
Characteristics of NH4+ and NO3− Fluxes in Taxodium Roots under Different Nitrogen Treatments

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation
2.2. Experimental Design
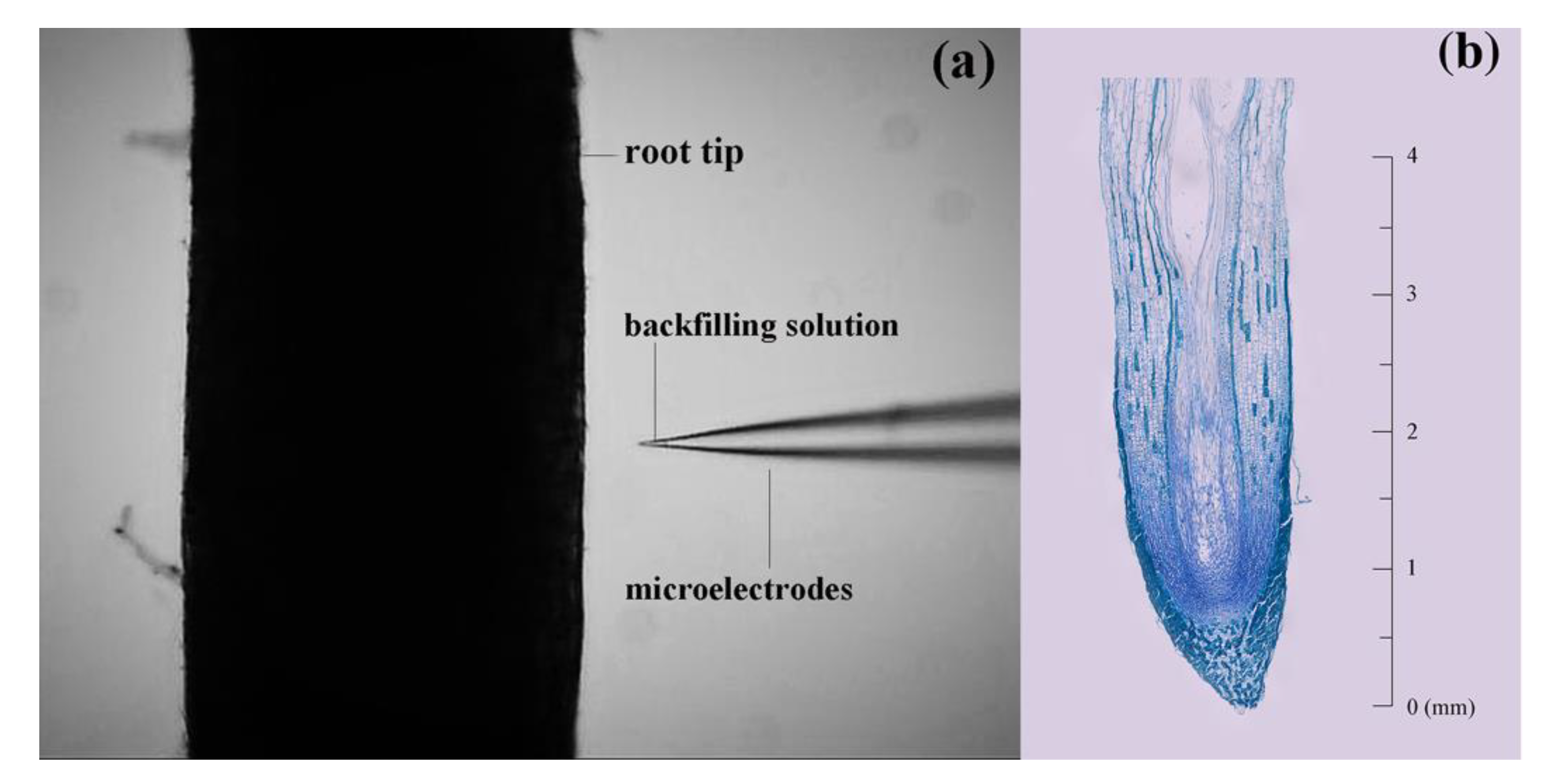
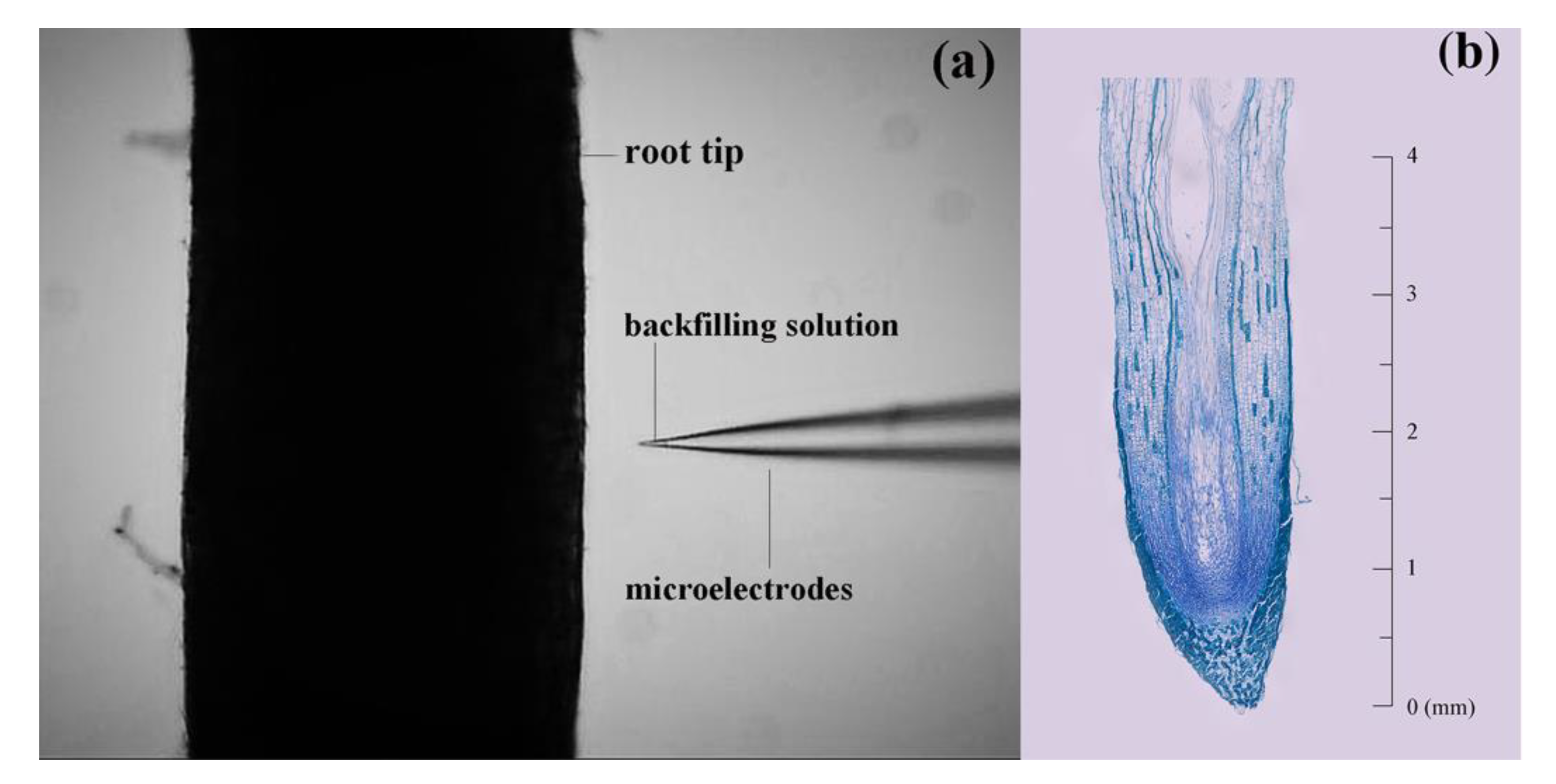
2.3. Measurement of NH4+, NO3− and H+ Fluxes
2.4. Determination of 15N Uptake and Enzyme Activities
2.5. Statistical Analysis
3. Results
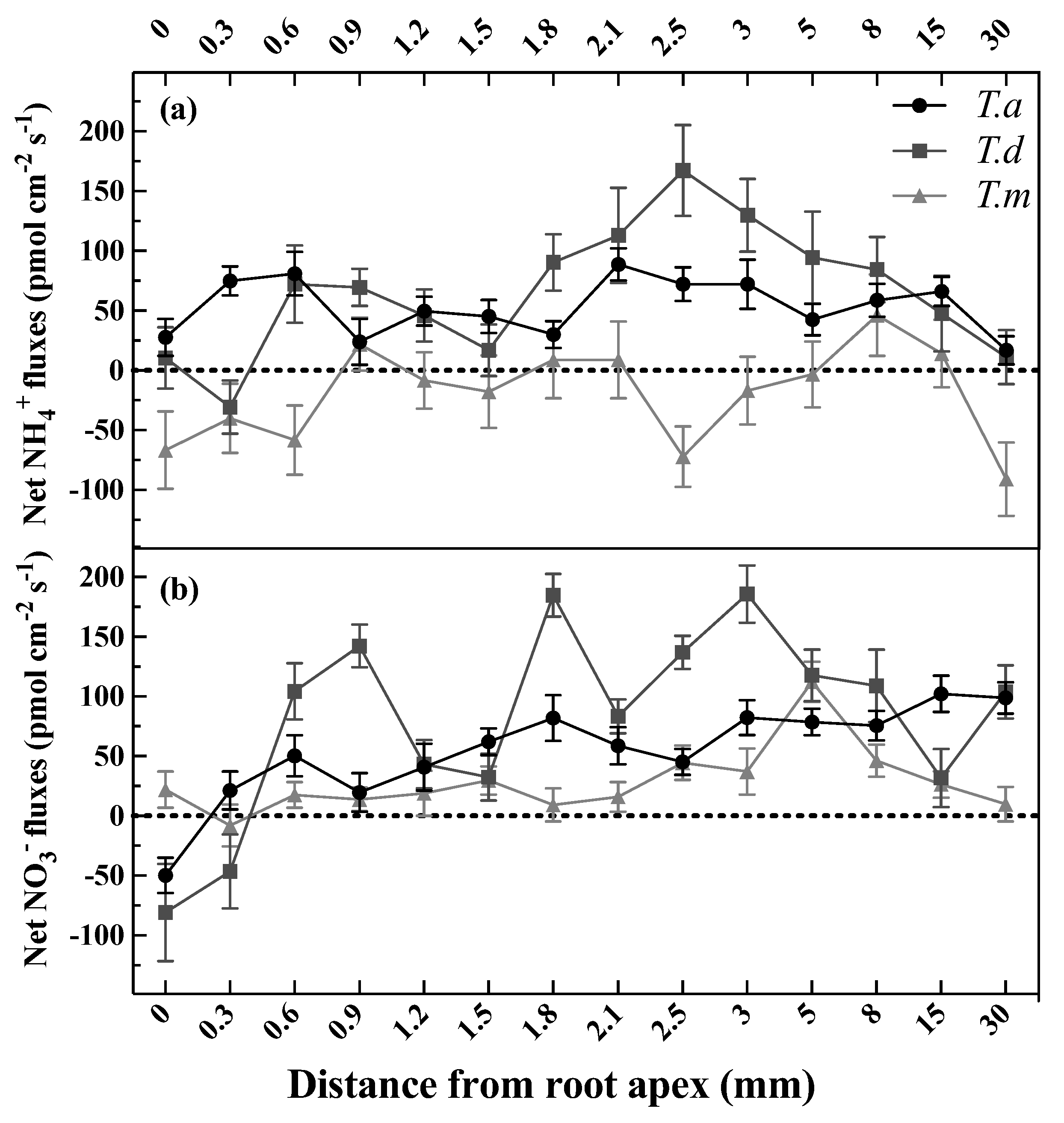
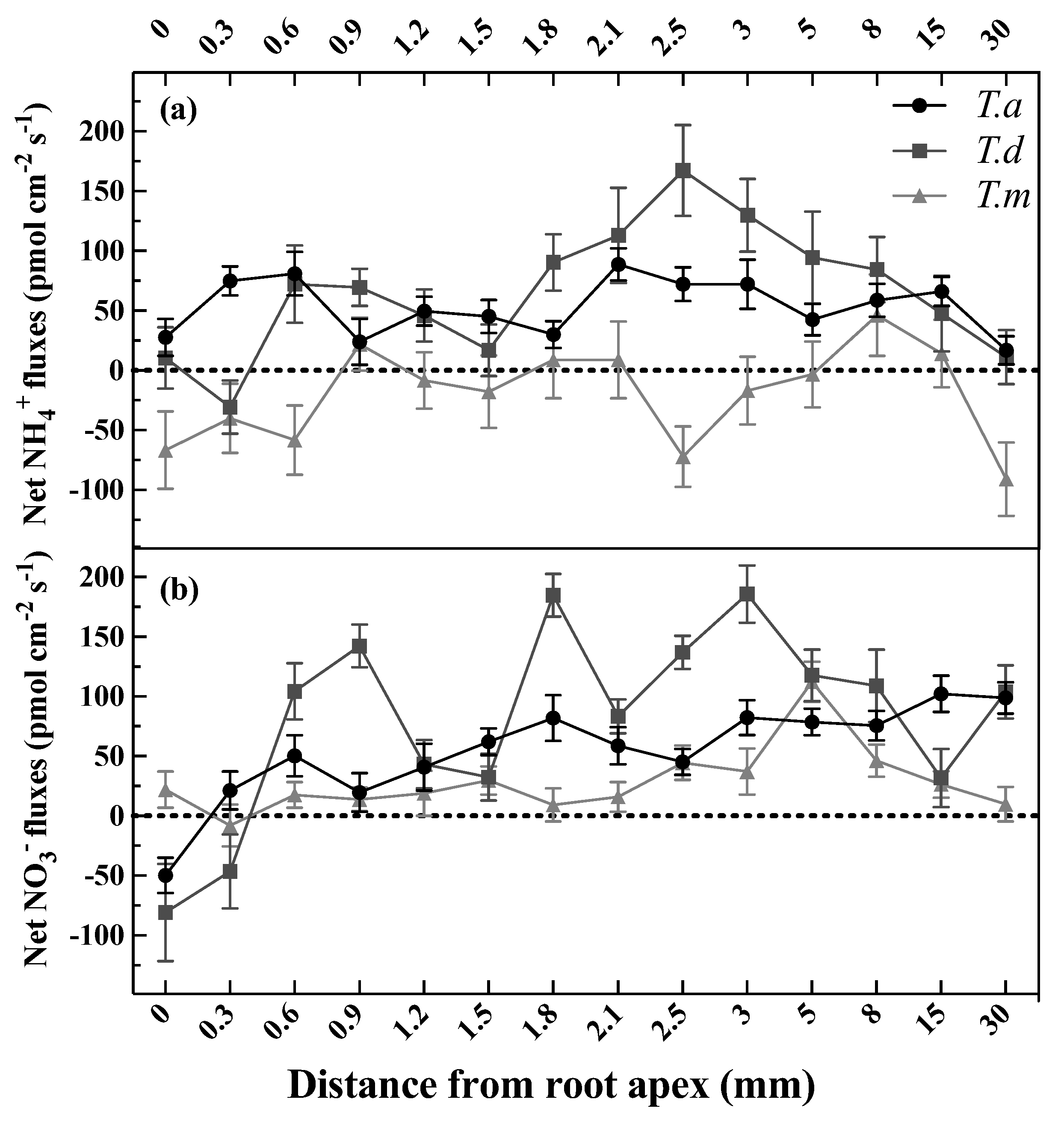
3.1. Net Fluxes of NH4+ and NO3− along the Root Tip
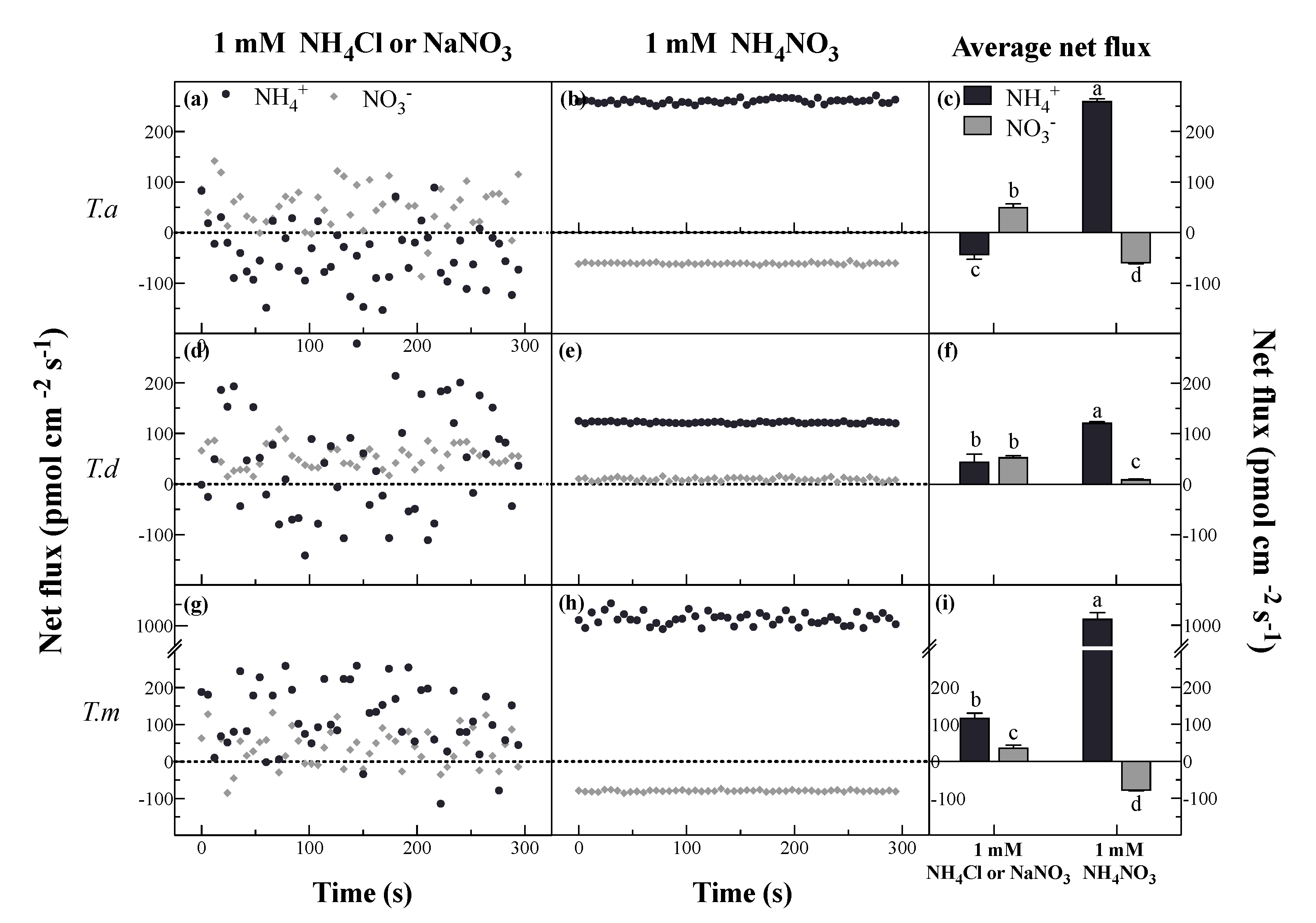
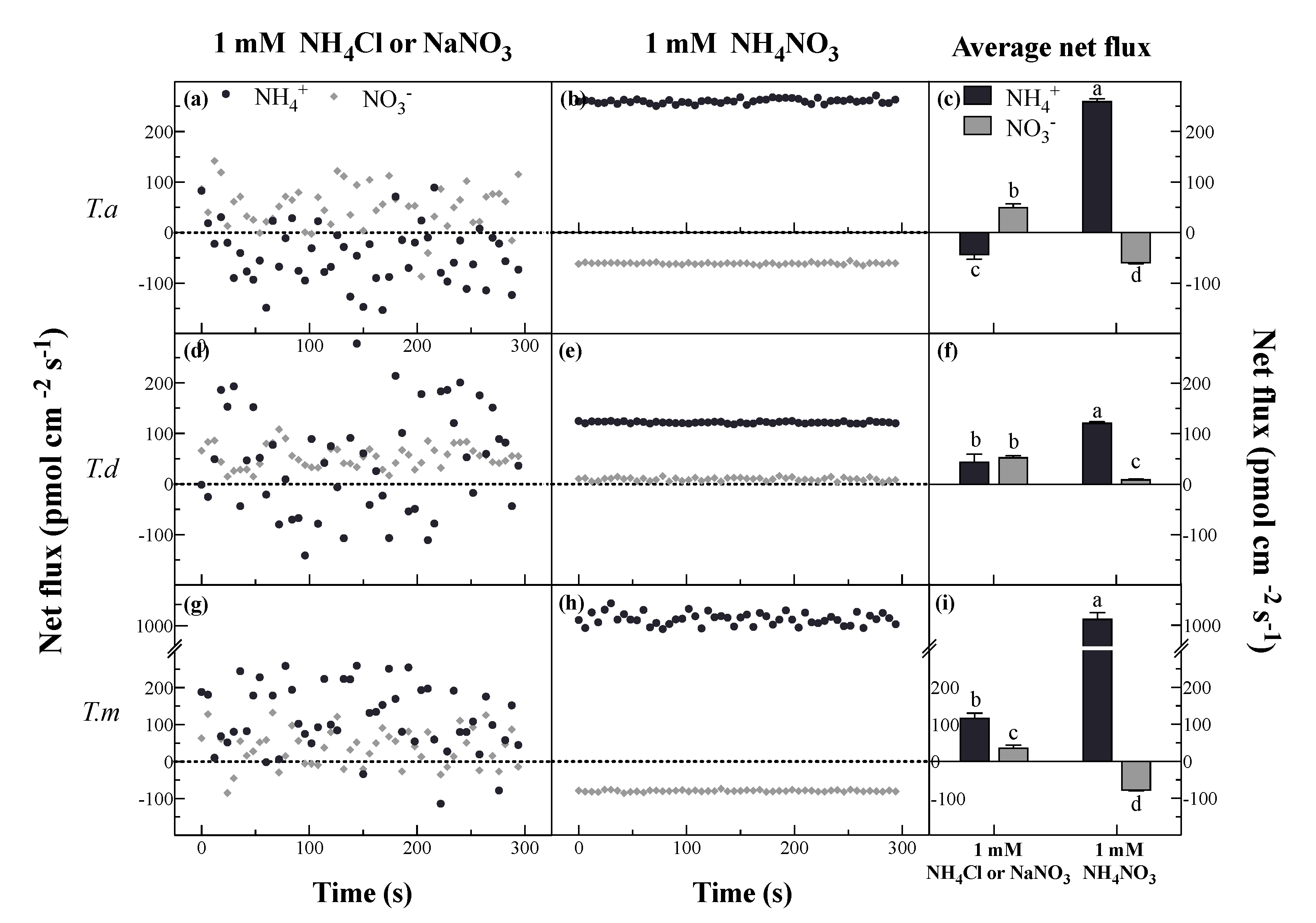
3.2. Net Fluxes of NH4+ and NO3− under Different N Forms
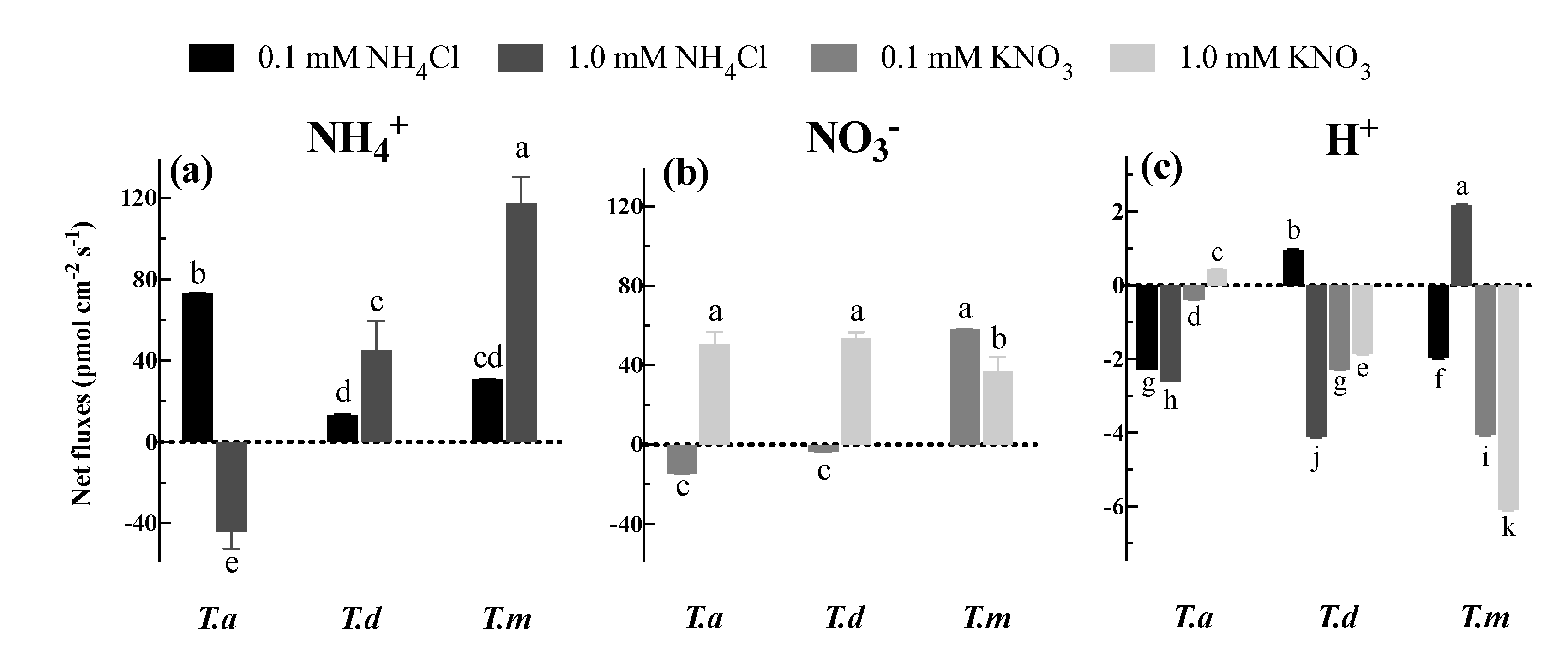
3.3. Net NH4+, NO3− and H+ Fluxes under Different N Concentrations
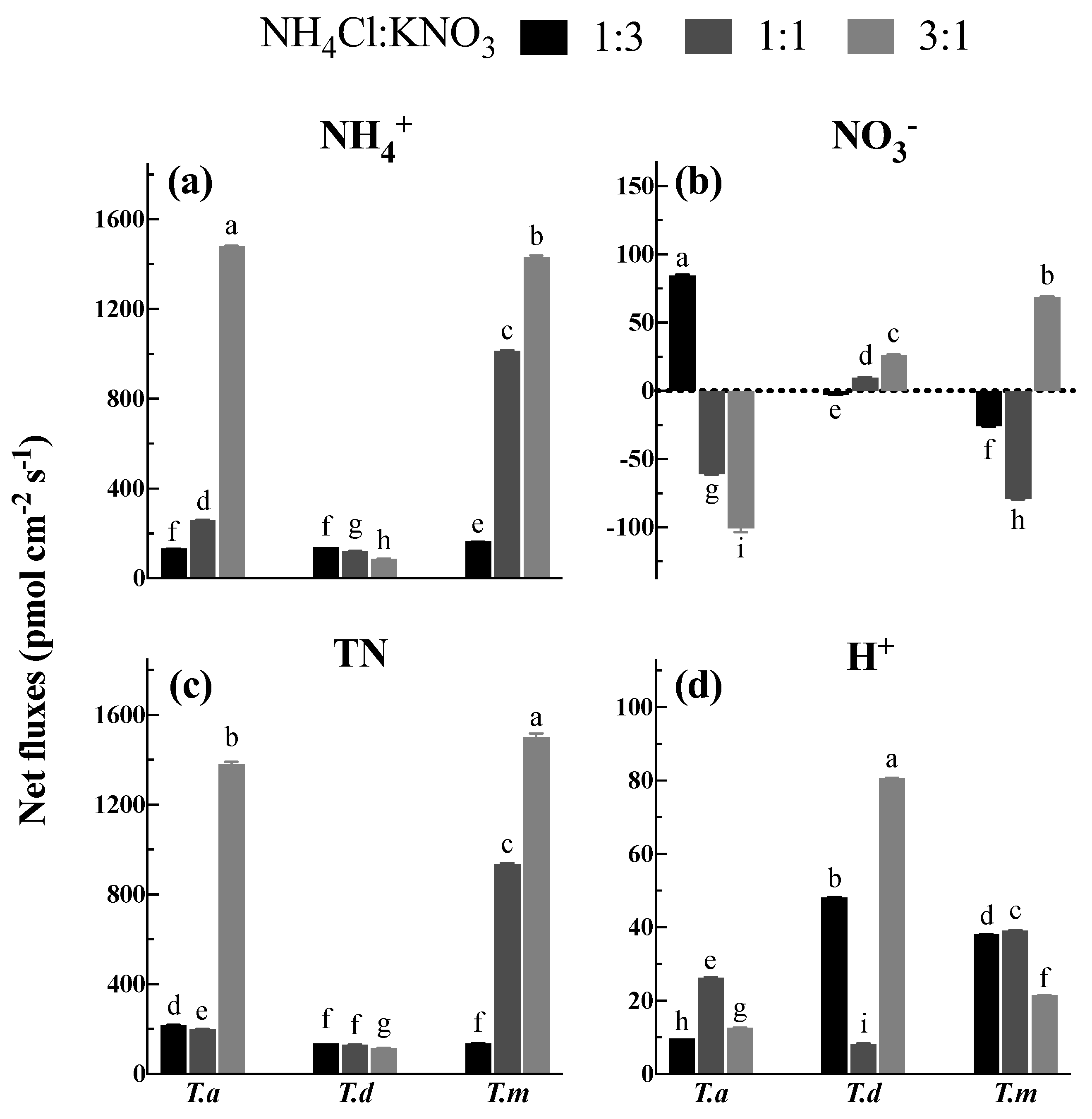
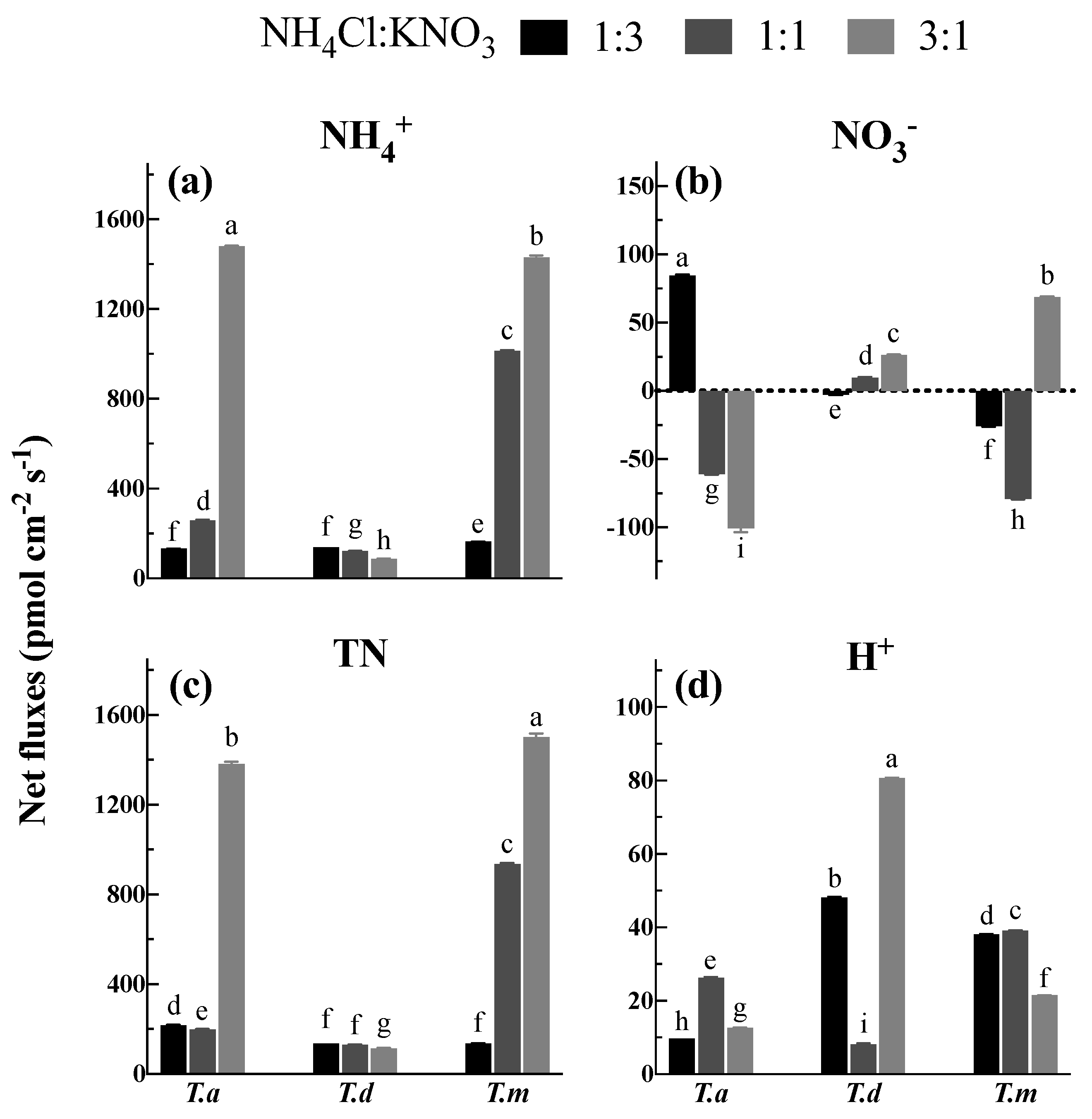
3.4. Net NH4+ and NO3− Fluxes under Different N Proportions
3.5. 15N AT%, Ndff% and Enzyme Activities in the Roots of T. distichum
4. Discussion
4.1. Spatial Variability of Net NH4+ and NO3− Fluxes along the Fine Roots
4.2. Net NH4+ and NO3− Fluxes under Single N Treatments
4.3. Net NH4+ and NO3− Fluxes under Mixed N Treatments
4.4. Net NH4+ and NO3− Fluxes Associated with H+
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruan, L.; Wei, K.; Wang, L.; Cheng, H.; Zhang, F.; Wu, L.; Bai, P.; Zhang, C. Characteristics of NH4+ and NO3− fluxes in tea (Camellia sinensis) roots measured by scanning ion-selective electrode technique. Sci. Rep. 2016, 6, 38370. [Google Scholar] [CrossRef] [PubMed]
- Colmer, T.D.; Bloom, A.J. A comparison of NH4+ and NO3– net fluxes along roots of rice and maize. Plant Cell Environ. 2010, 21, 240–246. [Google Scholar] [CrossRef]
- Hawkins, B.J.; Robbins, S.; Porter, R.B. Nitrogen uptake over entire root systems of tree seedlings. Tree Physiol. 2014, 34, 334–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Li, H.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.B. Nitrogen metabolism of two contrasting poplar species during acclimation to limiting nitrogen availability. J. Exp. Bot. 2013, 64, 4207–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Z.; Li, Y.; Meng, S.; Zhang, C. Net NH4+ and NO3− fluxes, and expression of NH4+ and NO3− transporter genes in roots of Populus simonii after acclimation to moderate salinity. Trees 2014, 28, 1813–1821. [Google Scholar] [CrossRef]
- Hawkins, B.J.; Boukcim, H.; Plassard, C. A comparison of ammonium, nitrate and proton net fluxes along seedling roots of Douglas-fir and lodgepole pine grown and measured with different inorganic nitrogen sources. Plant Cell Environ. 2008, 31, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Qin, J.; He, F.; Li, H.; Liu, T.; Polle, A.; Peng, C.; Luo, Z.B. Net fluxes of ammonium and nitrate in association with H+ fluxes in fine roots of Populus popularis. Planta 2013, 237, 919–931. [Google Scholar] [CrossRef]
- Alber, A.; Ehlting, B.; Ehlting, J.; Hawkins, B.; Rennenberg, H. Net NH4+ and NO3− flux, and expression of NH4+ and NO3− transporters in roots of Picea glauca. Trees 2012, 26, 1403–1411. [Google Scholar] [CrossRef]
- Zhu, Y.; Huang, X.; Hao, Y.; Su, W.; Liu, H.; Sun, G.; Chen, R.; Song, S. Ammonium transporter (BcAMT1.2) mediates the interaction of ammonium and nitrate in Brassica campestris. Front. Plant Sci. 2019, 10, 1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Y.; Qi, B.; Hao, Y.; Liu, H.; Sun, G.; Chen, R.; Song, S. Appropriate NH4+/NO3− ratio triggers plant growth and nutrient uptake of optimizing the pH value of nutrient solution. Front. Plant Sci. 2021, 12, 656144. [Google Scholar] [CrossRef]
- Tang, B.; Yin, C.; Yang, H.; Sun, Y.; Liu, Q. The coupling effects of water deficit and nitrogen supply on photosynthesis, WUE, and stable isotope composition in Picea asperata. Acta Physiol. Plant 2017, 39, 148. [Google Scholar] [CrossRef]
- Zhang, M.; Ding, M.; Xu, F.; Afzal, M.R.; Chen, X.; Zeng, H.; Yan, F.; Zhu, Y. Involvement of plasma membrane H+-ATPase in the ammonium-nutrition response of barley roots. J. Plant. Nutr. Soil Sci. 2018, 181, 878–885. [Google Scholar] [CrossRef]
- McClure, P.R.; Kochian, L.V.; Spanswick, R.M.; Shaff, J.E. Evidence for cotransport of nitrate and protons in maize roots: II. Measurement of NO3− and H+ fluxes with ion-selective microelectrodes. Plant Physiol. 1990, 93, 290–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, H.; Di, T.; Zhu, Y.; Subbarao, G.V. Transcriptional response of plasma membrane H+-ATPase genes to ammonium nutrition and its functional link to the release of biological nitrification inhibitors from sorghum roots. Plant Soil 2016, 398, 301–312. [Google Scholar] [CrossRef]
- Garnett, T.P.; Shabala, S.N.; Smethurst, P.J.; Newman, I.A. Kinetics of ammonium and nitrate uptake by eucalypt roots and associated proton fluxes measured using ion selective microelectrodes. Funct. Plant Biol. 2003, 30, 1165–1176. [Google Scholar] [CrossRef]
- Santi, S.; Locci, G.; Pinton, R.; Cesco, S.; Varanini, Z. Plasma membrane H+-ATPase in maize roots induced for NO3− uptake. Plant Physiol. 1995, 109, 1277–1283. [Google Scholar] [CrossRef] [Green Version]
- Sorgona, A.; Lupini, A.; Mercati, F.; Di Dio, L.; Sunseri, F.; Abenavoli, M.R. Nitrate uptake along the maize primary root: An integrated physiological and molecular approach. Plant Cell Environ. 2011, 34, 1127–1140. [Google Scholar] [CrossRef]
- Stalter, R. Some ecological observations of Taxodium distichum (L.) Richard, in Delaware. Castanea 1981, 46, 154–161. [Google Scholar] [CrossRef]
- Rockwood, D.; Naidu, C.; Carter, D.; Rahmani, M.; Spriggs, T.; Lin, C.; Alker, G.; Isebrands, J.; Segrest, S. Short-rotation woody crops and phytoremediation: Opportunities for agroforestry? In New Vistas in Agroforestry; Springer: Berlin/Heidelberg, Germany, 2004; Volume 61, pp. 51–63. [Google Scholar]
- Abdelsalam, N.R.; Salem, M.Z.; Ali, H.M.; Mackled, M.I.; Mervat, E.-H.; Elshikh, M.S.; Hatamleh, A.A. Morphological, biochemical, molecular, and oil toxicity properties of Taxodium trees from different locations. Ind. Crops Prod. 2019, 139, 111515. [Google Scholar] [CrossRef]
- Zaher, A.M.; Lin, J.; Arai, M. Cytotoxic activity of abietane-type diterpenes isolated from Taxodium distichum against cancer cells adapted to nutrient-starved conditions. Nat. Prod. Commun. 2020, 15, 1–6. [Google Scholar] [CrossRef]
- Hua, J.; Han, L.; Wang, Z.; Gu, C.; Yin, Y. Morpho-anatomical and photosynthetic responses of Taxodium hybrid’Zhongshanshan’406 to prolonged flooding. Flora 2017, 231, 29–37. [Google Scholar] [CrossRef]
- Xuan, L.; Hua, J.; Zhang, F.; Wang, Z.; Pei, X.; Yang, Y.; Yin, Y.; Creech, D.L. Identification and functional analysis of ThADH1 and ThADH4 genes involved in tolerance to waterlogging stress in Taxodium hybrid ‘Zhongshanshan 406’. Genes 2021, 12, 225. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Yan, W.; Chen, J.; Shangguan, Z. Net ammonium and nitrate fluxes in wheat roots under different environmental conditions as assessed by scanning ion-selective electrode technique. Sci. Rep. 2014, 4, 7223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Hu, R.; Zheng, Z.; Yang, J.; Fan, H.; Deng, X.; Yao, W.; Wang, Q.; Peng, S.; Li, J. Soil bacterial community in the multiple cropping system increased grain yield within 40 cultivation years. Front. Plant Sci. 2021, 12, 804527. [Google Scholar] [CrossRef] [PubMed]
- Enstone, D.E.; Peterson, C.A.; Hallgren, S.W. Anatomy of seedling tap roots of loblolly pine (Pinus taeda L.). Trees 2001, 15, 98–111. [Google Scholar] [CrossRef]
- Fang, Y.Y.; Babourina, O.; Rengel, Z.; Yang, X.E.; Pu, P.M. Spatial distribution of ammonium and nitrate fluxes along roots of wetland plants. Plant Sci. 2007, 173, 240–246. [Google Scholar] [CrossRef]
- Li, Q.; Li, B.H.; Kronzucker, H.J.; Shi, W.M. Root growth inhibition by NH4+ in Arabidopsis is mediated by the root tip and is linked to NH4+ efflux and GMPase activity. Plant Cell Environ. 2010, 33, 1529–1542. [Google Scholar] [CrossRef]
- Tang, B.; Yin, C.; Liu, Q. Characteristics of ammonium and nitrate fluxes along the roots of Picea asperata. J. Plant Nutr. Soil Sci. 2019, 42, 772–782. [Google Scholar] [CrossRef]
- Zou, N.; Shi, W.; Hou, L.; Kronzucker, H.J.; Huang, L.; Gu, H.; Yang, Q.; Deng, G.; Yang, G. Superior growth, N uptake and NH4+ tolerance in the giant bamboo Phyllostachys edulis over the broad-leaved tree Castanopsis fargesii at elevated NH4+ may underlie community succession and favor the expansion of bamboo. Tree Physiol. 2020, 40, 1606–1622. [Google Scholar] [CrossRef]
- Lu, Y.; Deng, S.; Li, Z.; Wu, J.; Zhu, D.; Shi, W.; Zhou, J.; Fayyaz, P.; Luo, Z.B. Physiological characteristics and transcriptomic dissection in two root segments with contrasting net fluxes of ammonium and nitrate of poplar under low nitrogen availability. Plant Cell Physiol. 2022, 63, 30–44. [Google Scholar] [CrossRef] [PubMed]
- Di, D.W.; Sun, L.; Wang, M.; Wu, J.; Kronzucker, H.J.; Fang, S.; Chu, J.; Shi, W.; Li, G. WRKY46 promotes ammonium tolerance in Arabidopsis by repressing NUDX9 and indole-3-acetic acid-conjugating genes and by inhibiting ammonium efflux in the root elongation zone. New Phytol. 2021, 232, 190–207. [Google Scholar] [CrossRef] [PubMed]
- Mackown, C.T.; Jackson, W.A.; Volk, R.J. Restricted nitrate influx and reduction in corn seedlings exposed to ammonium. Plant Physiol. 1982, 69, 353–359. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.; Gerendás, J.; Härdter, R.; Sattelmacher, B. Effect of nitrogen form and root-zone pH on growth and nitrogen uptake of tea (Camellia sinensis) plants. Ann. Bot. 2007, 99, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinsinger, P.; Plassard, C.; Tang, C.; Jaillard, B. Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: A review. Plant Soil 2003, 248, 43–59. [Google Scholar] [CrossRef]
- Tang, C.; Drevon, J.; Jaillard, B.; Souche, G.; Hinsinger, P. Proton release of two genotypes of bean (Phaseolus vulgaris L.) as affected by N nutrition and P deficiency. Plant Soil 2004, 260, 59–68. [Google Scholar] [CrossRef]
- Esteban, R.; Ariz, I.; Cruz, C.; Moran, J.F. Review: Mechanisms of ammonium toxicity and the quest for tolerance. Plant Sci. 2016, 248, 92–101. [Google Scholar] [CrossRef] [Green Version]
- Ruan, J.; Ma, L.; Shi, Y.; Zhang, F. Effects of litter incorporation and nitrogen fertilization on the contents of extractable aluminium in the rhizosphere soil of tea plant (Camallia sinensis (L.) O. Kuntze). Plant Soil 2004, 263, 283–296. [Google Scholar] [CrossRef]
- Bar-Yosef, B.; Mattson, N.; Lieth, H. Effects of NH4+:NO3−:Urea ratio on cut roses yield, leaf nutrients content and proton efflux by roots in closed hydroponic system. Sci. Hortic. 2009, 122, 610–619. [Google Scholar] [CrossRef]
- Xu, J.; Fang, Y.; Tavakkoli, E.; Pan, X.; Liao, F.; Chen, W.; Guo, W. Preferential ammonium: Nitrate ratio of blueberry isregulated by nitrogen transport and reduction systems. Sci. Hortic. 2021, 288, 110345. [Google Scholar] [CrossRef]
- De Bona, F.D.; Schmidt, F.; Monteiro, F.A. Importance of the nitrogen source in the grass species Brachiaria brizantha responses to sulfur limitation. Plant Soil 2013, 373, 201–216. [Google Scholar] [CrossRef] [Green Version]
- Yin, S.; Liang, Y.; Gao, W.; Wang, J.; Jing, S.; Zhang, Y.; Liu, H. Influence of medium salt strength and nitrogen source on biomass and metabolite accumulation in adventitious root cultures of Pseudostellaria heterophylla. Acta Physiol. Plant 2013, 35, 2623–2628. [Google Scholar] [CrossRef]
- Miller, A.; Cramer, M. Root nitrogen acquisition and assimilation. Plant Soil 2005, 274, 1–36. [Google Scholar] [CrossRef]
- Duan, F.; Giehl, R.F.H.; Geldner, N.; Salt, D.E.; von Wiren, N. Root zone-specific localization of AMTs determines ammonium transport pathways and nitrogen allocation to shoots. PLoS Biol. 2018, 16, e2006024. [Google Scholar] [CrossRef] [PubMed]
- Britto, D.T.; Kronzucker, H.J. Futile cycling at the plasma membrane: A hallmark of low-affinity nutrient transport. Trends Plant Sci. 2006, 11, 529–534. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Root Biomass/g | 15N AT% | Ndff% |
|---|---|---|---|
| CK | 0.30 ± 0.01 bc | 0.37 ± 0.00 c | / |
| 1 mM 15NH4+ | 0.34 ± 0.02 ab | 0.63 ± 0.03 a | 2.65 ± 0.31 a |
| 1 mM 15NO3− | 0.26 ± 0.01 c | 0.48 ± 0.04 b | 1.16 ± 0.38 b |
| Treatments | NR Activity μmol h−1 mg−1 Protein | NiR Activity μmol h−1 mg−1 Protein | GS Activity μmol h−1 mg−1 Protein | GDH Activity μmol h−1 mg−1 Protein | GOGAT Activity μmol h−1 mg−1 Protein |
|---|---|---|---|---|---|
| CK | 0.58 ± 0.02 c | 2.43 ± 0.04 c | 0.33 ± 0.00 b | 1.04 ± 0.06 b | 0.78 ± 0.03 b |
| 1 mM 15NH4+ | 0.87 ± 0.02 b | 3.25 ± 0.08 a | 0.46 ± 0.22 a | 1.54 ± 0.08 a | 1.24 ± 0.03 a |
| 1 mM 15NO3− | 1.92 ± 0.04 a | 2.72 ± 0.01 b | 0.34 ± 0.01 b | 1.01 ± 0.04 b | 0.74 ± 0.01 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, S.; Hua, J.; Lu, Y.; Zhang, R.; Yin, Y. Characteristics of NH4+ and NO3− Fluxes in Taxodium Roots under Different Nitrogen Treatments. Plants 2022, 11, 894. https://doi.org/10.3390/plants11070894
Wu S, Hua J, Lu Y, Zhang R, Yin Y. Characteristics of NH4+ and NO3− Fluxes in Taxodium Roots under Different Nitrogen Treatments. Plants. 2022; 11(7):894. https://doi.org/10.3390/plants11070894
Chicago/Turabian StyleWu, Shuting, Jianfeng Hua, Yan Lu, Rui Zhang, and Yunlong Yin. 2022. "Characteristics of NH4+ and NO3− Fluxes in Taxodium Roots under Different Nitrogen Treatments" Plants 11, no. 7: 894. https://doi.org/10.3390/plants11070894
APA StyleWu, S., Hua, J., Lu, Y., Zhang, R., & Yin, Y. (2022). Characteristics of NH4+ and NO3− Fluxes in Taxodium Roots under Different Nitrogen Treatments. Plants, 11(7), 894. https://doi.org/10.3390/plants11070894