
Invasive Stages within Alien Species and Hutchinson’s Duality: An Example Using Invasive Plants of the Family Fabaceae in Central Chile



 ,
,
Abstract
:1. Introduction
2. Results
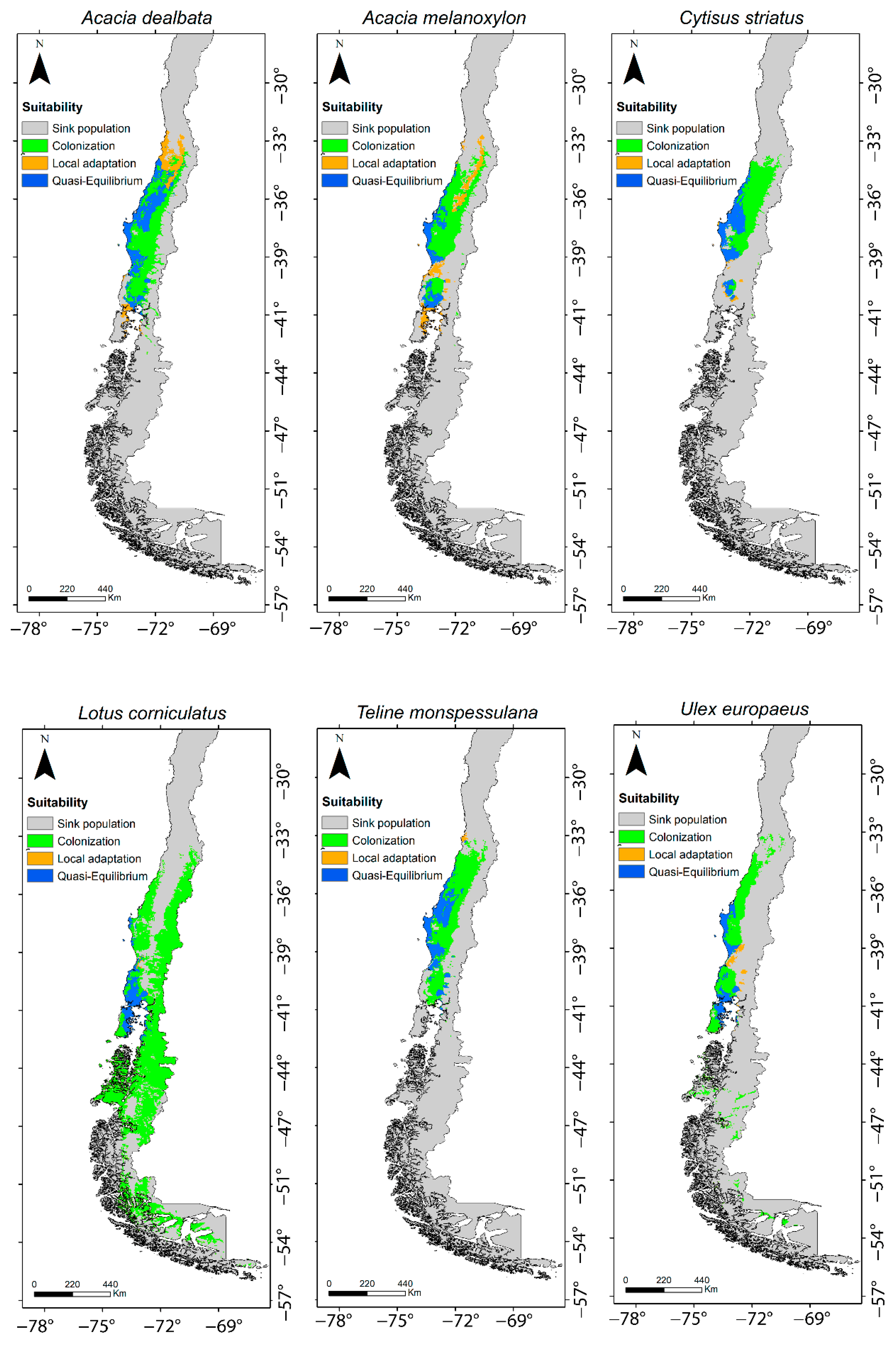
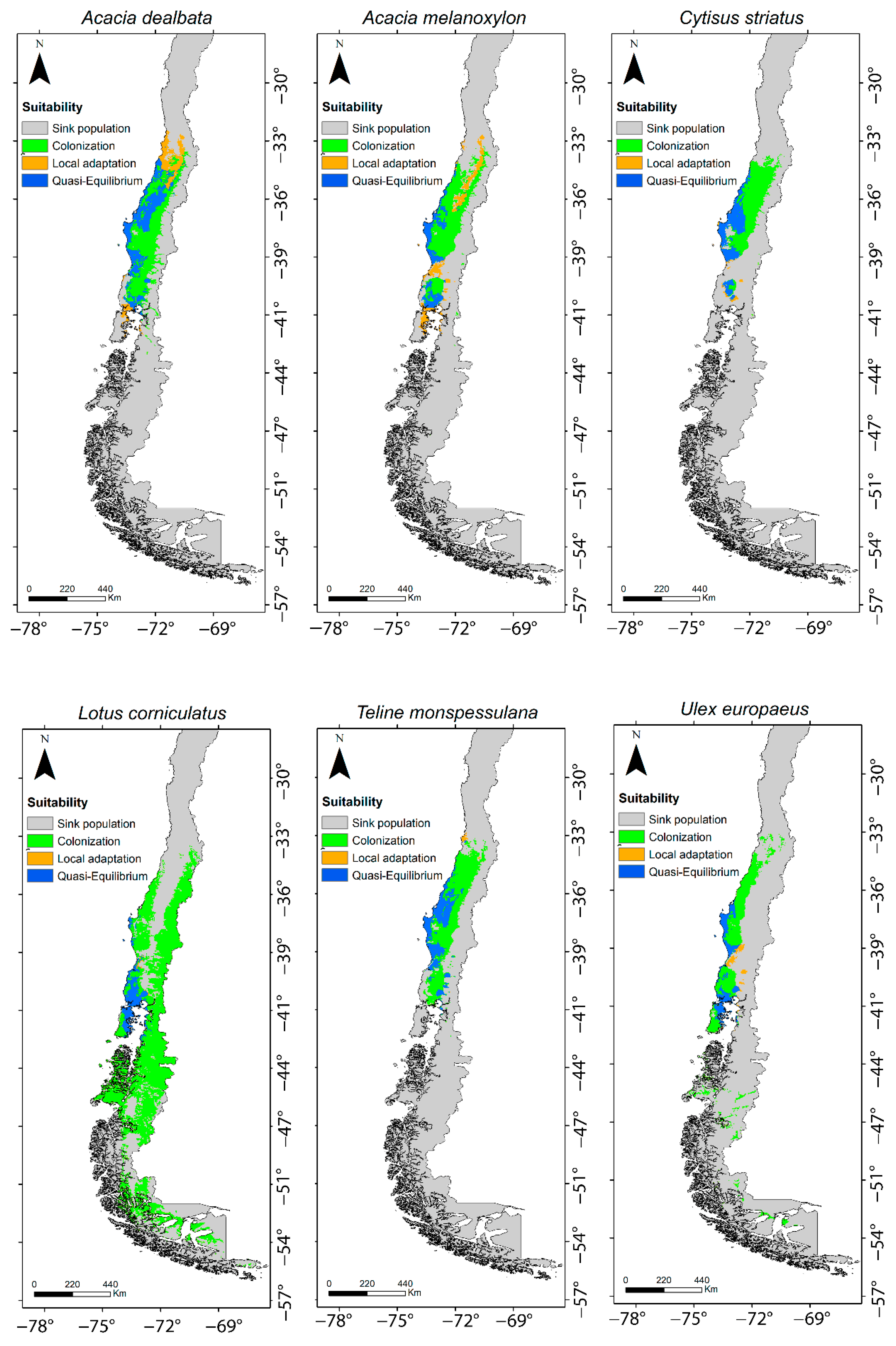
2.1. SDMs from Global and Regional Climatic Niches
2.2. Invasive Stages
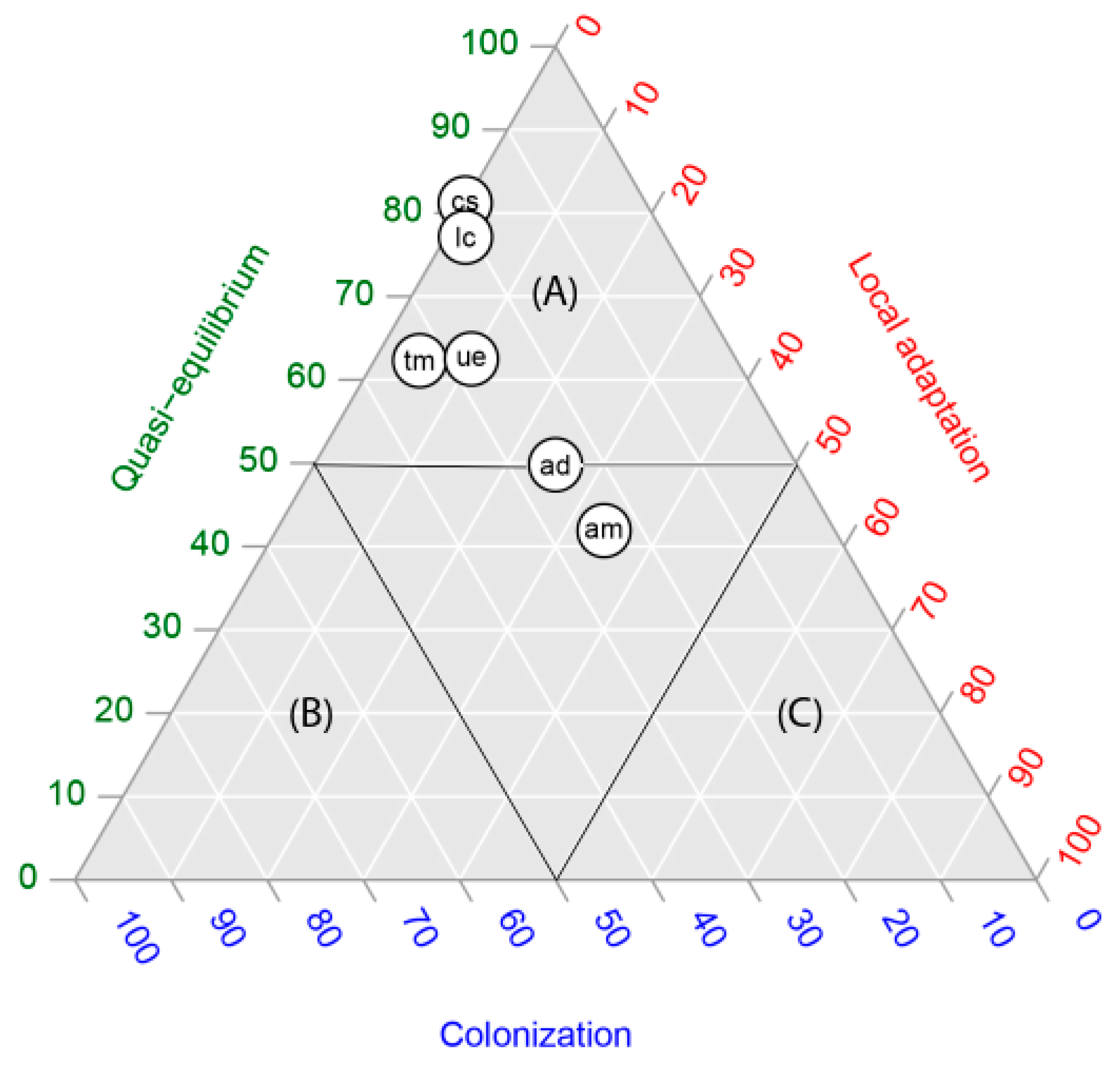
2.3. Ternary Plots
3. Discussion
3.1. Invasive Stages and Hypothesis
3.2. Hutchinson’s Duality
4. Materials and Methods
4.1. Study Area
4.2. Species Distribution Models (SDMs)
4.3. Invasive Stages
4.4. Ternary Plots
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Richardson, D.M.; Pysek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and Invasion of Alien Plants: Concepts and Definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Crooks, J.A. Lag Times and Exotic Species: The Ecology and Management of Biological Invasions in Slow-Motion. Écoscience 2005, 12, 316–329. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Peterson, A.; Soberón, J.; Overton, J.; Aragón, P.; Lobo, J. Use of Niche Models in Invasive Species Risk Assessments. Biol. Invasions 2011, 13, 2785–2797. [Google Scholar] [CrossRef]
- Soberon, J.; Nakamura, M. Colloquium Papers: Niches and Distributional Areas: Concepts, Methods, and Assumptions. Proc. Natl. Acad. Sci. USA 2009, 106, 19644–19650. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Thuiller, W. Predicting Species Distribution: Offering More than Simple Habitat Models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Langdon, B.; Pauchard, A.; Bustamante, R.O. Acacia dealbata Invasion in Chile: Surprises from Climatic Niche and Species Distribution Models. Ecol. Evol. 2019, 9, 7562–7573. [Google Scholar] [CrossRef] [Green Version]
- Araújo, M.B.; Pearson, R.G. Equilibrium of Species’ Distributions with Climate. Ecography 2005, 28, 693–695. [Google Scholar] [CrossRef]
- Goncalves, E.; Herrera, I.; Duarte, M.; Bustamante, R.O.; Lampo, M.; Velásquez, G.; Sharma, G.P.; García-Rangel, S. Global Invasion of Lantana Camara: Has the Climatic Niche Been Conserved across Continents? PLoS ONE 2014, 9, e111468. [Google Scholar] [CrossRef] [Green Version]
- Broennimann, O.; Treier, U.A.; Müller-Schärer, H.; Thuiller, W.; Peterson, A.T.; Guisan, A. Evidence of Climatic Niche Shift during Biological Invasion. Ecol. Lett. 2007, 10, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Pearman, P.B.; Guisan, A.; Broennimann, O.; Randin, C.F. Niche Dynamics in Space and Time. Trends Ecol. Evol. 2008, 23, 149–158. [Google Scholar] [CrossRef]
- Parker, I.M.; Gilbert, G.S. When there is no escape: The effects of natural enemies on native, invasive, and noninvasive plants. Ecology 2007, 88, 1210–1224. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.T. Predicting the Geography of Species’ Invasions via Ecological Niche Modeling. Q. Rev. Biol. 2003, 78, 419–433. [Google Scholar] [CrossRef] [Green Version]
- Gallien, L.; Douzet, R.; Pratte, S.; Zimmermann, N.E.; Thuiller, W. Invasive Species Distribution Models—How Violating the Equilibrium Assumption Can Create New Insights: Beyond the Equilibrium Assumption of SDMs. Glob. Ecol. Biogeogr. 2012, 21, 1126–1136. [Google Scholar] [CrossRef]
- Peterson, A.; Soberón, J.; Pearson, R.G.; Anderson, R.; Martinez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions; Princeton University Press: Oxford, UK, 2011. [Google Scholar]
- Guisan, A.; Petitpierre, B.; Broennimann, O.; Daehler, C.; Kueffer, C. Unifying Niche Shift Studies: Insights from Biological Invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Václavík, T.; Meentemeyer, R.K. Equilibrium or Not? Modelling Potential Distribution of Invasive Species in Different Stages of Invasion: Equilibrium and Invasive Species Distribution Models. Divers. Distrib. 2012, 18, 73–83. [Google Scholar] [CrossRef]
- Taucare-Ríos, A.; Bizama, G.; Bustamante, R.O. Using Global and Regional Species Distribution Models (SDM) to Infer the Invasive Stage of Latrodectus Geometricus (Araneae: Theridiidae) in the Americas. Environ. Entomol. 2016, 45, 1379–1385. [Google Scholar] [CrossRef]
- Hargreaves, A.L.; Samis, K.E.; Eckert, C.G. Are Species’ Range Limits Simply Niche Limits Writ Large? A Review of Transplant Experiments beyond the Range. Am. Nat. 2013, 183, 157–173. [Google Scholar] [CrossRef] [Green Version]
- Colwell, R.K.; Rangel, T.F. Colloquium Papers: Hutchinson’s Duality: The Once and Future Niche. Proc. Natl. Acad. Sci. USA 2009, 106, 19651–19658. [Google Scholar] [CrossRef] [Green Version]
- Foggi, B.; Benesperi, R.; Viciani, D.; Giunti, M.; Lastrucci, L. Long-Term Monitoring of an Invasion Process: The Case of an Isolated Small Wetland on a Mediterranean Island, Second Stage: Toward a Complete Restoration. Biologia 2014, 69, 977–985. [Google Scholar] [CrossRef]
- Grime, J.P. The C-S-R Model of Primary Plant Strategies—Origins, Implications and Tests. In Plant Evolutionary Biology; Gottlieb, L.D., Jain, S.K., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 371–393. ISBN 978-94-010-7036-2. [Google Scholar]
- Guo, W.-Y.; van Kleunen, M.; Winter, M.; Weigelt, P.; Stein, A.; Pierce, S.; Pergl, J.; Moser, D.; Maurel, N.; Lenzner, B.; et al. The Role of Adaptive Strategies in Plant Naturalization. Ecol. Lett. 2018, 21, 1380–1389. [Google Scholar] [CrossRef]
- Kannegiesser, S. Antecedentes Generales Sobre Acacia melanoxylon (Aromo Australiano). CIFOR 1989, 3, 90–97. [Google Scholar] [CrossRef]
- La Pierre, K.J.; Simms, E.L.; Tariq, M.; Zafar, M.; Porter, S.S. Invasive Legumes Can Associate with Many Mutualists of Native Legumes, but Usually Do Not. Ecol. Evol. 2017, 7, 8599–8611. [Google Scholar] [CrossRef] [Green Version]
- Parker, I.M.; Haubensak, K.A. Comparative Pollinator Limitation of Two Non-Native Shrubs: Do Mutualisms Influence Invasions? Oecologia 2002, 130, 250–258. [Google Scholar] [CrossRef]
- Atlan, A.; Hornoy, B.; Delerue, F.; Gonzalez, M.; Pierre, J.-S.; Tarayre, M. Phenotypic Plasticity in Reproductive Traits of the Perennial Shrub Ulex europaeus in Response to Shading: A Multi-Year Monitoring of Cultivated Clones. PLoS ONE 2015, 10, e0137500. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, A.; Thomann, T.; Agret, S. Foreign Exploration & Host Specificity Testing of Biological Control Agents of French Broom in California. 2002. Available online: https://www.cal-ipc.org/docs/ip/research/biocontrols/broom/pdf/ForeignExploration.pdf (accessed on 16 February 2022).
- Sheppard, A.; Thomann, T. Quantitative Field Surveys for the Selection of Biological Control Agents for Genista Monspessulana, Based on Host Range and Efficacy Assessment. In XI International Symposium on Biological Control of Weeds; CSIRO Entomology: Canberra, Australia, 2004; p. 162. [Google Scholar]
- Chan, K.; Turner, C. Discovery of the Gall Mite Aceria Genistae (Nalepa)(Acarina: Eriophyidae) on Gorse and French Broom in the United States. Pan-Pac. Entomol. 1998, 74, 55–57. [Google Scholar]
- Münzbergová, Z.; Herben, T. Seed, Dispersal, Microsite, Habitat and Recruitment Limitation: Identification of Terms and Concepts in Studies of Limitations. Oecologia 2005, 145, 1–8. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding Remarks. Cold Spring Harb. Symp. Quant. Biol. 1957, 22, 4l5–427. [Google Scholar] [CrossRef]
- Fuentes, N.; Pauchard, A.; Sánchez, P.; Esquivel, J.; Marticorena, A. A New Comprehensive Database of Alien Plant Species in Chile Based on Herbarium Records. Biol. Invasions 2013, 15, 847–858. [Google Scholar] [CrossRef]
- Dickinson, J.L.; Zuckerberg, B.; Bonter, D.N. Citizen Science as an Ecological Research Tool: Challenges and Benefits. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 149–172. [Google Scholar] [CrossRef] [Green Version]
- García, R.A.; Engler, M.L.; Peña, E.; Pollnac, F.W.; Pauchard, A. Fuel Characteristics of the Invasive Shrub Teline monspessulana (L.) K. Koch. Int. J. Wildland Fire 2015, 24, 372. [Google Scholar] [CrossRef]
- Pauchard, A.; Garcia, R.A.; Pena, E.; González, C.; Cavieres, L.A.; Bustamante, R.O. Positive Feedbacks between Plant Invasions and Fire Regimes: Teline monspessulana (L.) K. Koch (Fabaceae) in Central Chile. Biol. Invasions 2008, 10, 547–553. [Google Scholar] [CrossRef]
- Quiroz, C.; Pauchard, A.; Marticorena, A.; Cavieres, L.A. Manual de Plantas Invasoras del Centro-Sur de Chile; Laboratorio de Invasiones Biológicas: Concepción, Chile, 2009. [Google Scholar]
- Pauchard, A.; Alaback, P.B. Influence of Elevation, Land Use, and Landscape Context on Patterns of Alien Plant Invasions along Roadsides in Protected Areas of South-Central Chile. Conserv. Biol. 2004, 18, 238–248. [Google Scholar] [CrossRef]
- Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A Toolbox for Comparative Studies of Environmental Niche Models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum Entropy Modeling of Species Geographic Distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Species | Quasi-Equilibrium | Local Adaptation | Colonization | Total Occurrences (Sink Stage Excluded) |
|---|---|---|---|---|
| Acacia dealbata Link | 0.50 | 0.25 | 0.25 | 243 |
| Cytisus striatus (Hill) Rothm | 0.81 | 0.00 | 0.19 | 48 |
| Teline monpessulana (L.) K. Koch | 0.62 | 0.05 | 0.33 | 106 |
| Acacia melanoxylon R. Br. | 0.42 | 0.34 | 0.24 | 129 |
| Ulex europaeus Brot. | 0.63 | 0.10 | 0.28 | 80 |
| Lotus corniculatus L. | 0.77 | 0.02 | 0.21 | 48 |
| Average (Range) | 0.62 (0.42−0.81) | 0.13 (0.00–0.34) | 0.25 (0.19–0.33) | 109 (48–243) |
| Species | Quasi-Equilibrium | Local Adaptation | Colonization | Total Area (km2) |
|---|---|---|---|---|
| Acacia dealbata Link | 0.33 | 0.16 | 0.52 | 119.25 |
| Cytisus striatus (Hill) Rothm | 0.33 | 0.03 | 0.64 | 76.19 |
| Teline monpessulana (L.) K. Koch | 0.31 | 0.13 | 0.68 | 92.50 |
| Acacia melanoxylon R. Br. | 0.20 | 0.20 | 0.60 | 95.28 |
| Ulex europaeus Brot. | 0.23 | 0.05 | 0.72 | 78.80 |
| Lotus corniculatus L. | 0.08 | 0.004 | 0.91 | 233.30 |
| Average (Range) | 0.25 (0.08–0.33) | 0.08 (0.004–0.20) | 0.68 (0.52–0.91) | 115.89 (76.19–233.30) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bustamante, R.O.; Quiñones, D.; Duarte, M.; Goncalves, E.; Cavieres, L.A. Invasive Stages within Alien Species and Hutchinson’s Duality: An Example Using Invasive Plants of the Family Fabaceae in Central Chile. Plants 2022, 11, 1063. https://doi.org/10.3390/plants11081063
Bustamante RO, Quiñones D, Duarte M, Goncalves E, Cavieres LA. Invasive Stages within Alien Species and Hutchinson’s Duality: An Example Using Invasive Plants of the Family Fabaceae in Central Chile. Plants. 2022; 11(8):1063. https://doi.org/10.3390/plants11081063
Chicago/Turabian StyleBustamante, Ramiro O., Daniela Quiñones, Milen Duarte, Estefany Goncalves, and Lohengrin A. Cavieres. 2022. "Invasive Stages within Alien Species and Hutchinson’s Duality: An Example Using Invasive Plants of the Family Fabaceae in Central Chile" Plants 11, no. 8: 1063. https://doi.org/10.3390/plants11081063
APA StyleBustamante, R. O., Quiñones, D., Duarte, M., Goncalves, E., & Cavieres, L. A. (2022). Invasive Stages within Alien Species and Hutchinson’s Duality: An Example Using Invasive Plants of the Family Fabaceae in Central Chile. Plants, 11(8), 1063. https://doi.org/10.3390/plants11081063