
Drying Grapes after the Application of Different Dipping Solutions: Effects on Hormones, Minerals, Vitamins, and Antioxidant Enzymes in Gök Üzüm (Vitis vinifera L.) Raisins





Abstract
1. Introduction
2. Results
2.1. Hormone Content in the Raisins
2.2. Antioxydant Enzyme Activity in the Raisins
2.3. Vitamins Content in the Raisins
2.4. Mineral Content in the Raisins
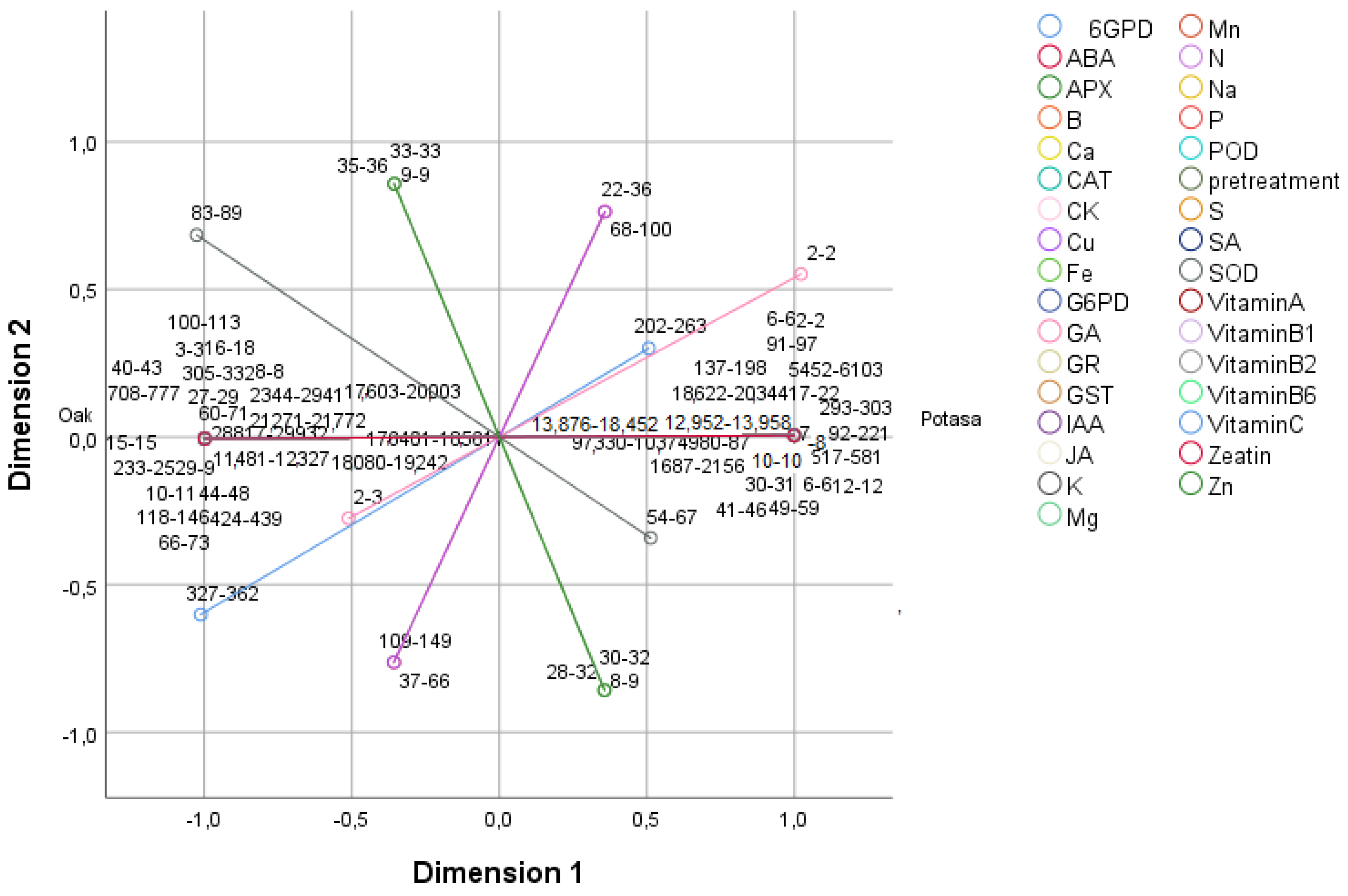
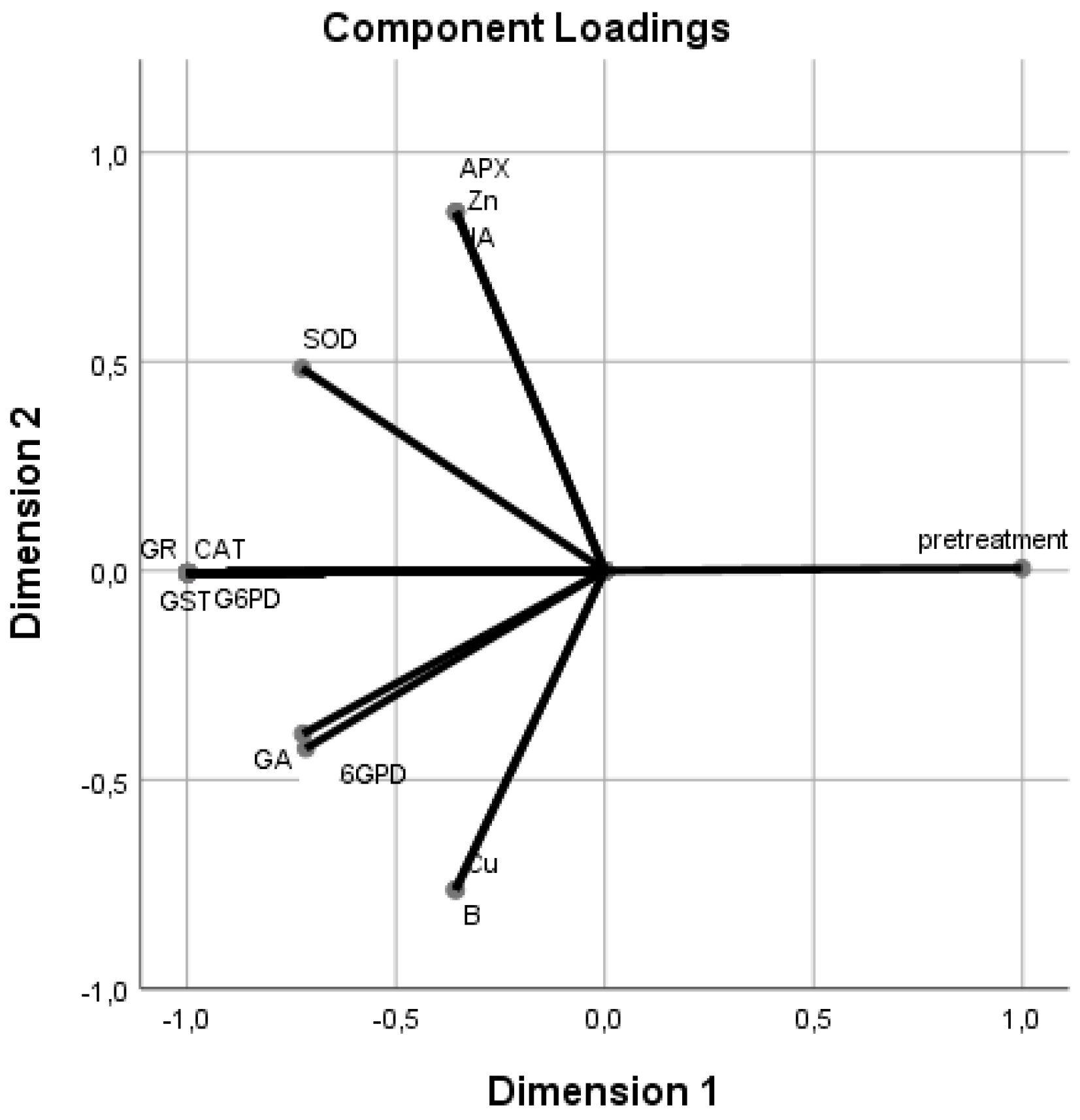
2.5. Principal Component Analysis
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Treatments
5.2. Hormone Analysis in Raisins
5.3. Antioxidant Enzyme Analysis in Raisins
5.4. Vitamin Analysis in Raisins
5.5. Mineral Element Analysis in Raisins
5.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Agriculture Data. Available online: http://apps.fao.org/page/collections?subset=agriculture (accessed on 10 January 2022).
- Uysal, H.; Karabat, S. Forecasting and evaluation for raisin export in turkey. BIO Web Conf. 2017, 9, 03002. [Google Scholar] [CrossRef]
- Carughi, A. Health Benefits of Sun-Dried Raisins; Health Research and Studies Center: Kingsburg, CA, USA, 2009; pp. 2–63. [Google Scholar]
- Sério, S.; Rivero-Pérez, M.D.; Correia, A.C.; Jordão, A.M.; González-San José, M.L. Analysis of commercial grape raisins: Phenolic content, antioxidant capacity and radical scavenger activity. Ciênc. Tec. Vitivin. 2014, 29, 1–8. [Google Scholar] [CrossRef]
- Franco, M.; Peinado, R.A.; Medina, M.; Moreno, J. Off-vine grape drying effect on volatile compounds and aromatic series in must from Pedro Ximénez grape variety. J. Agric. Food Chem. 2004, 52, 3905–3910. [Google Scholar] [CrossRef] [PubMed]
- Panagopoulou, E.A.; Chiou, A.; Nikolidaki, E.K.; Christea, M.; Karathanos, V.T. Corinthian raisins (Vitis vinifera L., var. Apyrena) antioxidant and sugar content as affected by the drying process: A 3-year study. J. Sci. Food Agric. 2019, 99, 915–922. [Google Scholar] [CrossRef] [PubMed]
- EL-Mesery, H.S.; EL-Seesy, A.I.; Hu, Z.; Li, Y. Recent developments in solar drying technology of food and agricultural products: A review. Renew. Sustain. Energ. Rev. 2022, 157, 112070. [Google Scholar] [CrossRef]
- Panagopoulou, E.A.; Chiou, A.; Karathanos, V.T. Water-soluble vitamin content of sun-dried Corinthian raisins (Vitis vinifera L., var. Apyrena). J. Sci. Food Agric. 2019, 99, 5327–5333. [Google Scholar] [CrossRef]
- Akın, A. Effects of cluster reduction, herbagreen and humic acid applications on grape yield and quality of Horoz Karasi and Gök üzüm grape cultivars. Afr. J. Biotechnol. 2011, 10, 5593–5600. [Google Scholar]
- Esmaiili, M.; Sotudeh-Gharebagh, R.; Cronin, K.; Mousavi, M.A.E.; Rezazadeh, G. Grape drying: A review. Food Rev. Int. 2007, 23, 257–280. [Google Scholar] [CrossRef]
- Pastori, G.M.; Foyer, C.H. Common components networks and pathways of cross-tolerance to stress the central role of “redox” and abscisic acid-mediated. Plant Physiol. 2002, 129, 460–468. [Google Scholar] [CrossRef]
- Raskin, I. Role of salicylic acid in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 439–463. [Google Scholar] [CrossRef]
- Murphy, A.M.; Chivasa, S.; Singh, D.P.; Carr, J.P. Salicylic acid-induced resistance to viruses and other pathogens: A parting of the ways? Trends Plant Sci. 1999, 4, 155–160. [Google Scholar] [CrossRef]
- Rüffer, M.; Steipe, B.; Zenk, M.H. Evidence against specific binding of salicylic acid to plant catalase. FEBS Lett. 1995, 377, 175–180. [Google Scholar] [CrossRef]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Van Montagu, M.; Inzé, D.; Van Camp, W. Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J. 1997, 16, 4806–4816. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yuan, B.; Leng, P. The role of ABA in triggering ethylene biosynthesis and ripening of tomato fruit. J. Exp. Bot. 2009, 60, 1579–1588. [Google Scholar] [CrossRef]
- Martínez-Romero, D.; Valero, D.; Serrano, M.; Burló, F.; Carbonell, A.; Burgos, L.; Riquelme, F. Exogenous polyamines and gibberellic acid effects on peach (Prunus persica L.) storability improvement. J. Food Sci. 2000, 65, 288–294. [Google Scholar] [CrossRef]
- Singh, R.; Singh, P.; Pathak, N.; Singh, V.K.; Dwivedi, U.N. Modulation of mango ripening by chemicals: Physiological and biochemical aspects. Plant Growth Regul. 2007, 53, 137–145. [Google Scholar] [CrossRef]
- Sudha, R.R.; Amutha, S.; Muthulaksmi, W.; Baby, R.; Indira, K.; Mareeswari, P. Influence of pre and post harvest chemical treatments on physical characteristics of sapota (Achras sapota L.) var. PKM 1. J. Agric. Biol. Sci. 2007, 3, 450–452. [Google Scholar]
- Fan, X.; Mattheis, J.P.; Fellman, J.K. A role for jasmonates in climacteric fruit ripening. Planta 1998, 204, 444–449. [Google Scholar] [CrossRef]
- Bingol, G.; Roberts, J.S.; Balaban, M.O.; Devres, Y.O. Effect of dipping temperature and dipping time on drying rate and color change of grapes. Dry. Technol. 2012, 30, 597–606. [Google Scholar] [CrossRef]
- Ribalta-Pizarro, C.; Muñoz, P.; Munné-Bosch, S. Tissue-specific hormonal variations in grapes of ırrigated and non-irrigated grapevines (Vitis vinifera cv. “Merlot”) growing under mediterranean field conditions. Front. Plant Sci. 2021, 12, 621587. [Google Scholar] [CrossRef]
- Demir, H.; Güler, A.; Demir, C. Determination of some antioxidan enzyme activities in fresh and dry grapes (Vitis vinifera L.). Van Tıp. Derg. 2020, 27, 150–154. [Google Scholar] [CrossRef]
- Breksa III, A.P.; Takeoka, G.R.; Hidalgo, M.B.; Vilches, A.; Vasse, J.; Ramming, D.W. Antioxidant activity and phenolic content of 16 raisin grape (Vitis vinifera L.) cultivars and selections. Food Chem. 2010, 121, 740–745. [Google Scholar] [CrossRef]
- Hogan, S.; Zhang, L.; Li, J.; Zoecklein, B.; Zhou, K. Antioxidant properties and bioactive components of Norton (Vitis vinifera) wine grapes. LWT 2009, 42, 1269–1274. [Google Scholar] [CrossRef]
- Zhao, D.Y.; Shen, L.; Yu, M.M.; Zheng, Y.; Sheng, J.P. Relationship between activities of antioxidant enzymes and cold tolerance of postharvest tomato fruits. Food Sci. 2009, 14, 309–313. [Google Scholar]
- OIV. Resolutıon OIV-VITI 522-2016. OIV Good Practıses for Drıed Grape Productıon Systems. Bento Gonçalves, Brazil. 2016. Available online: https://www.oiv.int/public/medias/4956/oiv-viti-522-2016-en.pdf (accessed on 28 January 2022).
- Olivati, C.; de Oliveira Nishiyama, Y.P.; de Souza, R.T.; Janzantti, N.S.; Mauro, M.A.; Gomes, E.; Hermosín-Gutiérrezc, I.; da Silva, R.; Lago-Vanzela, E.S. Effect of the pre-treatment and the drying process on the phenolic composition of raisins produced with a seedless Brazilian grape cultivar. Food Res. Int. 2019, 116, 190–199. [Google Scholar] [CrossRef]
- Khiari, R.; Zemni, H.; Mihoubi, D. Raisin processing: Physicochemical, nutritional and microbiological quality characteristics as affected by drying process. Food Rev. Int. 2019, 35, 246–298. [Google Scholar] [CrossRef]
- Rustioni, L.; Maghradze, D.; Failla, O. Optical properties of berry epicuticular waxes in four Georgian grape cultivars (Vitis vinifera L.). S. Afr. J. Enol. Vitic. 2012, 33, 138–143. [Google Scholar] [CrossRef][Green Version]
- Çaglarirmak, N.; Ochratoxin, A. Hydroxymethylfurfural and vitamin c levels of sun-dried grapes and sultanas. J. Food Process. Preserv. 2006, 30, 549–562. [Google Scholar] [CrossRef]
- USDA. Basic Report: 09298, Raisins, Seedless, National Nutrient Database for Standard Reference Release 28, United States Department of Agriculture, Agricultural Service. Available online: https://ndb.nal.usda.gov/ndb/foods/show/2371 (accessed on 15 September 2021).
- Iqbal, M.P.; Kazim, S.F.; Mehboobali, N. Ascorbic acid contents of Pakistani fruits and vegetables. Pak. J. Pharm. Sci. 2006, 19, 282–285. [Google Scholar]
- Nikniaz, Z.; Mahdavi, R.; Rafraf, M.; Jouyban, A. Total phenols and vitamin C contents of Iranian fruits. Nutr. Food Sci. 2009, 6, 603–608. [Google Scholar] [CrossRef]
- Keskin, N.; Bilir Ekbic, H.; Kaya, O.; Keskin, S. Antioxidant activity and biochemical compounds of Vitis vinifera L. (cv.‘Katıkara’) and Vitis labrusca L.(cv.‘Isabella’) grown in Black Sea Coast of Turkey. Erwerbs-Obstbau 2021, 63, 115–122. [Google Scholar] [CrossRef]
- Langová, R.; Jůzl, M.; Cwiková, O.; Kos, I. Effect of different method of drying of five varieties grapes (Vitis vinifera L.) on the bunch stem on physicochemical, microbiological, and sensory quality. Foods 2020, 9, 1183. [Google Scholar] [CrossRef] [PubMed]
- Clary, C.D.; Mejia-Meza, E.; Wang, S.; Petrucci, V.E. ımproving grape quality using microwave vacuum drying associated with temperature control. J. Food Sci. 2007, 72, E023–E028. [Google Scholar] [CrossRef] [PubMed]
- Joshi, V.K. Concise Encyclopedia of Science and Technology of Wine; CRC Press: Boca Raton, FL, USA, 2021; p. 710. [Google Scholar]
- Fabani, M.P.; Baroni, M.V.; Luna, L.; Lingua, M.S.; Monferran, M.V.; Paños, H.; Feresin, G.E. Changes in the phenolic profile of Argentinean fresh grapes during production of sun-dried raisins. J. Food Compos. Anal. 2017, 58, 23–32. [Google Scholar] [CrossRef]
- Ghrairi, F.; Lahouar, L.; Brahmi, F.; Ferchichi, A.; Achour, L.; Said, S. Physicochemical composition of different varieties of raisins (Vitis vinifera L.) from Tunisia. Ind. Crops Prod. 2013, 43, 73–77. [Google Scholar] [CrossRef]
- Zemni, H.; Sghaier, A.; Khiari, R.; Chebil, S.; Ben Ismail, H.; Nefzaoui, R.; Hamdi, S.; Lasram, S. Physicochemical, phytochemical and mycological characteristics of Italia Muscat raisins obtained using different pre-treatments and drying techniques. Food Bioprocess Technol. 2016, 10, 479–490. [Google Scholar] [CrossRef]
- Carranza-Concha, J.; Benlloch, M.; Camacho, M.M.; Martínez-Navarrete, N. Effects of drying and pretreatment on the nutritional and functional quality of raisins. Food Bioprod. Process. 2012, 90, 243–248. [Google Scholar] [CrossRef]
- Battal, P.; Tileklioğlu, B. The effects of different mineral nutrients on the levels of cytokinins in Maize (Zea mays L.). Turk. J. Bot. 2001, 25, 123–130. [Google Scholar]
- Abedi, T.; Pakniyat, H. Antioxidant enzyme changes in response to drought stress in ten cultivars of oilseed rape (Brassica napus L.). Czech J. Genet. Plant Breed. 2010, 46, 27–34. [Google Scholar] [CrossRef]
- Angelini, R.; Manes, F.; Federico, R. Spatial and functional correlation between daimine- oxidase and peroxidase activities and their dependence upon deetilation and wounding in chick-pea. Planta 1990, 182, 89–96. [Google Scholar] [CrossRef]
- Minucci, A.; Giardina, B.; Zuppi, C.; Capoluongo, E. Glucose-6-phosphate dehydrogenase laboratory assay: How, when, and why? IUBMB Life 2009, 61, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Chikezie, P.C.; Chikezie, C.M.; Uwakwe, A.A.; Monako, C.C. Comparative study of glutathione S-transferase activity of three human erythrocyte genotypes infected with plasmodium falciparum. JASEM 2009, 13, 13–18. [Google Scholar] [CrossRef]
- Kumar, S.S.; Samydurai, P.; Ramakrishnan, R.; Nagarajan, N. Polyphenols, vitamin-E estimation and in vitro antioxidant activity of Adiantum capillus-veneris. Int. J. Innov. Pharm. Sci. 2013, 4, 258–262. [Google Scholar]
- Mozumder, N.R.; Akhter, M.J.; Khatun, A.A.; Rokibuzzaman, M.; Akhtaruzzaman, M. Estimation of water-soluble vitamin B-complex in selected leafy and non-leafy vegetables by HPLC method. Orient. J. Chem. 2019, 35, 1344. [Google Scholar] [CrossRef]
- AOAC. Official Method Analysis 975.03. Metals in Plants and Pets Food Atomic Absorption Spectrophotometric Method. 2005. Available online: https://scirp.org/reference/referencespapers.aspx?referenceid=2783175 (accessed on 10 January 2022).



{kind=link}
{kind=link}
| Hormones | Oak Ash | Potassium Carbonate |
|---|---|---|
| Indol-3-acetic acid (IAA) | 8.75 ± 0.28 b | 6.40 ± 0.24 a |
| Abscisic acid (ABA) | 11,868.40 ± 427.65 b | 5751.18 ± 328.91 a |
| Gibberellic acid (GA) | 2.98 ± 0.05 a | 2.87 ± 0.06 a |
| Salicylic acid (SA) | 9.45 ± 0.12 b | 6.55 ± 0.04 a |
| Cytokinin (CK) | 15.28 ± 0.17 b | 10.23 ± 0.11 a |
| Zeatin | 3.36 ± 0.09 b | 2.24 ± 0.14 a |
| Jasmonic acid (JA) | 9.23 ± 0.52 a | 9.34 ± 0.21 a |
| Antioxidant Enzymes | Oak Ash | Potassium Carbonate |
|---|---|---|
| Glutathione reductase (GR) | 27.90 ± 1.05 b | 19.69 ± 2.13 a |
| Glutathione S-transferase (GST) | 744.85 ± 35.05 b | 540.08 ± 35.63 a |
| Glucose-6-phosphate dehydrogenase (G6PD) | 241.87 ± 9.59 b | 208.25 ± 14.62 a |
| 6 Glucose phosphate dehydrogenase (6GPD) | 317.44 ± 50.26 b | 214.50 ± 10.96 a |
| Catalase (CAT) | 41.54 ± 2.02 b | 32.53 ± 2.01 a |
| Peroxidase (POD) | 60.01 ± 5.30 b | 44.56 ± 2.83 a |
| Superoxide dismutase (SOD) | 79.89 ± 11.15 b | 56.23 ± 2.25 a |
| Ascorbate peroxidase (APX) | 34.70 ± 2.09 a | 31.33 ± 4.25 a |
| Vitamins | Oak Ash | Potassium Carbonate |
|---|---|---|
| Vitamin B1 | 70.19 ± 3.58 b | 55.02 ± 5.02 a |
| Vitamin B2 | 135.54 ± 14.88 b | 95.17 ± 3.26 a |
| Vitamin B6 | 107.30 ± 6.51 b | 83.88 ± 0.60 a |
| Vitamin C | 17.28 ± 1.08 b | 12.54 ± 0.30 a |
| Vitamin A | 46.64 ± 1.61 b | 31.24 ± 0.26 a |
| Minerals | Oak Ash | Potassium Carbonate |
|---|---|---|
| Nitrogen | 11.31 ± 0.47 b | 8.02 ± 0.07 a |
| Calcium | 29,466.93 ± 579.74 b | 19,332.86 ± 899.59 a |
| Potassium | 180,020.47 ± 4911.81 b | 101,177.28 ± 3393.71 a |
| Magnesium | 18,628.89 ± 582.34 b | 12,426.03 ± 419.78 a |
| Sodium | 2688.42 ± 308.38 b | 1893.81 ± 239.39 a |
| Phosphorous | 21,540.92 ± 252.62 b | 13,448.70 ± 498.18 a |
| Sulfur | 18,892.61 ± 1210.07 b | 13,896.70 ± 704.97 a |
| Manganese | 320.33 ± 13.63 b | 176.68 ± 33.62 a |
| Copper | 107.15 ± 5.73 a | 96.05 ± 46.29 a |
| Iron | 430.48 ± 8.13 b | 300.32 ± 5.62 a |
| Zinc | 32.68 ± 1.85 a | 33.08 ± 0.76 a |
| Boron | 37.75 ± 1.50 a | 37.86 ± 25.21 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Keskin, N.; Kaya, O.; Ates, F.; Turan, M.; Gutiérrez-Gamboa, G. Drying Grapes after the Application of Different Dipping Solutions: Effects on Hormones, Minerals, Vitamins, and Antioxidant Enzymes in Gök Üzüm (Vitis vinifera L.) Raisins. Plants 2022, 11, 529. https://doi.org/10.3390/plants11040529
Keskin N, Kaya O, Ates F, Turan M, Gutiérrez-Gamboa G. Drying Grapes after the Application of Different Dipping Solutions: Effects on Hormones, Minerals, Vitamins, and Antioxidant Enzymes in Gök Üzüm (Vitis vinifera L.) Raisins. Plants. 2022; 11(4):529. https://doi.org/10.3390/plants11040529
Chicago/Turabian StyleKeskin, Nurhan, Ozkan Kaya, Fadime Ates, Metin Turan, and Gastón Gutiérrez-Gamboa. 2022. "Drying Grapes after the Application of Different Dipping Solutions: Effects on Hormones, Minerals, Vitamins, and Antioxidant Enzymes in Gök Üzüm (Vitis vinifera L.) Raisins" Plants 11, no. 4: 529. https://doi.org/10.3390/plants11040529
APA StyleKeskin, N., Kaya, O., Ates, F., Turan, M., & Gutiérrez-Gamboa, G. (2022). Drying Grapes after the Application of Different Dipping Solutions: Effects on Hormones, Minerals, Vitamins, and Antioxidant Enzymes in Gök Üzüm (Vitis vinifera L.) Raisins. Plants, 11(4), 529. https://doi.org/10.3390/plants11040529