
Effects of Auxin (Indole-3-butyric Acid) on Adventitious Root Formation in Peach-Based Prunus Rootstocks


 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
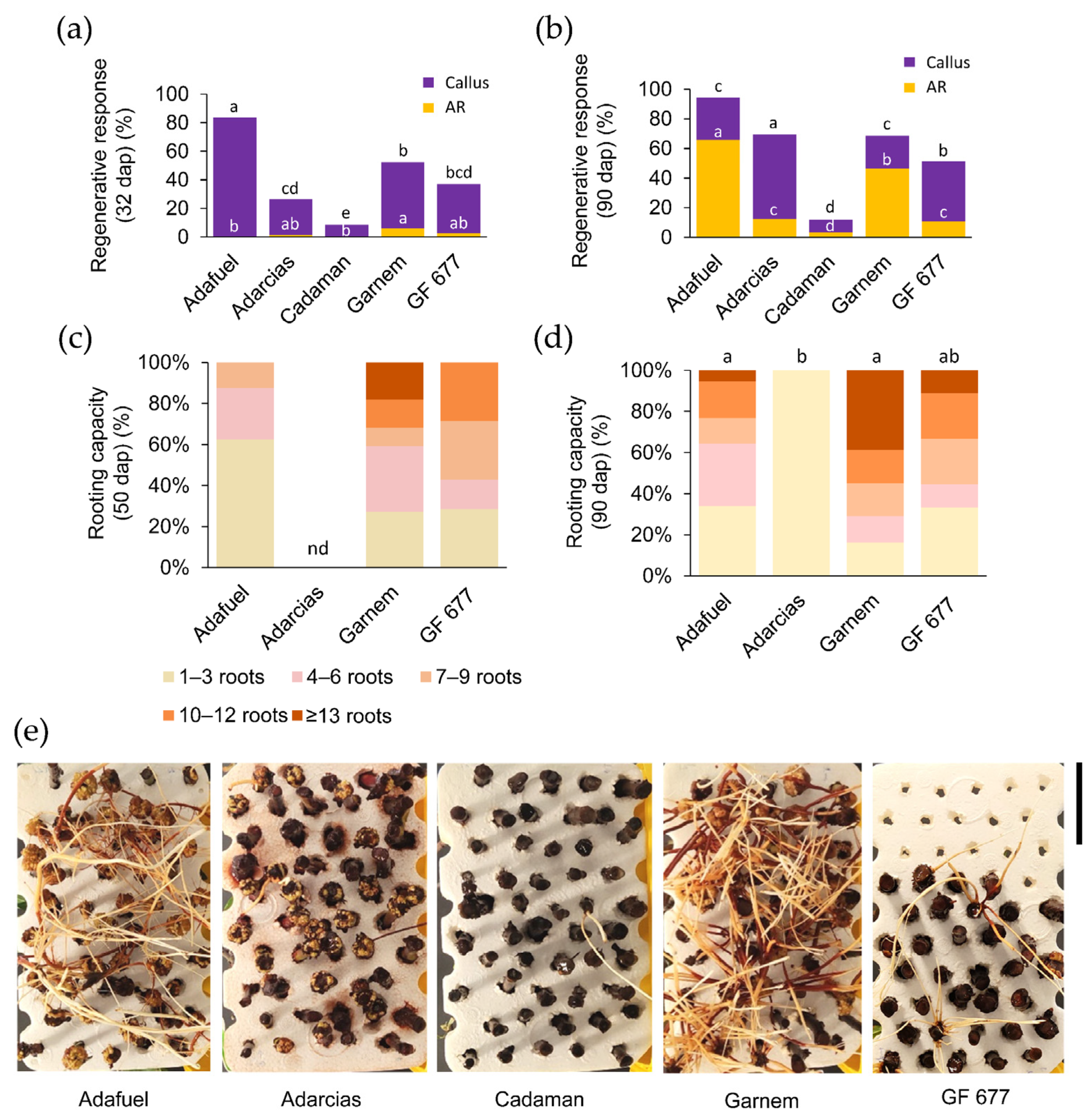
2.1. Rooting Response in Five Prunus Rootstocks during Hydroponic Culture
2.1.1. Callus Formation and Rooting Efficiency
2.1.2. Correlations between Regenerative Responses
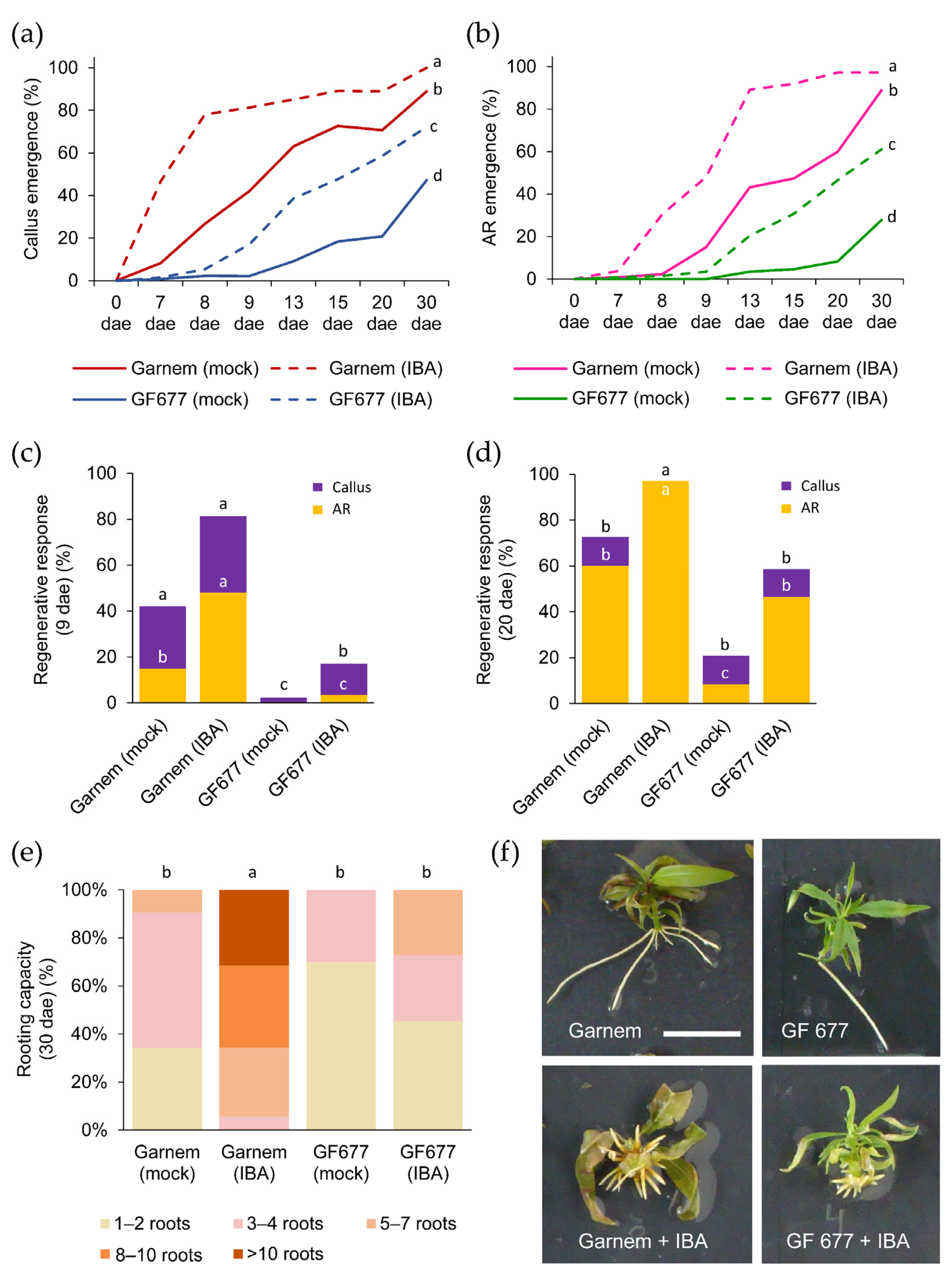
2.2. Regenerative Response of Garnem and GF 677 during In Vitro Rooting
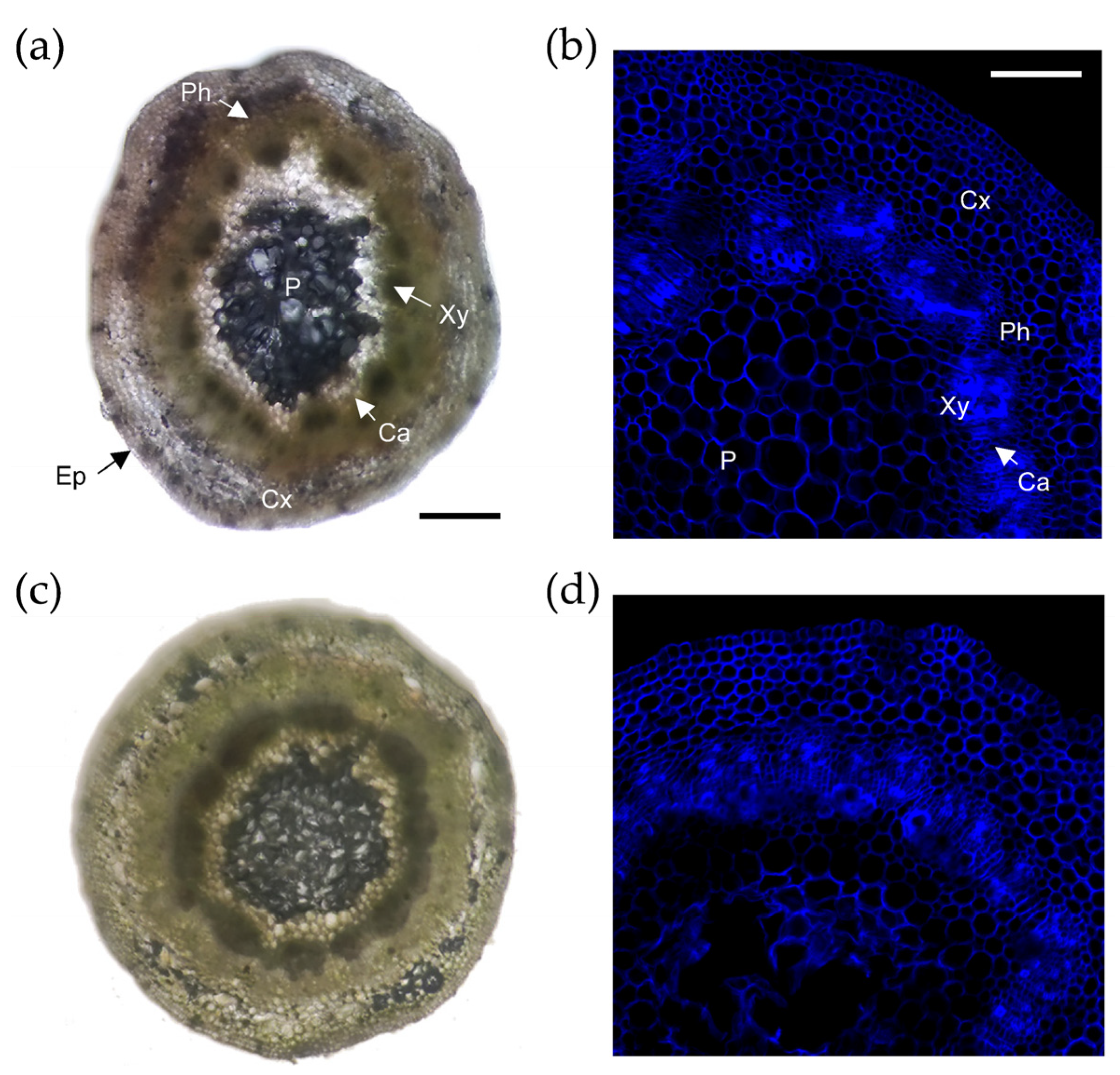
2.2.1. Histology
2.2.2. Auxin Treatment Improves Callus Formation and Enhances AR Number
2.2.3. Improved Rooting Performance on Prunus Rootstocks by an IBA Pulse
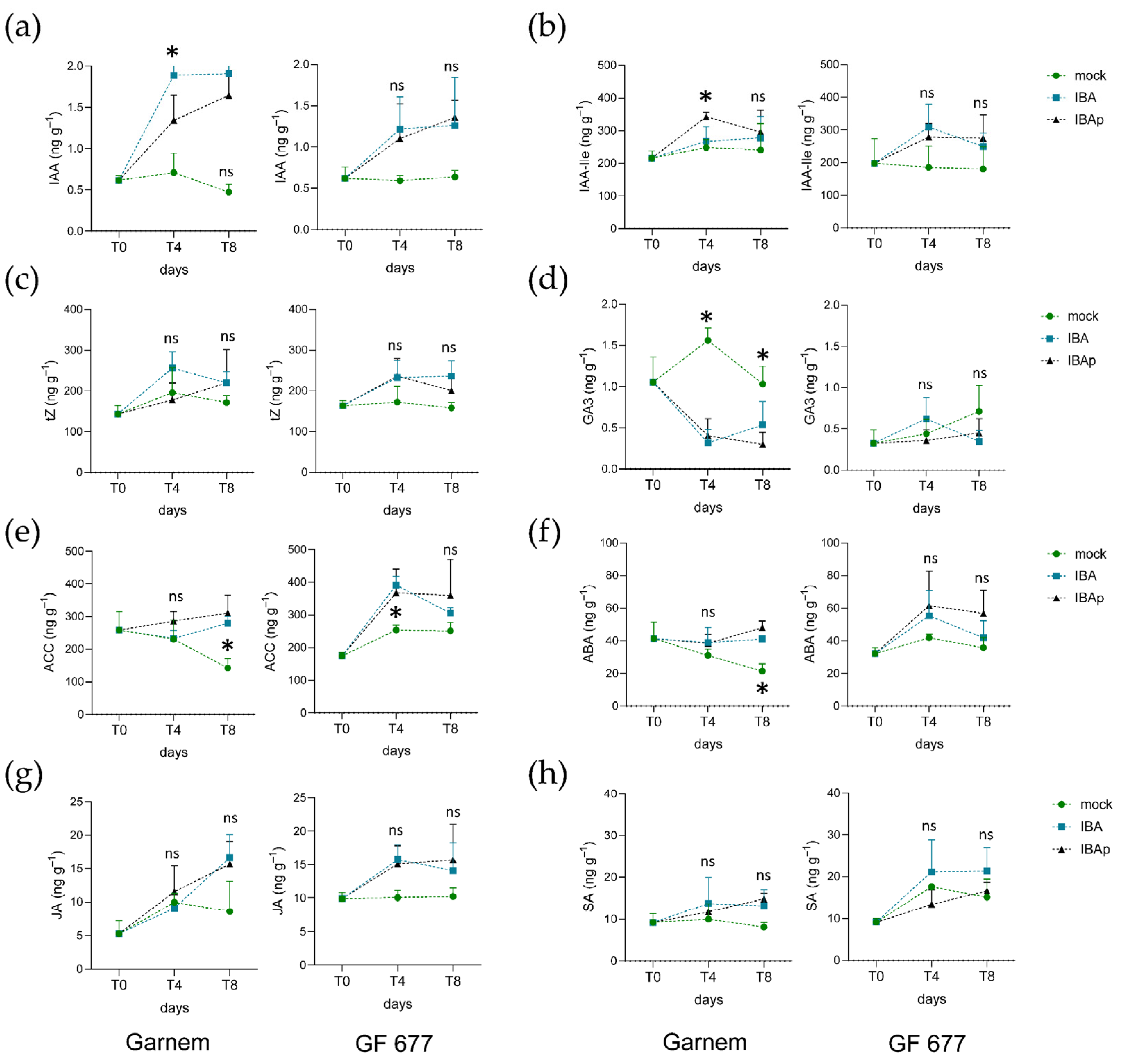
2.2.4. Hormone Profiling of Garnem and GF 677 Microcuttings during IBA-Induced Rooting
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions, and Sample Collection
4.1.1. Hydroponic Culture of Hardwood Cuttings
4.1.2. In Vitro Culture of Garnem and GF 677 Microcuttings
4.1.3. Direct Rooting on Substrate
4.1.4. Sample Collection
4.2. Histology
4.3. Picture, Image Processing, and Parameter Measurement
4.4. Phytohormone Extraction and Analysis
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Osmont, K.S.; Sibout, R.; Hardtke, C.S. Hidden branches: Developments in root system architecture. Annu. Rev. Plant Biol. 2007, 58, 93–113. [Google Scholar] [CrossRef]
- da Costa, C.T.; de Almeida, M.R.; Ruedell, C.M.; Schwambach, J.; Maraschin, F.S.; Fett-Neto, A.G. When stress and development go hand in hand: Main hormonal controls of adventitious rooting in cuttings. Front. Plant Sci. 2013, 4, 133. [Google Scholar] [CrossRef] [Green Version]
- Druege, U.; Hilo, A.; Pérez-Pérez, J.M.; Klopotek, Y.; Acosta, M.; Shahinnia, F.; Zerche, S.; Franken, P.; Hajirezaei, M.R. Molecular and physiological control of adventitious rooting in cuttings: Phytohormone action meets resource allocation. Ann. Bot. 2019, 123, 929–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielsa, B.; Hewitt, S.; Reyes-Chin-Wo, S.; Dhingra, A.; Rubio-Cabetas, M.J. Identification of water use efficiency related genes in “Garnem” almond-peach rootstock using time-course transcriptome analysis. PLoS ONE 2018, 13, e0205493. [Google Scholar] [CrossRef] [PubMed]
- Bastias, A.; Oviedo, K.; Almada, R.; Correa, F.; Sagredo, B. Identifying and validating housekeeping hybrid Prunus spp. Genes for root gene-expression studies. PLoS ONE 2020, 15, e0228403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batlle, I.; Iglesias, I.; Cantin, C.M.; Badenes, M.L.; Rios, G.; Ruiz, D.; Dicenta, F.; Egea, J.; López-Corrales, M.; Guerra, E.; et al. Frutales de hueso y pepita. In Influencia del Cambio Climático en la Mejora Genética de Plantas; Sociedad Española de Ciencias Hortícolas: Madrid, Spain, 2018; pp. 79–130. [Google Scholar]
- FAOSTAT. Food and Agricultural Organization. Available online: https://www.fao.org/faostat/en/ (accessed on 9 March 2022).
- Moreno, M.Á.; Gogorcena, Y.; Pinochet, J. Mejora y selección de patrones Prunus tolerantes a estreses abióticos. In La Adaptación al Ambiente y los Estreses Abióticos en la Mejora Vegetal; Junta de Andalucía, Dirección General de Planificación y Análisis de Mercados, Servicio de Publicaciones y Divulgación: Sevilla, Spain, 2008; pp. 451–475. ISBN 978-84-8474-248-2. [Google Scholar]
- Felipe, A.J. “Felinem”, “Garnem” and “Monegro” almond × peach hybrid rootstocks. HortScience 2009, 44, 196–197. [Google Scholar] [CrossRef] [Green Version]
- Mestre, L.; Reig, G.; Betrán, J.A.; Pinochet, J.; Moreno, M.Á. Influence of peach-almond hybrids and plum-based rootstocks on mineral nutrition and yield characteristics of “Big Top” nectarine in replant and heavy-calcareous soil conditions. Sci. Hortic. 2015, 192, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Font i Forcada, C.; Reig, G.; Mestre, L.; Mignard, P.; Betrán, J.Á.; Moreno, M.Á. Scion × rootstock response on production, mineral composition and fruit quality under heavy-calcareous soil and hot climate. Agronomy 2020, 10, 1159. [Google Scholar] [CrossRef]
- Reighard, G.L.; Beckman, T.; Belding, R.; Black, P.; Byers, P.; Cline, J.; Cowgill, W.; Godin, R.; Johnson, R.S.; Kamas, J.; et al. Six-Year Performance of 14 Prunus Rootstocks at 11 Sites in the 2001 NC-140 Peach Trial. J. Am. Pomol. Soc. 2011, 65, 26–41. [Google Scholar]
- Rubio-Cabetas, M.J. Estudio de las Posibilidades de Hibridación en el Genero Prunus Para la Mejora Genética de Patrones. Available online: https://www.tesisenred.net/handle/10803/8358#page=1 (accessed on 9 March 2022).
- Font i Forcada, C.; Gogorcena, Y.; Moreno, M.Á. Agronomical and fruit quality traits of two peach cultivars on peach-almond hybrid rootstocks growing on Mediterranean conditions. Sci. Hortic. 2012, 140, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Moreno, M.A.; Cambra, R. Adarcias: An almond x peach hybrid rootstock. HortScience 1994, 29, 925. [Google Scholar] [CrossRef]
- Moreno, M.; Tabuenca, M.; Cambra, R. Performance of Adafuel and Adarcias as peach rootstocks. HortScience 1994, 29, 1271–1273. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, V.J.; Rubio, M.; Trainotti, L.; Verde, I.; Bonghi, C.; Martínez-Gómez, P. Prunus transcription factors: Breeding perspectives. Front. Plant Sci. 2015, 6, 443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blythe, E.K.; Sibley, J.L.; Tilt, K.M.; Ruter, J.M. Methods of auxin application in cutting propagation: A review of 70 years of scientific discovery and commercial practice. J. Environ. Hortic. 2007, 25, 166–185. [Google Scholar] [CrossRef]
- Cambra, R. ‘Adafuel’, an almond × peach hybrid rootstock. HortScience 1990, 25, 584. [Google Scholar] [CrossRef] [Green Version]
- Selby, C.; Kennedy, S.J.; Harvey, B.M.R. Adventitious root formation in hypocotyl cuttings of Picea sitchensis (Bong.) Carr. The influence of plant growth regulators. New Phytol. 1992, 120, 453–457. [Google Scholar] [CrossRef]
- Agulló-Antón, M.Á.; Ferrández-Ayela, A.; Fernández-García, N.; Nicolás, C.; Albacete, A.; Pérez-Alfocea, F.; Sánchez-Bravo, J.; Pérez-Pérez, J.M.; Acosta, M. Early steps of adventitious rooting: Morphology, hormonal profiling and carbohydrate turnover in carnation stem cuttings. Physiol. Plant. 2014, 150, 446–462. [Google Scholar] [CrossRef] [PubMed]
- Cantabella, D.; Teixidó, N.; Segarra, G.; Torres, R.; Casanovas, M.; Dolcet-Sanjuan, R. Rhizosphere microorganisms enhance in vitro root and plantlet development of Pyrus and Prunus rootstocks. Planta 2021, 253, 78. [Google Scholar] [CrossRef] [PubMed]
- Gainza, F.; Opazo, I.; Guajardo, V.; Meza, P.; Ortiz, M.; Pinochet, J.; Muñoz, C. Rootstock breeding in Prunus species: Ongoing efforts and new challenges. Chil. J. Agric. Res. 2015, 75, 6–16. [Google Scholar] [CrossRef] [Green Version]
- Mayer, N.A.; Ueno, B.; Reighard, G.L. Selection of Prunus mume as rootstocks for peaches on PTSL site. Acta Hortic. 2015, 1084, 89–96. [Google Scholar] [CrossRef]
- Moreno, M.A. Breeding and selection of Prunus rootstocks at the Aula Dei Experimental Station, Zaragoza, Spain. Acta Hortic. 2004, 658, 519–528. [Google Scholar] [CrossRef]
- Eliyahu, A.; Duman, Z.; Sherf, S.; Genin, O.; Cinnamon, Y.; Abu-Abied, M.; Weinstain, R.; Dag, A.; Sadot, E. Vegetative propagation of elite Eucalyptus clones as food source for honeybees (Apis mellifera); adventitious roots versus callus formation. Isr. J. Plant Sci. 2020, 67, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Daskalakis, I.; Biniari, K.; Bouza, D.; Stavrakaki, M. The effect that indolebutyric acid (IBA) and position of cane segment have on the rooting of cuttings from grapevine rootstocks and from Cabernet franc (Vitis vinifera L.) under conditions of a hydroponic culture system. Sci. Hortic. 2018, 227, 79–84. [Google Scholar] [CrossRef]
- Shiozaki, S.; Makibuchi, M.; Ogata, T. Indole-3-acetic acid, polyamines, and phenols in hardwood cuttings of recalcitrant-to-root wild grapes native to east asia: Vitis davidii and Vitis kiusiana. J. Bot. 2013, 2013, 819531. [Google Scholar]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magyar-Tábori, K.; Dobránszky, J.; Jámbor-Benczúr, E.; Lazányi, J.; Szalai, J.; Ferenczy, A. Effects of indole-3-butyric acid levels and activated charcoal on rooting of in vitro shoots of apple rootstocks. Int. J. Hortic. Sci. 2002, 8, 25–28. [Google Scholar] [CrossRef] [Green Version]
- Massai, R.; Loreti, F. Preliminary observations on nine peach rootstocks grown in a replant soil. Acta Hortic. 2004, 658, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Gonin, M.; Bergougnoux, V.; Nguyen, T.D.; Gantet, P.; Champion, A. What makes adventitious roots? Plants 2019, 8, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vielba, J.M.; Vidal, N.; Carmen San José, M.; Rico, S.; Sánchez, C. Recent advances in adventitious root formation in Chestnut. Plants 2020, 9, 1543. [Google Scholar] [CrossRef] [PubMed]
- Druege, U.; Franken, P.; Hajirezaei, M.R. Plant hormone homeostasis, signaling, and function during adventitious root formation in cuttings. Front. Plant Sci. 2016, 7, 381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cano, A.; Sánchez-García, A.B.; Albacete, A.; González-Bayón, R.; Justamante, M.S.; Ibáñez, S.; Acosta, M.; Pérez-Pérez, J.M. Enhanced conjugation of auxin by GH3 enzymes leads to poor adventitious rooting in carnation stem cuttings. Front. Plant Sci. 2018, 9, 566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Tahir, M.M.; Wu, T.; Xie, L.; Zhang, X.; Mao, J.; Ayyoub, A.; Xing, L.; Zhang, D.; Shao, Y. Transcriptome analysis reveals multiple genes and complex hormonal-mediated interactions with PEG during adventitious root formation in apple. Int. J. Mol. Sci. 2022, 23, 976. [Google Scholar] [CrossRef] [PubMed]
- Bellini, C.; Pacurar, D.I.; Perrone, I. Adventitious roots and lateral roots: Similarities and differences. Annu. Rev. Plant Biol. 2014, 65, 639–666. [Google Scholar] [CrossRef] [PubMed]
- De Klerk, G.J.; Van Der Krieken, W.; De Jong, J.C. Review: The formation of adventitious roots: New concepts, new possibilities. In Vitro Cell. Dev. Biol. 1999, 35, 189–199. [Google Scholar] [CrossRef]
- Quambusch, M.; Gruß, S.; Pscherer, T.; Winkelmann, T.; Bartsch, M. Improved in vitro rooting of Prunus avium microshoots using a dark treatment and an auxin pulse. Sci. Hortic. 2017, 220, 52–56. [Google Scholar] [CrossRef]
- Jawahir, V.; Zolman, B.K. Long chain acyl CoA synthetase 4 catalyzes the first step in peroxisomal indole-3-butyric acid to IAA conversion. Plant Physiol. 2021, 185, 120–136. [Google Scholar] [CrossRef] [PubMed]
- Li, S.W.; Shi, R.F.; Leng, Y.; Zhou, Y. Transcriptomic analysis reveals the gene expression profile that specifically responds to IBA during adventitious rooting in mung bean seedlings. BMC Genom. 2016, 17, 43. [Google Scholar] [CrossRef] [Green Version]
- Fattorini, L.; Veloccia, A.; Della Rovere, F.; D’Angeli, S.; Falasca, G.; Altamura, M.M. Indole-3-butyric acid promotes adventitious rooting in Arabidopsis thaliana thin cell layers by conversion into indole-3-acetic acid and stimulation of anthranilate synthase activity. BMC Plant Biol. 2017, 17, 121. [Google Scholar] [CrossRef] [Green Version]
- Arismendi, M.J.; Almada, R.; Pimentel, P.; Bastias, A.; Salvatierra, A.; Rojas, P.; Hinrichsen, P.; Pinto, M.; Di Genova, A.; Travisany, D.; et al. Transcriptome sequencing of Prunus sp. rootstocks roots to identify candidate genes involved in the response to root hypoxia. Tree Genet. Genomes 2015, 11, 11. [Google Scholar] [CrossRef]
- Alaguero-Cordovilla, A.; Gran-Gómez, F.J.; Jadczak, P.; Mhimdi, M.; Ibáñez, S.; Bres, C.; Just, D.; Rothan, C.; Pérez-Pérez, J.M. A quick protocol for the identification and characterization of early growth mutants in tomato. Plant Sci. 2020, 301, 110673. [Google Scholar] [CrossRef]
- Ursache, R.; Andersen, T.G.; Marhavý, P.; Geldner, N. A protocol for combining fluorescent proteins with histological stains for diverse cell wall components. Plant J. 2018, 93, 399–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Albacete, A.; Ghanem, M.E.; Martínez-Andújar, C.; Acosta, M.; Sánchez-Bravo, J.; Martínez, V.; Lutts, S.; Dodd, I.C.; Pérez-Alfocea, F. Hormonal changes in relation to biomass partitioning and shoot growth impairment in salinized tomato (Solanum lycopersicum L.) plants. J. Exp. Bot. 2008, 59, 4119–4131. [Google Scholar] [CrossRef] [PubMed]
- Großkinsky, D.K.; Albacete, A.; Jammer, A.; Krbez, P.; Van Der Graaff, E.; Pfeifhofer, H.; Roitsch, T. A rapid phytohormone and phytoalexin screening method for physiological phenotyping. Mol. Plant 2014, 7, 1053–1056. [Google Scholar] [CrossRef] [Green Version]
- Aguinis, H.; Gottfredson, R.K.; Joo, H. Best-practice recommendations for defining, identifying, and handling outliers. Organ. Res. Methods 2013, 16, 270–301. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rootstock Name | Species | Origin |
|---|---|---|
| Adafuel | P. dulcis x P. persica | CSIC, Spain |
| Adarcias | P. dulcis x P. persica | CSIC, Spain |
| Cadaman | P. persica x P. davidiana | IFGO, Hungary and INRA, France |
| Garnem | P. dulcis x P. persica | CITA, Spain |
| GF 677 | P. dulcis x P. persica | INRA, France |
| Genotype (Treatment) | Callus Emergence | AR Emergence |
|---|---|---|
| Garnem (mock) | 9.7 | 15.7 |
| Garnem (IBA) | 6.3 | 8.9 |
| GF 677 (mock) | >30 | >30 |
| GF 677 (IBA) | 12.7 | 16.3 |
| Genotype (Treatment) | Callus Emergence | AR Emergence |
|---|---|---|
| Garnem (mock) | 7.6 | 13.5 |
| Garnem (IBAp) | 6.7 | 7.5 |
| GF 677 (mock) | >20 | >20 |
| GF 677 (IBAp) | 11.9 | 8.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Justamante, M.S.; Mhimdi, M.; Molina-Pérez, M.; Albacete, A.; Moreno, M.Á.; Mataix, I.; Pérez-Pérez, J.M. Effects of Auxin (Indole-3-butyric Acid) on Adventitious Root Formation in Peach-Based Prunus Rootstocks. Plants 2022, 11, 913. https://doi.org/10.3390/plants11070913
Justamante MS, Mhimdi M, Molina-Pérez M, Albacete A, Moreno MÁ, Mataix I, Pérez-Pérez JM. Effects of Auxin (Indole-3-butyric Acid) on Adventitious Root Formation in Peach-Based Prunus Rootstocks. Plants. 2022; 11(7):913. https://doi.org/10.3390/plants11070913
Chicago/Turabian StyleJustamante, María Salud, Mariem Mhimdi, Marta Molina-Pérez, Alfonso Albacete, María Ángeles Moreno, Inés Mataix, and José Manuel Pérez-Pérez. 2022. "Effects of Auxin (Indole-3-butyric Acid) on Adventitious Root Formation in Peach-Based Prunus Rootstocks" Plants 11, no. 7: 913. https://doi.org/10.3390/plants11070913
APA StyleJustamante, M. S., Mhimdi, M., Molina-Pérez, M., Albacete, A., Moreno, M. Á., Mataix, I., & Pérez-Pérez, J. M. (2022). Effects of Auxin (Indole-3-butyric Acid) on Adventitious Root Formation in Peach-Based Prunus Rootstocks. Plants, 11(7), 913. https://doi.org/10.3390/plants11070913