
Growth Optimization and Secondary Metabolites Evaluation of Anabaena variabilis for Acetylcholinesterase Inhibition Activity

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cyanobacteria Isolates
2.2. Molecular Identification and Phylogenetic Analysis of Anabaena variabilis Based on 16S Ribosomal RNA Analysis
2.3. Effect of Different Nutrient Media on Growth of the Isolated Cyanobacteria
2.4. Screening the Effect of Modified Navicula Nutrient Medium Components on Biomass Production of Anabaena variabilis by Plackett–Burman
2.5. Verification Experiment
2.6. Biomass Harvesting
2.7. Extraction with Methylene Chloride: Methanol (1:1) v/v
2.8. Fractionation of Methylene Chloride: Methanol Crude Extract
2.9. In Vitro Acetylcholinesterase Activity Assay
2.10. Analysis of the Chemical Constituents of Methylene Chloride: Methanol Extract
2.11. Molecular Docking Study of Certain Biochemical Constituents
2.12. Statistical Analysis
3. Results
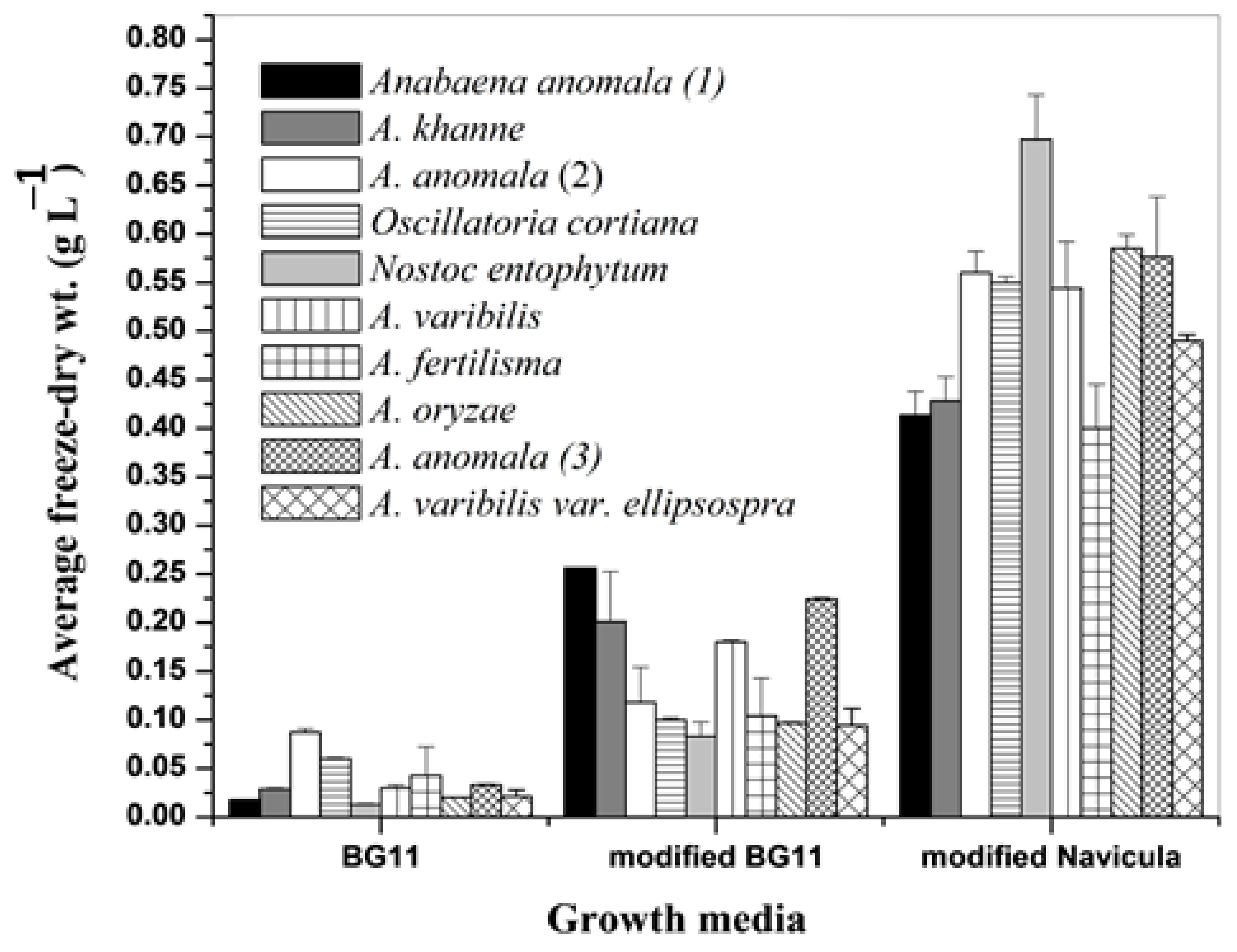
3.1. Growth of Different Cyanobacteria
3.2. Inhibition of Acetylcholinesterase Activity by Tested Cyanobacteria Crude Extracts
3.3. Screening the Effect of Modified Navicula Nutrient Medium Component on Biomass Production of A. variabilis by Plackett–Burman Design
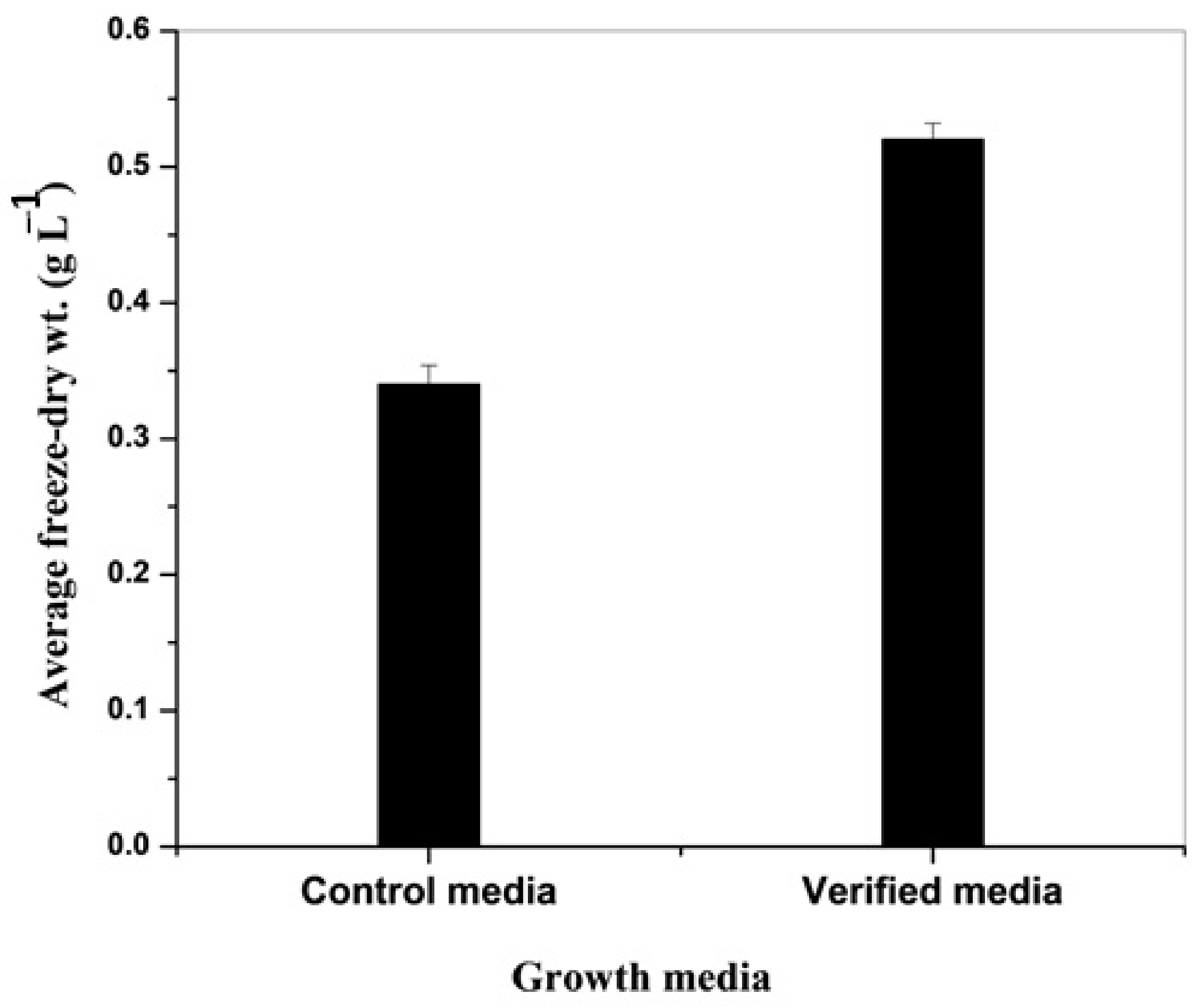
3.4. Verification Experiments
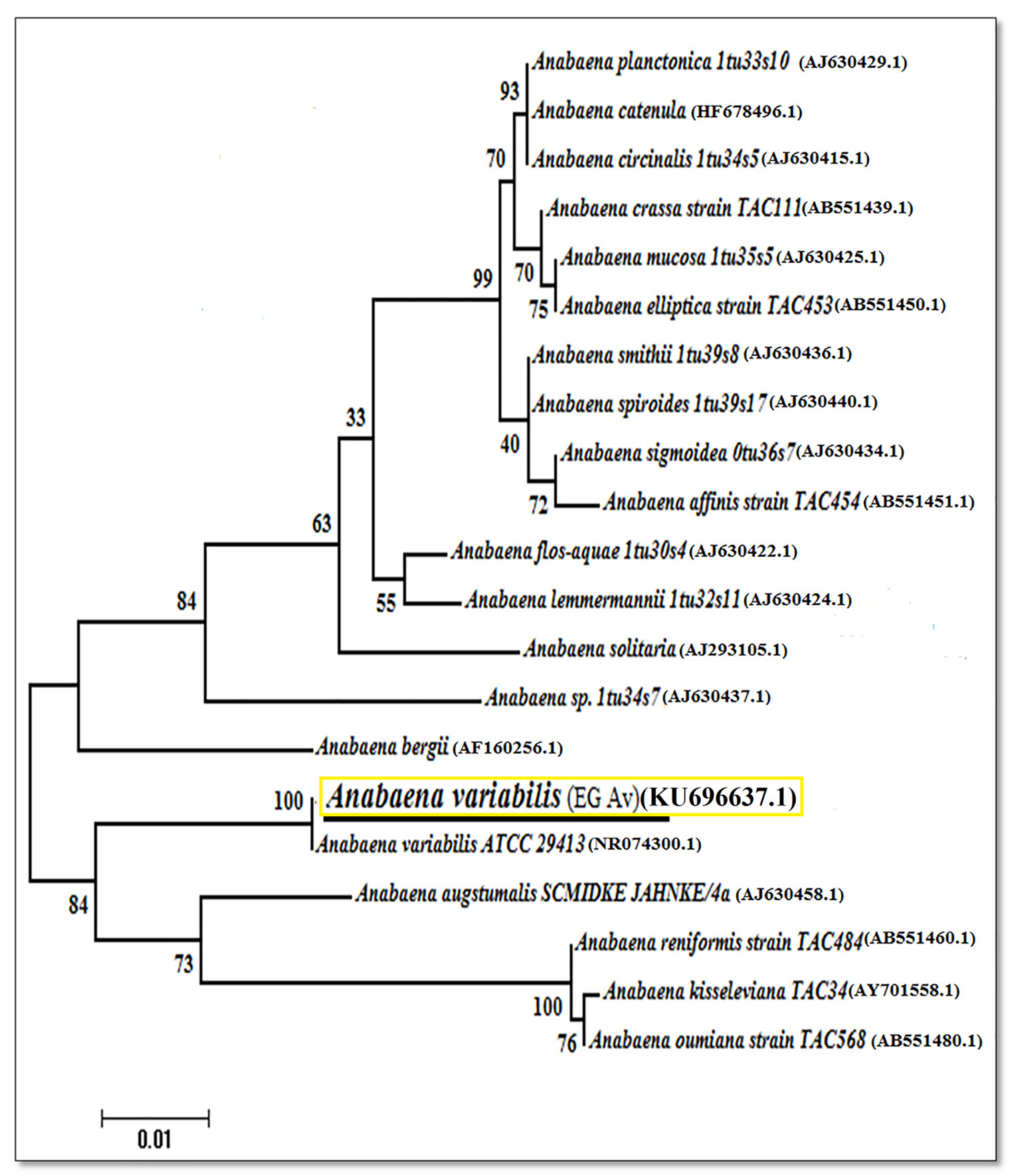
3.5. Phylogenetic Analysis and Placement of Anabaena variabilis
3.6. The AChE Inhibitory Activity of Ten TLC Fractions
3.7. GC/MS Analyses of the Fraction F7
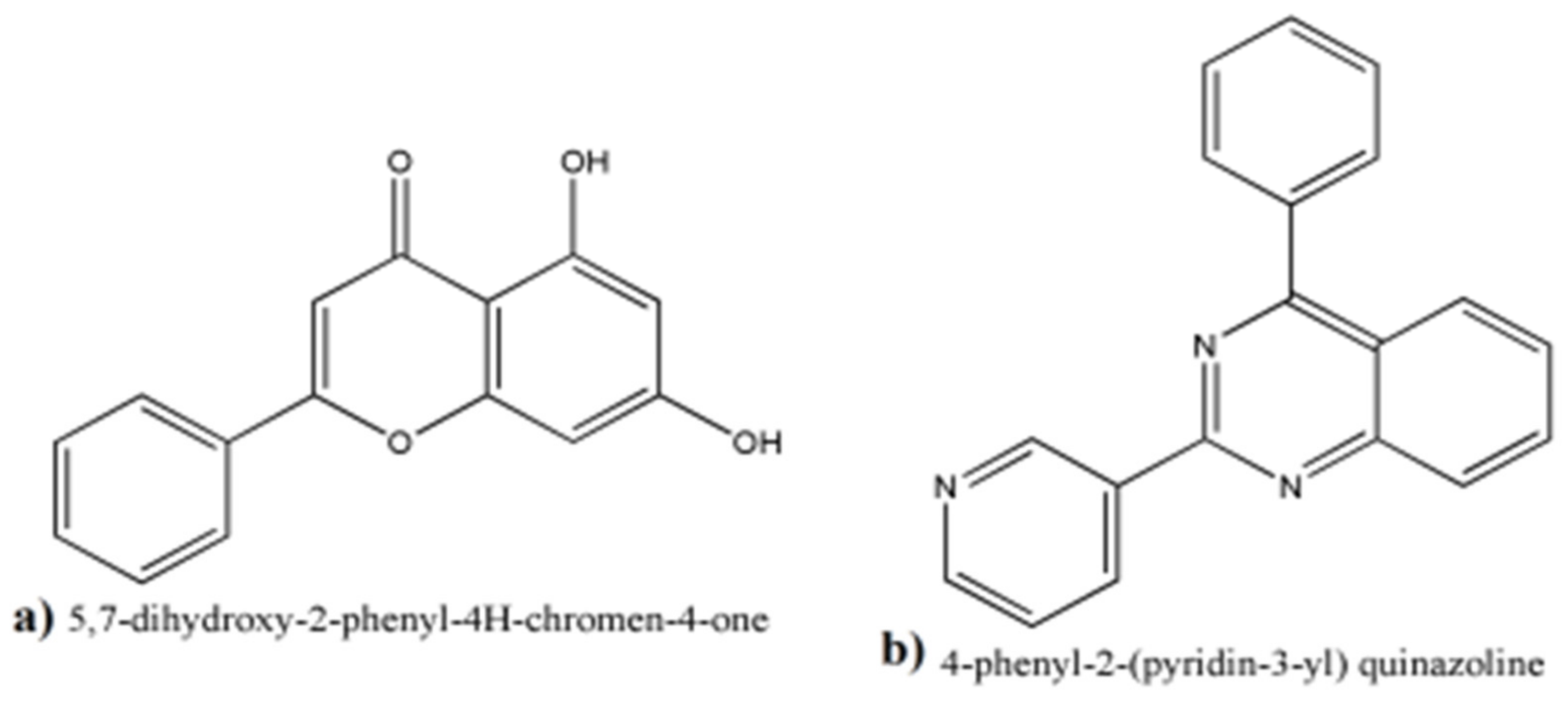
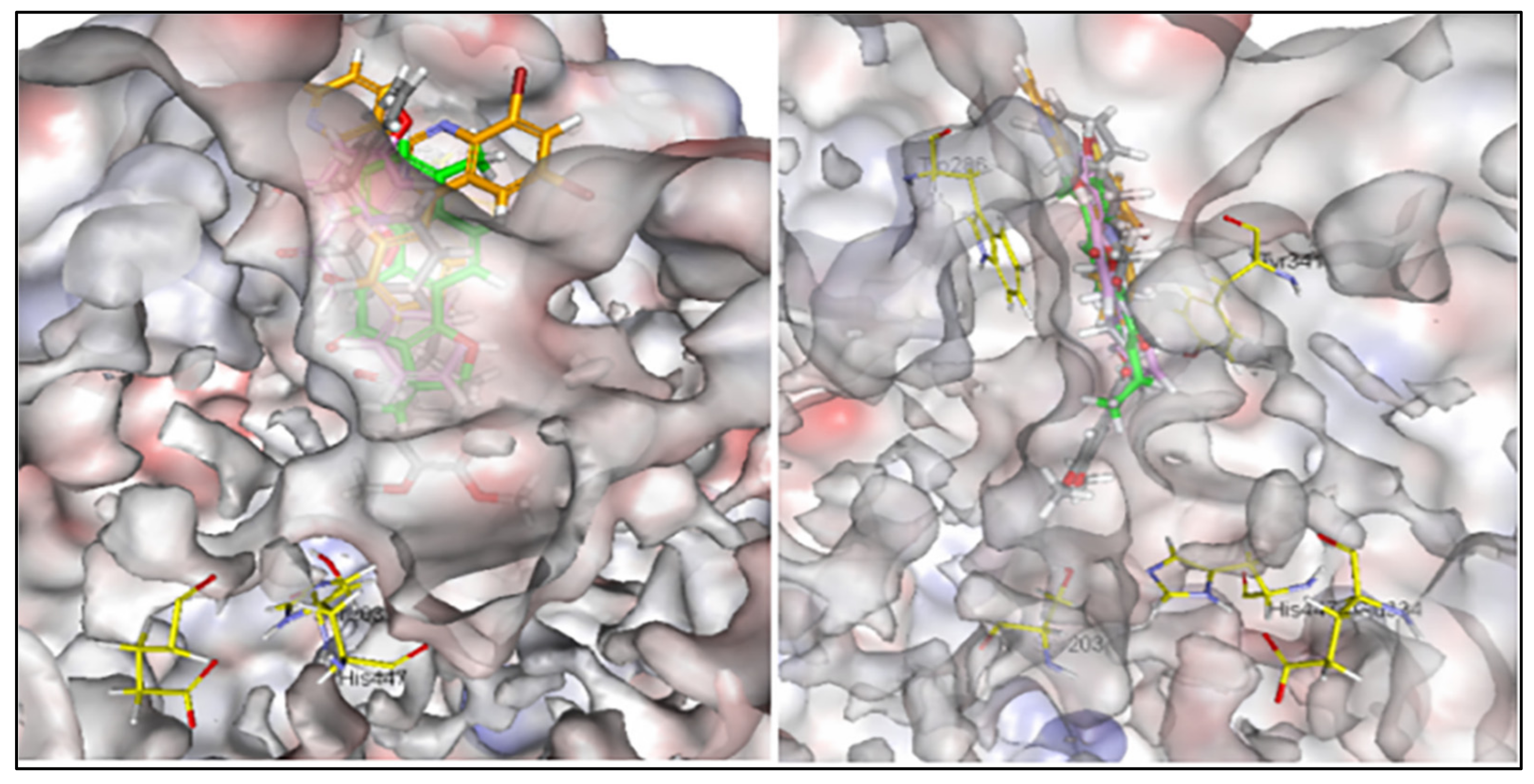
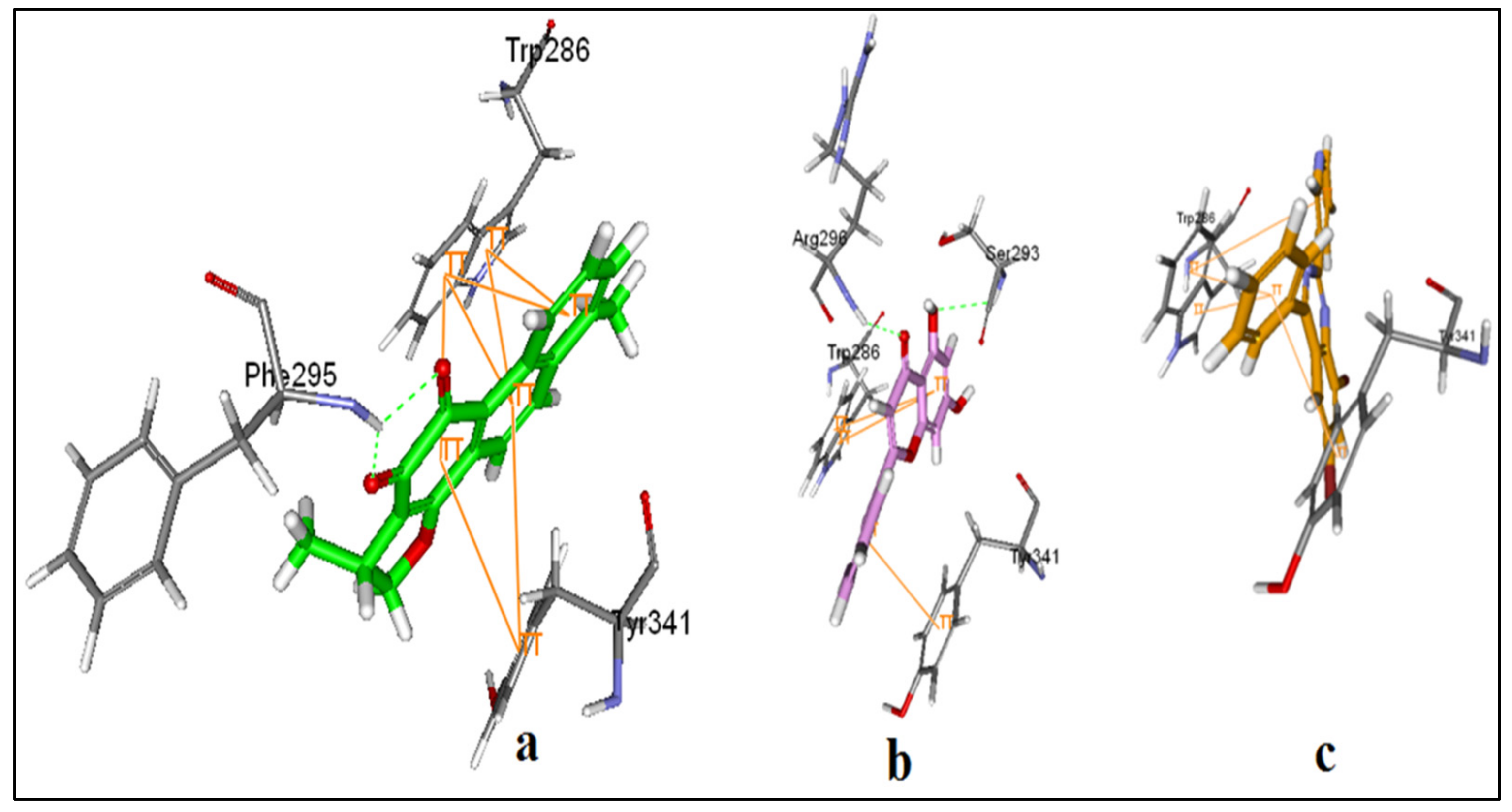
3.8. Alignments and Docking Poses of DHI, Donepezil, 5,7-dihydroxy-2-phenyl-4H-chrome-4-one and 4-phenyl-2-(pyridin-3-yl) Quinazoline
3.9. Binding Energies of the Selected Poses of DHI, Donepezil, 5,7-dihydroxy-2-phenyl-4H-chrome-4-one, and 4-phenyl-2-(pyridin-3-yl) Quinazoline
3.10. Pharmacokinetic Profiles of 5,7-dihydroxy-2-phenyl-4H-chrome-4-one and 4-phenyl-2-(pyridin-3-yl) Quinazoline
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, M.; Zhang, J.; He, S.; Yan, X. A Review study on macrolides isolated from cyanobacteria. Mar. Drugs 2017, 15, 126. [Google Scholar] [CrossRef] [Green Version]
- Soo, R.M.; Hemp, J.; Hugenholtz, P. Evolution of photosynthesis and aerobic respiration in the cyanobacteria. Free Radic. Biol. Med. 2019, 140, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Nandagopal, P.; Steven, A.N.; Chan, L.-W.; Rahmat, Z.; Jamaluddin, H.; Mohd Noh, N.I. Bioactive Metabolites Produced by Cyanobacteria for Growth Adaptation and Their Pharmacological Properties. Biology 2021, 10, 1061. [Google Scholar] [CrossRef] [PubMed]
- Patocka, J.; Gupta, R.C.; Kuca, K. Anatoxin-a (s): Natural organophosphorus anticholinesterase agent. Mil. Med. Sci Lett. 2011, 80, 129–139. [Google Scholar] [CrossRef]
- Becher, P.G.; Beuchat, J.; Gademann, K.; Juttner, F. Nostocarboline: Isolation and synthesis of a new cholinesterase inhibitor from Nostoc 78-12A. J. Nat. Prod. 2005, 68, 1793–1795. [Google Scholar] [CrossRef] [PubMed]
- De la Torre, J.; Aliev, G.; Perry, G. Drug therapy in Alzheimer’s disease. N. Engl. J. Med. 2004, 351, 1911–1913. [Google Scholar] [PubMed]
- Lane, C.A.; Hardy, J.; Schott, J.M. M. Alzheimer’s disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef]
- Reiss, A.B.; Arain, H.A.; Stecker, M.M.; Siegart, N.M.; Kasselman, L.J. Amyloid toxicity in Alzheimer’s disease. Rev. Neurosci. 2018, 29, 613–627. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Ripoll, D.R.; Faerman, C.H.; Axelsen, P.H.; Silman, I.; Sussman, J.L. An electrostatic mechanism for substrate guidance down the aromatic gorge of acetylcholinesterase. Proc. Natl. Acad. Sci. USA 1993, 90, 5128–5132. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the Cholinergic System. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogura, H.; Kosasa, T.; Kuriya, Y.; Yamanishi, Y. Comparison of inhibitory activities of donepezil and other cholinesterase inhibitors on acetylcholinesterase and butylcholinesterase in vitro. Methods Find. Exp. Clin. Pharm. 2000, 22, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, H.; Ogura, H.; Arai, Y.; Limura, Y.; Yamanishi, Y. Research and development of donepezil hydrochloride, a new type of acetylcholinesterase inhibitor. Jpn. J. Pharmacol. 2002, 89, 7–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mimica, N.; Presečki, P. Side effects of approved antidementives. Psychiatr. Danub. 2009, 21, 108–113. [Google Scholar]
- Bajaj, L.B.; Lele, S.S.; Singhal, R.S. A statistical approach to optimization of fermentative production of poly (γ-glutamic acid) from Bacillus licheniformis NCIM2324. Bioresour. Technol. 2009, 100, 826–832. [Google Scholar] [CrossRef]
- Tarko, T.; Duda-Chodak, A.; Kobus, M. Influence of growth medium composition on synthesis of bioactive compounds and antioxidant properties of selected strains of Arthrospira cyanobacteria. Czech J. Food Sci. 2012, 30, 258–267. [Google Scholar] [CrossRef]
- Plackett, R.L.; Burman, J.P. The design of optimum multi factorial experiments. Biometrica 1946, 33, 305–325. [Google Scholar] [CrossRef]
- Anagnostidis, K.; Komárek, J. Modern approach to the classification system of cyanophytes. 3-Oscillatoriales. Algol. Stud./Arch. Hydrobiol. Suppl. 1988, 50–53, 327–472. [Google Scholar]
- Komárek, J.; Anagnostidis, K. Modern approach to the classification system of Cyanophytes 4-Nostocales. Algol. Stud./Arch. Hydrobiol. Suppl. 1989, 82, 247–345. [Google Scholar]
- Stanier, R.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171. [Google Scholar] [CrossRef]
- Araújo, W.L.; Attili de Angellis, D.; Azevedo, J.L. Direct RAPD evaluation of bacteria without conventional DNA extraction. Braz. Arch. Biol. Technol. 2004, 47, 375–380. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.M. Simple conditions for growth of unicellular blue-green algae on plates1, 2. J. Phycol. 1968, 4, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, I.M.; El-Refaay, A.D.; Abdel-Mogib, M.; Azab, A.Y. Studies on biomass and lipid production of seven diatom species withspecial emphasis on lipid composition of Nitzschia palea (Bacillariophyceae) asreliable biodiesel feedstock. Algol. Stud. 2013, 143, 65–68. [Google Scholar] [CrossRef]
- Stein, J.R. Handbook of Phycological Methods, Culture Methods and Growth Measurements; Cambridge University Press: Cambridge, UK, 1973; 417p. [Google Scholar]
- Sadasivam, S.; Manickam, A. Biochemical Methods; New Age International (P) Limited: New Delhi, India, 1996; Volume 2, pp. 124–126. [Google Scholar]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharm. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Komersova, A.; Komers, K.; Čegan, A. New findings about Ellman’s method to determine cholinesterase activity. Z. Nat. C 2007, 62, 150–154. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.-J.; Lee, C.G. Statistical optimization of culture media for production of phycobiliprotein by Synechocystis sp. PCC 6701. Biotechnol. Bioprocess Eng. 2008, 13, 491–498. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Culturing microalgae in outdoor ponds. In Algal Culturing Techniques; Anderson, R.A., Ed.; Elsevier Academic Press: Urlington, VT, USA, 2005; pp. 205–219. [Google Scholar]
- Johnson, T.J.; Jahandideh, A.; Isaac, I.C.; Baldwin, E.L.; Muthukumarappan, K.; Zhou, R.; Gibbons, W.R. Determining the optimal nitrogen source for large-scale cultivation of filamentous cyanobacteria. J. Appl. Phycol. 2017, 29, 1–13. [Google Scholar] [CrossRef]
- Sugimoto, H. The new approach in development of anti-Alzheimer’s disease drugs via the cholinergic hypothesis. Chem. Biol. Interact. 2008, 175, 204–208. [Google Scholar] [CrossRef]
- Zelík, P.; Lukešová, A.; Voloshko, L.N.; Štys, D.; Kopecký, J. Screening for acetylcholinesterase inhibitory activity in cyanobacteria of the genus Nostoc. J. Enzym. Inhib. Med. Chem. 2009, 24, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, L.R.; Costa-Neves, A.; Conserva, G.A.A.; Brunetti, R.L.; Hentschke, G.S.; Malone, C.F.S.; Torres, L.M.B.; Sant’Anna, C.L.; Rangel, M. Biologically active compounds from cyano bacteria extracts: In vivo and in vitro aspects. Braz. J. Pharm. 2013, 23, 471–480. [Google Scholar] [CrossRef] [Green Version]
- Mehta, M.; Adem, A.; Sabbagh, M. New acetylcholinesterase inhibitors for Alzheimer’s disease. Int. J. Alzheimer’s Dis. 2012, 2012, 728983. [Google Scholar] [CrossRef]
- Kong, W.-B.; Hua, S.-F.; Cao, H.; Mu, Y.-W.; Yang, H.; Song, H.; Xia, C.-G. Optimization of the mixotrophic culture medium composition for biomass production by Chlorella vulgaris using response surface methdology. China Biotechnol. 2012, 32, 70–75. [Google Scholar]
- Rania, M.; Abedin, A.; Taha, H.M. Antibacterial and antifungal activity of cyanobacteria and green microlagae. Evaluation of medium components by Plackket-Burman design for antimicrobial activity of Spirulina platensis. J. Biotechnol. Biochem. 2008, 3, 22–31. [Google Scholar]
- Browning, T.J.; Achterberg, E.P.; Jawchuen, Y.; Rapp, I.; Utermann, C.; Engel, A. Iron limitation of microbial phosphorus acquisition in the tropical North Atlantic. Nat. Commun. 2017, 8, 15465. [Google Scholar] [CrossRef] [Green Version]
- Knapp, A.N. The sensitivity of marine N2 fixation to dissolved inorganic nitrogen. Front. Microbiol. 2012, 3, 374. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Juárez, V.; Bennasar-Figueras, A.; Tovar-Sanchez, A.; Agawin, N.S.R. The role of iron in the P-acquisition mechanisms of the unicellular N2-fixing Cyanobacteria halothece sp., found in association with the Mediterranean seagrass Posidonia oceanica. Front. Microbiol. 2019, 10, 1903. [Google Scholar] [CrossRef] [Green Version]
- Boyd, P.W.; Abraham, E.R. Iron-mediated changes in phytoplankton photosynthetic competence during SOIREE. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 48, 2529–2550. [Google Scholar] [CrossRef]
- Sunda, W.G.; Huntsman, S.A. High iron requirement for growth, photosynthesis, and low-light acclimation in the coastal cyanobacterium Synechococcus bacillaris. Front. Microbiol. 2015, 6, 561. [Google Scholar] [CrossRef] [Green Version]
- Deshmukh, D.V.; Puranik, P.R. Application of Plackett-Burman design to evaluate media components affecting antibacterial activity of alkaliphilic cyanobacteria isolated from Lonar lake. Turk. J. Biochem. 2010, 35, 114–120. [Google Scholar]
- Domınguez-Bocanegra, A.; Legarreta, I.G.; Jeronimo, F.M.; Campocosio, A.T. Influence of environmental and nutritional factors in the production of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 2004, 92, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Mayo, D.W.; Pike, R.M.; Forbes, D.C. Microscale Organic Laboratory: With Multistep and Multiscale Syntheses, 5th ed.; J. Wiley & Sons: Hoboken, NJ, USA, 2011; 681p. [Google Scholar]
- Armarego, W.L.F.; Chai, C.L.L. Purification of Laboratory Chemicals, 5th ed.; Butterworth-Heinemann: Amsterdam, The Netherlands, 2003; 609p. [Google Scholar]
- Priya, S. Identification of acetylcholine esterase inhibitors from Morus alba L. leaves. J. Nat. Prod. Plant Resour. 2012, 2, 440–444. [Google Scholar]
- Mukherjee, P.K.; Kumar, V.; Mal, M.; Houghton, P.J. Acetylcholinesterase inhibitors from plants. Phytomedicine 2007, 14, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Cheung, J.; Gary, E.N.; Shiomi, K.; Rosenberry, T.L. Structures of human acetylcholinesterase bound to dihydrotanshinone I and territrem B show peripheral site flexibility. ACS Med. Chem. Lett. 2013, 4, 1091–1096. [Google Scholar] [CrossRef] [Green Version]
- Cheng, T.; Li, X.; Li, Y.; Liu, Z.; Wang, R. Comparative assessment of scoring functions on a diverse test set. J. Chem. Inf. Model. 2009, 49, 1079–1093. [Google Scholar] [CrossRef]
- Podlogar, B.L.; Muegge, I.; Brice, L.J. Computational methods to estimate drug development parameters. Curr. Opin. Drug Disc. Dev. 2001, 4, 102–109. [Google Scholar]
- Carlson, T.J.; Segall, M.D. Predictive, computational models of ADME properties. Curr. Drug Disc. 2002, 7, 34–36. [Google Scholar]
- Egan, W.J.; Merz, K.M.; Baldwin, J.J. Prediction of drug absorption using multivariate statistics. J. Med. Chem. 2000, 43, 3867–3877. [Google Scholar] [CrossRef]
- Bodor, N.; Buchwald, P. Chapter 2.2: Pharmacokinetic Phase: ADME. In Retrometabolic Drug Design and Targeting, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; ISBN 978-1-118-40776-9. [Google Scholar]
- Edwards, M.P.; Price, D.A. Role of Physicochemical Properties and Ligand Lipophilicity Efficiency in Addressing Drug Safety Risks. Annu. Rep. Med. Chem. 2010, 45, 381–391. [Google Scholar]
- Shargel, L.; Susanna, W.P.; Yu, A.B. Chapter 10: Physiological Drug Distribution and Protein Binding. In Applied Biopharmaceutics & Pharmacokinetics, 6th ed.; McGraw-Hill Medical: New York, NY, USA, 2012; p. 211. [Google Scholar]
- Clark, D.E.; Pickett, S. Computational methods for the prediction of ‘drug-likeness’. Drug Discov. Today 2000, 5, 49–58. [Google Scholar] [CrossRef]
- Kerns, E. High throughput physicochemical profiling for drug discovery. J. Pharm. Sci. 2001, 90, 1838–1858. [Google Scholar] [CrossRef]
- Saiakhov, R.D.; Stefan, L.R.; Klopman, G. Multiple computer-automated structure evaluation model of the plasma protein binding affinity of diverse drugs. Perspec. Drug Disc. Des. 2000, 19, 133–155. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Factors | Variables | Zero Level (0) | Low Level (−) | High Level (+) |
|---|---|---|---|---|---|
| 1 | X1 | Ca(NO3)2·4H2O | 0.1 | 0.05 | 0.15 |
| 2 | X2 | K2HPO4·3H2O | 0.14 | 0.07 | 0.21 |
| 3 | X3 | MgSO4·7H2O | 0.025 | 0.0125 | 0.0375 |
| 4 | X4 | Na2SiO3·9H2O | 0.1 | 0.05 | 0.15 |
| 5 | X5 | Na2CO3 | 0.02 | 0.01 | 0.03 |
| 6 | X6 | H3BO3 | 0.0028 | 0.0014 | 0.0042 |
| 7 | X7 | MnCl2·4H2O | 0.0009 | 0.00045 | 0.00135 |
| 8 | X8 | CuSO4·5H2O | 0.0008 | 0.0004 | 0.0012 |
| 9 | X9 | HMoO4 | 0.0009 | 0.00045 | 0.00135 |
| 10 | X10 | ZnCl2 | 0.00013 | 0.00006 | 0.00019 |
| 11 | X11 | CoCl2·6H2O | 0.00004 | 0.00002 | 0.00006 |
| 12 | X12 | FeCl3·6H2O | 0.005 | 0.0025 | 0.0075 |
| 13 | X13 | Na·EDTA·2H2O | 0.03 | 0.015 | 0.045 |
| 14 | X14 | Dummy | - |
| Cyanobacteria Isolates | % Inhibition | Effect |
|---|---|---|
| Anabaena anomala (1) | 25 ± 0.87 | Inhibitory |
| Anabaena khanne | -- * | Stimulatory |
| Anabaena anomala (2) | -- * | Stimulatory |
| Oscillatoria cortiana | -- * | Stimulatory |
| Nostoc entophytum | -- * | Stimulatory |
| Anabaena variabilis | 62 ± 1.3 | Inhibitory |
| Anabaena fertilisma | -- * | Stimulatory |
| Anabaen oryzae | 49 ± 1.5 | Inhibitory |
| Anabaena anomala (3) | -- * | Stimulatory |
| Anabaena varibilis var. ellipsospora | -- * | Stimulatory |
| Donepezil | 100 ± 0.0 |
| Variable | Effect | Coefficient | T-Value | p-Value | Significance |
|---|---|---|---|---|---|
| X1 (Ca(NO3)2·4H2O) | −0.4305 | −0.215 | −1.45 | 0.28 | NS |
| X2 (K2HPO4·3H2O) | −0.0251 | −0.0126 | −0.07 | 0.94 | NS |
| X3 (MgSO4·7H2O) | 0.126 | 0.063 | 0.35 | 0.742 | NS |
| X4 (Na2SiO3·9H2O) | 0.155 | 0.077 | 0.43 | 0.68 | NS |
| X5 (Na2CO3) | 0.236 | 0.118 | 0.65 | 0.54 | NS |
| X6 (H3BO3) | 0.122 | 0.061 | 0.34 | 0.74 | NS |
| X7 (MnCl2·4H2O) | 0.472 | 0.236 | 1.59 | 0.135 | NS |
| X8 (CuSO4·5H2O) | 0.831 | 0.415 | 2.8 | 0.014 | S |
| X9 (HMoO4) | 0.337 | 0.168 | 0.93 | 0.39 | NS |
| X10 (ZnCl2) | −0.568 | −0.284 | −1.91 | 0.076 | S |
| X11 (CoCl2·6H2O) | 0.2449 | 0.122 | 0.68 | 0.52 | NS |
| X12 (FeCl3·6H2O) | 0.7227 | 0.361 | 2.43 | 0.029 | S |
| X13 (Na·EDTA·2H2O) | −0.383 | −0.194 | −1.06 | 0.33 | NS |
| Fractions | Components | % Inhibition |
|---|---|---|
| F1 | Saturated fatty acids | - |
| F2 | Saturated fatty acids | - |
| F3 | Saturated fatty acids | - |
| F4 | Saturated fatty acids | - |
| F5 | Saturated fatty acids | - |
| F6 | Saturated fatty acids | 32.8 |
| F7 | Aromatic compounds, saturated and unsaturated fatty acids | 73.6 |
| F8 | Aromatic compounds, saturated and unsaturated fatty acids | 50 |
| F9 | Fatty materials | 8 |
| F10 | Fatty materials | 45.2 |
| Donepezeil | - | 100 ± 0.0 |
| Compound | CDOCKER Interaction Energy (Kcal mole−1) | CDOCKER Energy (Kcal mole−1) | Scoring Function | ||||
|---|---|---|---|---|---|---|---|
| LigScore1 | LigScore2 | PLP1 | PLP2 | Jain | |||
| Donepezil | 45.18 | 7.24 | 2.68 | 5.52 | 86.14 | 84.95 | 3.81 |
| DHI | 38.92 | 1.30 | 3.28 | 5.54 | 99.96 | 91.50 | 4.27 |
| A | 35.28 | 30.23 | 4.17 | 5.57 | 79.80 | 85.20 | 2.52 |
| B | 32.65 | 5.64 | 1.76 | 5.20 | 76.97 | 68.99 | 0.37 |
| ADMET Tests | ADMET Level | Donepezil | A | B |
|---|---|---|---|---|
| A log P 98 | <5 (good) | 4.12 | 2.652 | 5.301 |
| PSA | <140 Å (good) | 38.77 | 67.861 | 33.783 |
| Absorption | 0 (good) | 0 | 0 | |
| BBB | 0.9 (very good) 2 (medium) | 0.9953 | 0.408 | 0.95 |
| Solubility | 1 (poor) 3 (good) | −3.259 | −6.895 | |
| Hepatotoxicity | 1 (toxic) | 0.933 | 0.933 | |
| CYP2D6 | 1 (inhibitor) | 0.8684 | 0.772 | 0.801 |
| PPB level | 2 (binding ≥ 95%) | 96% | 2 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Refaay, D.A.; Abdel-Hamid, M.I.; Alyamani, A.A.; Abdel Mougib, M.; Ahmed, D.M.; Negm, A.; Mowafy, A.M.; Ibrahim, A.A.; Mahmoud, R.M. Growth Optimization and Secondary Metabolites Evaluation of Anabaena variabilis for Acetylcholinesterase Inhibition Activity. Plants 2022, 11, 735. https://doi.org/10.3390/plants11060735
Refaay DA, Abdel-Hamid MI, Alyamani AA, Abdel Mougib M, Ahmed DM, Negm A, Mowafy AM, Ibrahim AA, Mahmoud RM. Growth Optimization and Secondary Metabolites Evaluation of Anabaena variabilis for Acetylcholinesterase Inhibition Activity. Plants. 2022; 11(6):735. https://doi.org/10.3390/plants11060735
Chicago/Turabian StyleRefaay, Dina A., Mohammed I. Abdel-Hamid, Amal A. Alyamani, Mamdouh Abdel Mougib, Dalia M. Ahmed, Amr Negm, Amr M. Mowafy, Amira A. Ibrahim, and Rania M. Mahmoud. 2022. "Growth Optimization and Secondary Metabolites Evaluation of Anabaena variabilis for Acetylcholinesterase Inhibition Activity" Plants 11, no. 6: 735. https://doi.org/10.3390/plants11060735
APA StyleRefaay, D. A., Abdel-Hamid, M. I., Alyamani, A. A., Abdel Mougib, M., Ahmed, D. M., Negm, A., Mowafy, A. M., Ibrahim, A. A., & Mahmoud, R. M. (2022). Growth Optimization and Secondary Metabolites Evaluation of Anabaena variabilis for Acetylcholinesterase Inhibition Activity. Plants, 11(6), 735. https://doi.org/10.3390/plants11060735