, M.; Oikonen, V.; Kurki, T. Brain acetylcholinesterase activity in mild cognitive impairment and early Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 2003, 74, 113–115. [Google Scholar] [CrossRef]
- Terry, A.V.; Buccafusco, J.J. The cholinergic hypothesis of age and Alzheimer’s disease-related cognitive deficits: Recent challenges and their implications for novel drug development. J. Pharmacol. Exp. Ther. 2003, 306, 821–827. [Google Scholar] [CrossRef]
- Trinh, N.H.; Hoblyn, J.; Mohanty, S.; Yaffe, K. Efficacy of cholinesterase inhibitors in the treatment of neuropsychiatric symptoms and functional impairment in Alzheimer disease: A meta-analysis. J. Am. Med. Assoc. 2003, 289, 210–216. [Google Scholar] [CrossRef]
- Melo, J.B.; Agostinho, P.; Oliveira, C.R. Involvement of oxidative stress in the enhancement of acetylcholinesterase activity induced by amyloid beta-peptide. Neurosci. Res. 2003, 45, 117–127. [Google Scholar] [CrossRef]
- El-Sherbiny, D.A.; Khalifa, A.E.; Attia, A.S.; Eldenshary, E.E.D.S. Hypericum perforatum extract demonstrates antioxidant properties against elevated rat brain oxidative status induced by amnestic dose of scopolamine. Pharmacol. Biochem. Behav. 2003, 76, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Dumont, M.; Beal, M.F. Neuroprotective strategies involving ROS in Alzheimer’s disease. Free Radic. Biol. Med. 2011, 51, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Beppe, G.J.; Kenko Djoumessie, L.B.; Keugong Wado, E.; Ngatanko Abaïssou, H.H.; Nkwingwa, B.K.; Damo Kamda, J.L.; Nhouma, R.R.; Foyet, H.S. Aqueous Root Bark Extract of Daniellia oliveri (Hutch. & Dalz.) (Fabaceae) Protects Neurons against Diazepam-Induced Amnesia in Mice. Evid. Based. Complement. Alternat. Med. 2020, 2020, 7815348. [Google Scholar] [CrossRef]
- Ren, Z.; Li, Y.; Zhang, R.; Li, Y.; Yang, Z.; Yang, H. Ferulic acid exerts neuroprotective effects against cerebral ischemia/reperfusion-induced injury via antioxidant and anti-apoptotic mechanisms in vitro and in vivo. Int. J. Mol. Med. 2017, 40, 1444–1456. [Google Scholar] [CrossRef]
- Keugong Wado, E.; Kubicki, M.; Ngatanko, A.H.H.; Léa Blondelle, K.D.; Jorelle Linda, D.; Roland, R.N.; Balbine, K.; Lamshoeft, M.; Assongalem, A.E.; Foyet, H.S. Anxiolytic and antidepressant effects of Ziziphus mucronata hydromethanolic extract in male rats exposed to unpredictable chronic mild stress: Possible mechanisms of actions. J. Ethnopharmacol. 2020, 260, 112987. [Google Scholar] [CrossRef]
- Tu, Q.; Wang, R.; Ding, B.; Zhong, W.; Cao, H. Protective and antioxidant effect of Danshen polysaccharides on cerebral ischemia/reperfusion injury in rats. Int. J. Biol. Macromol. 2013, 60, 268–271. [Google Scholar] [CrossRef]
- Uddin, M.S.; Mamun, A.; Al Hossain, M.S.; Ashaduzzaman, M.; Noor, M.A.A.; Hossain, M.S.; Uddin, M.J.; Sarker, J.; Asaduzzaman, M.; Uddin, M.S.; et al. Neuroprotective Effect of Phyllanthus acidus L. on Learning and Memory Impairment in Scopolamine-Induced Animal Model of Dementia and Oxidative Stress: Natural Wonder for Regulating the Development and Progression of Alzheimer’s Disease. Adv. Alzheimer’s Dis. 2016, 5, 53–72. [Google Scholar] [CrossRef]
- Parvez, M.K.; Alam, P.; Arbab, A.H.; Al-Dosari, M.S.; Alhowiriny, T.A.; Alqasoumi, S.I. Analysis of antioxidative and antiviral biomarkers β-amyrin, β-sitosterol, lupeol, ursolic acid in Guiera senegalensis leaves extract by validated HPTLC methods. Saudi Pharm. J. SPJ Off. Publ. Saudi Pharm. Soc. 2018, 26, 685–693. [Google Scholar] [CrossRef]
- Imam, N.; Abdelwahab, S.I.; Taha, M.M.; Mohan, S.; Alhazmi, H.; Ahmed, R.H. Antioxidant and hepatoprotective activities of Blepharis linariifolia pers and Guiera senegalensis J. F. Gmel. against CCl4-induced hepatotoxicity. World J. Pharm. Res. 2018, 7, 70–80. [Google Scholar]
- Cachat, J.; Stewart, A.; Grossman, L.; Gaikwad, S.; Kadri, F.; Chung, K.M.; Wu, N.; Wong, K.; Roy, S.; Suciu, C.; et al. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat. Protoc. 2010, 5, 1786–1799. [Google Scholar] [CrossRef] [PubMed]
- Cognato, G.P.; Bortolotto, J.W.; Blazina, A.R.; Christoff, R.R.; Lara, D.R.; Vianna, M.R.; Bonan, C.D. Y-maze memory task in zebrafish (Danio rerio): The role of glutamatergic and cholinergic systems on the acquisition and consolidation periods. Neurobiol. Learn. Mem. 2012, 98, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zhu, Y.; Wang, Y.; Qi, S.; Wang, Y.; Ma, C.; Li, S.; Jiang, B.; Cheng, X.; Wang, Z.; et al. Anti-amnesic effect of extract and alkaloid fraction from aerial parts of Peganum harmala on scopolamine-induced memory deficits in mice. J. Ethnopharmacol. 2017, 204, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Gaspary, K.V.; Reolon, G.K.; Gusso, D.; Bonan, C.D. Novel object recognition and object location tasks in zebrafish: Influence of habituation and NMDA receptor antagonism. Neurobiol. Learn. Mem. 2018, 155, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Boiangiu, R.S.; Mihasan, M.; Gorgan, D.L.; Stache, B.A.; Hritcu, L. Anxiolytic, Promnesic, Anti-Acetylcholinesterase and Antioxidant Effects of Cotinine and 6-Hydroxy-L-Nicotine in Scopolamine-Induced Zebrafish (Danio rerio) Model of Alzheimer’s Disease. Antioxidants 2021, 10, 212. [Google Scholar] [CrossRef]
- Ellman, G.; Courtney, K.; Andres, V.J.; Feather-Stone, R. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Winterbourn, C.; Hawkins, R.; Brian, M.; Carrell, R. The estimation of red cell superoxide dismutase activity. J. Lab. Clin. Med. 1975, 85, 337. [Google Scholar] [PubMed]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef]
- Sharma, M.; Gupta, Y.K. Chronic treatment with trans resveratrol prevents intracerebroventricular streptozotocin induced cognitive impairment and oxidative stress in rats. Life Sci. 2002, 7, 2489–2498. [Google Scholar] [CrossRef]
- Fukuzawa, K.; Tokumura, A. Glutathione peroxidase activity in tissues of vitamin E-deficient mice. J. Nutr. Sci. Vitaminol. 1976, 22, 405–407. [Google Scholar] [CrossRef]
- Oliver, C.N.; Ahn, B.W.; Moerman, E.J.; Goldstein, S.; Stadtman, E.R. Age-related changes in oxidized proteins. J. Biol. Chem. 1987, 262, 5488–5491. [Google Scholar] [CrossRef]
- Luo, S.; Wehr, N.B. Protein carbonylation: Avoiding pitfalls in the 2,4-dinitrophenylhydrazine assay. Redox. Rep. 2009, 14, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemical Group | Guiera senegalensis Extract | |
|---|---|---|
| Compounds | Quantity (µg/g Dry Extract) | |
| Flavonoids | Quercetin-3-O-arabinoside | 316.30 |
| Catechin | 108.57 | |
| Luteolin-7-O-glucoside | 56.46 | |
| Rutoside | 205.71 | |
| Quercetin | 306.86 | |
| Apigenin | 141.65 | |
| Epigallocatechin | 120.05 | |
| Polyphenolic acids | Caffeic acid | 126.45 |
| Chlorogenic acid | 55.20 | |
| Cinnamic acid | 106.14 | |
| Ellagic acid | 80.13 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damo, J.L.K.; Boiangiu, R.S.; Brinza, I.; Kenko Djoumessi, L.B.; Rebe, R.N.; Kamleu, B.N.; Guedang, S.D.N.; Camdi, G.W.; Bouvourné, P.; Keugong, E.W.; et al. Neuroprotective Potential of Guiera senegalensis (Combretaceae) Leaf Hydroethanolic Extract against Cholinergic System Dysfunctions and Oxidative Stress in Scopolamine-Induced Cognitive Impairment in Zebrafish (Danio rerio). Plants 2022, 11, 1149. https://doi.org/10.3390/plants11091149
Damo JLK, Boiangiu RS, Brinza I, Kenko Djoumessi LB, Rebe RN, Kamleu BN, Guedang SDN, Camdi GW, Bouvourné P, Keugong EW, et al. Neuroprotective Potential of Guiera senegalensis (Combretaceae) Leaf Hydroethanolic Extract against Cholinergic System Dysfunctions and Oxidative Stress in Scopolamine-Induced Cognitive Impairment in Zebrafish (Danio rerio). Plants. 2022; 11(9):1149. https://doi.org/10.3390/plants11091149
Chicago/Turabian StyleDamo, Jorelle Linda Kamda, Razvan Stefan Boiangiu, Ion Brinza, Léa Blondelle Kenko Djoumessi, Roland Nhouma Rebe, Balbine Nkwingwa Kamleu, Simon Désiré Nyayi Guedang, Guillaume Woumitna Camdi, Parfait Bouvourné, Eglantine Wado Keugong, and et al. 2022. "Neuroprotective Potential of Guiera senegalensis (Combretaceae) Leaf Hydroethanolic Extract against Cholinergic System Dysfunctions and Oxidative Stress in Scopolamine-Induced Cognitive Impairment in Zebrafish (Danio rerio)" Plants 11, no. 9: 1149. https://doi.org/10.3390/plants11091149
APA StyleDamo, J. L. K., Boiangiu, R. S., Brinza, I., Kenko Djoumessi, L. B., Rebe, R. N., Kamleu, B. N., Guedang, S. D. N., Camdi, G. W., Bouvourné, P., Keugong, E. W., Ngatanko, H. H. A., Cioanca, O., Hancianu, M., Foyet, H. S., & Hritcu, L. (2022). Neuroprotective Potential of Guiera senegalensis (Combretaceae) Leaf Hydroethanolic Extract against Cholinergic System Dysfunctions and Oxidative Stress in Scopolamine-Induced Cognitive Impairment in Zebrafish (Danio rerio). Plants, 11(9), 1149. https://doi.org/10.3390/plants11091149