
Impact of Silica Ions and Nano Silica on Growth and Productivity of Pea Plants under Salinity Stress



Abstract
:1. Introduction
2. Results
2.1. Plant Vegetative Growth
2.2. Yield Components
2.3. Ion Selectivity
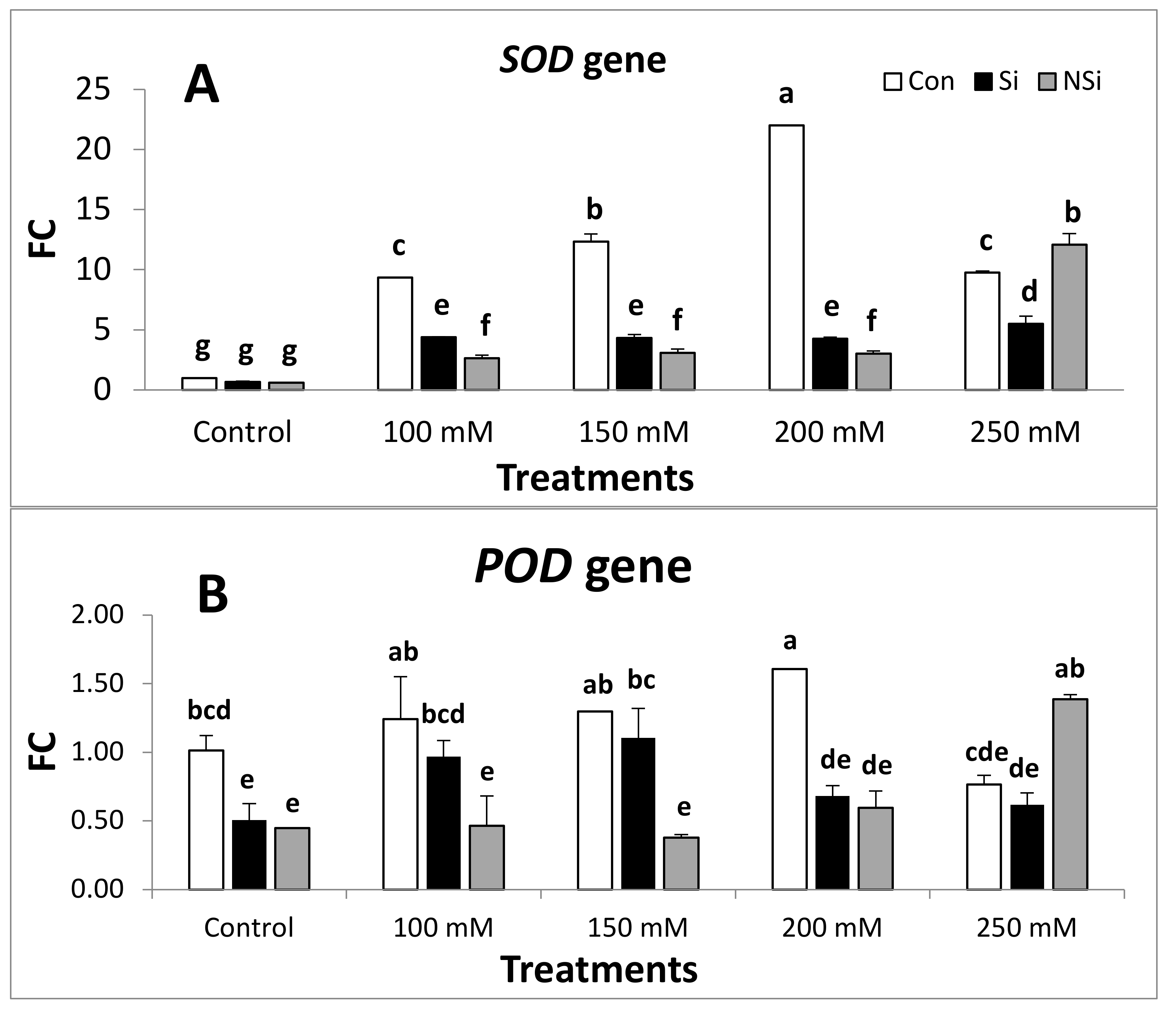
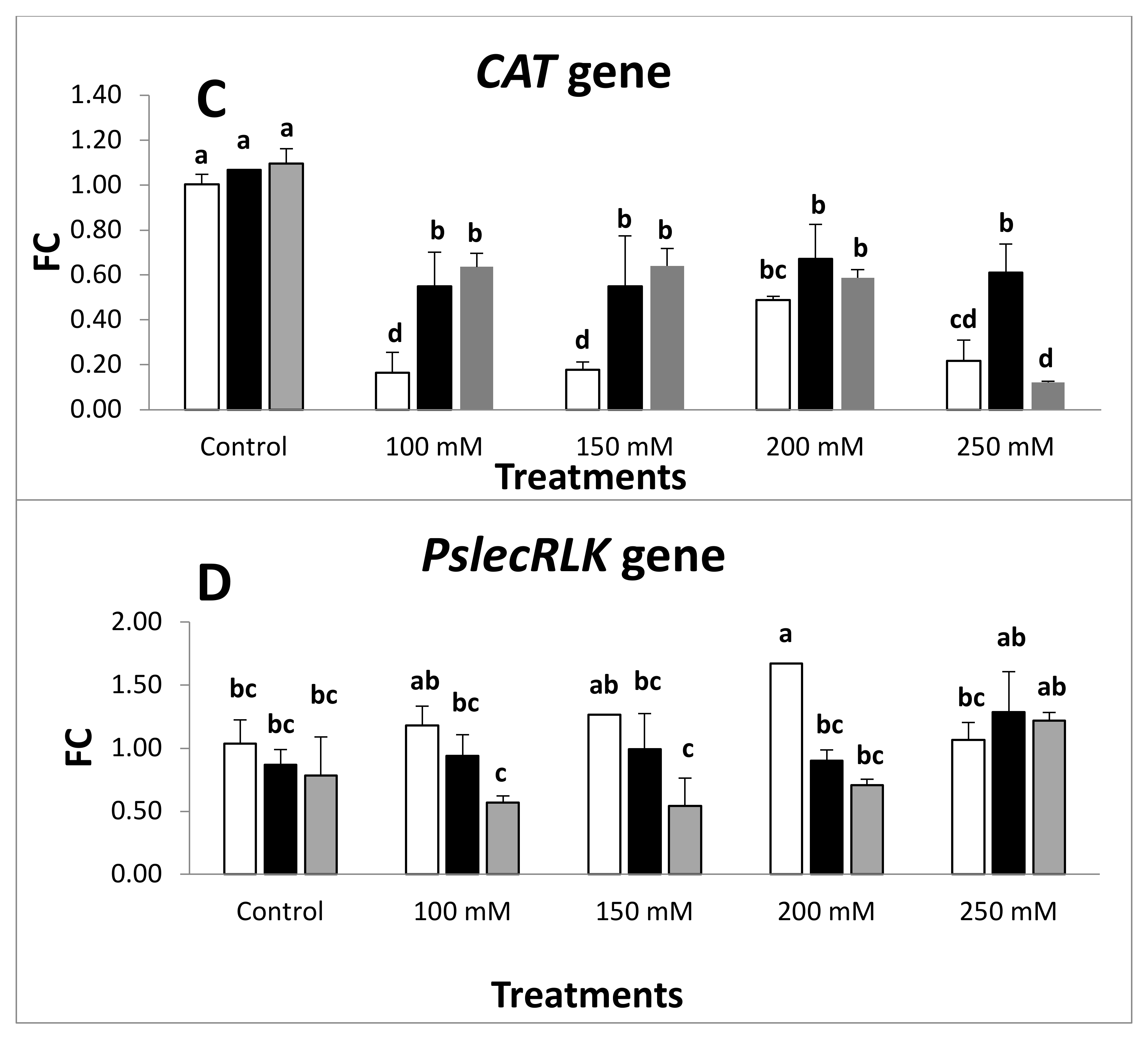
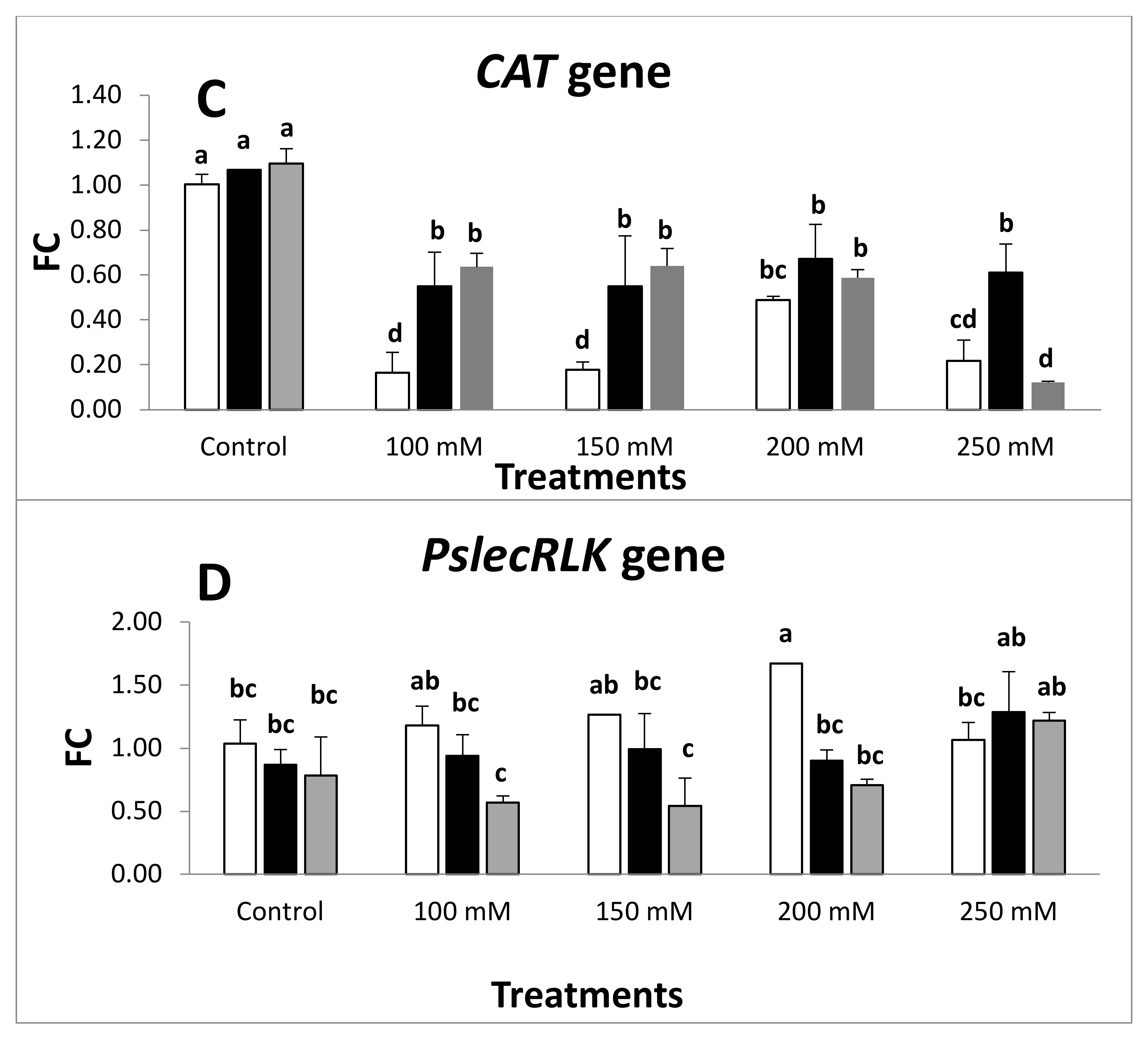
2.4. Expression of Antioxidant Defense Genes
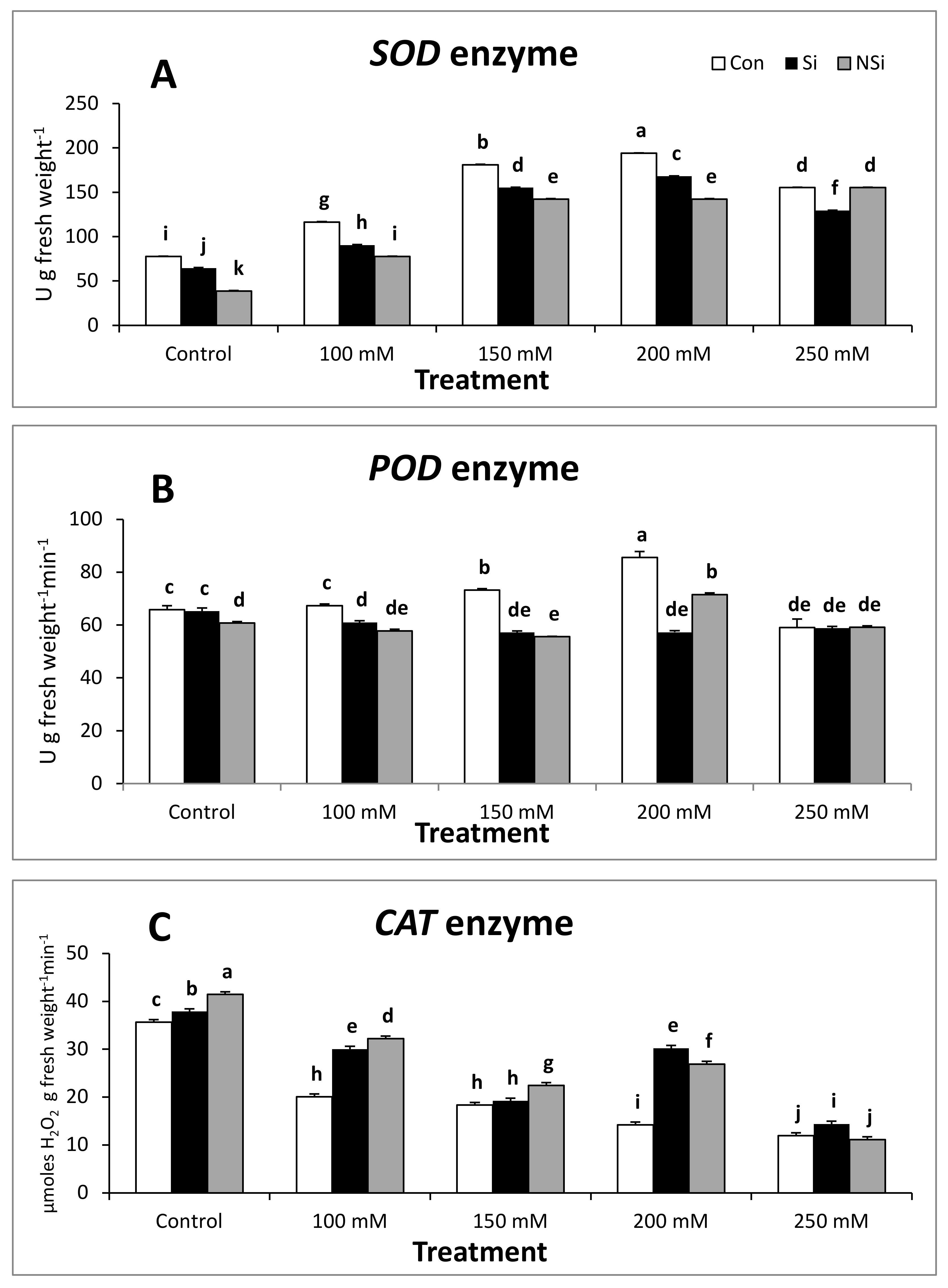
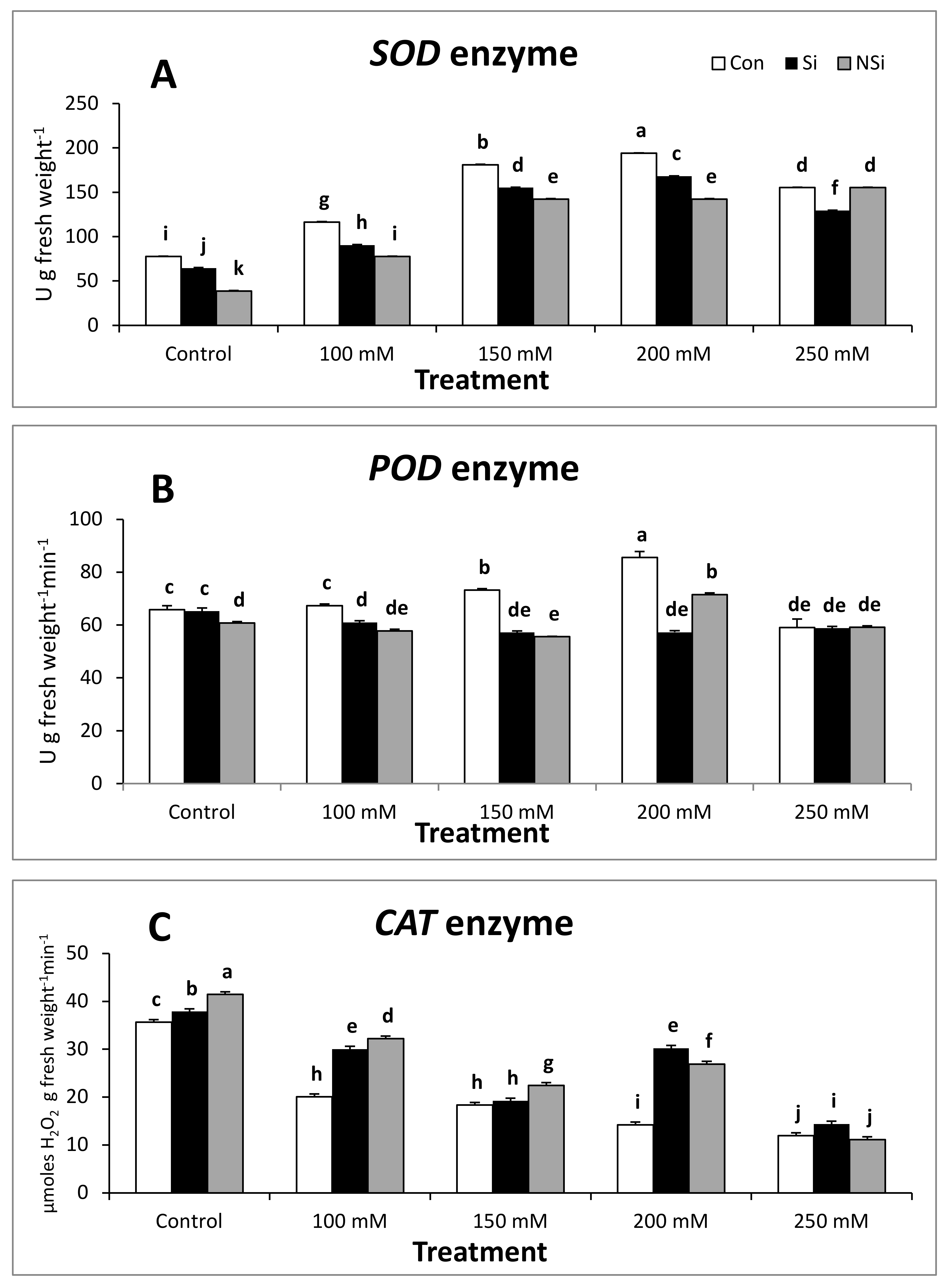
2.5. Antioxidant Enzyme Activities
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Estimation of Growth and Yield Parameters
4.3. Estimation of Ions
4.4. Gene Expression Analysis
4.5. Estimation of Antioxidant Enzyme Activities
4.5.1. Enzyme Extraction
4.5.2. Estimation of Superoxide Dismutase Activity (SOD, EC1.11.1.7)
4.5.3. Estimation of Peroxidase Activity (POD, EC1.11.1.7)
4.5.4. Estimation of Catalase Activity (CAT, EC1.11.1.6)
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Taha, R.S.; Seleiman, M.F.; Alhammad, B.A.; Alkahtani, J.; Alwahibi, M.S.; Mahdi, A.H.A. Activated Yeast Extract Enhances Growth, Anatomical Structure, and Productivity of Lupinus termis L. Plants under Actual Salinity Conditions. Agronomy 2021, 11, 74. [Google Scholar]
- Zheng, L.; Shannon, M.C.; Lesch, S.M. Timing of salinity stress affecting rice growth and yield components. Agric. Water Manag. 2001, 48, 191–206. [Google Scholar] [CrossRef]
- Zayed, B.A.; Okasha, A.M.; Rashwan, E. Impact of different rates of phosphoric acid foliar applicationing on rice growth and yield traits under normal and saline soils conditions. J. Agric. Life Sci. 2019, 2, 56–66. [Google Scholar]
- Dahl, W.J.; Foster, L.M.; Tyler, R.T. Review of the health benefits of peas (Pisum sativum L.). Br. J. Nutr. 2012, 108 (Suppl. 1), S3–S10. [Google Scholar] [CrossRef] [Green Version]
- Tulbek, M.C.; Lam, R.S.H.; Wang, Y.C.; Asavajaru, P.; Lam, A. Pea: A sustainable vegetable protein crop. In Sustainable Protein Sources; Nadathur, S.R., Wanasundara, J.P.D., Scanlin, L., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 145–164. [Google Scholar]
- Kosev, V.; Pachev, I. Genetic improvement of field pea (Pisum sativum L.) in Bulgaria. Ratar. Povrt. Field Veg. Crop Res. 2010, 47, 403–408. [Google Scholar]
- Omokanye, A.T.; Kelleher, F.M.; McInnes, A. Low-Input cropping systems and nitrogen fertilizer effects on crop production: Soil nitrogen dynamics and efficiency of nitrogen use in maize crop. Am.-Eur. J. Agric. Environ. Sci. 2011, 11, 282–295. [Google Scholar]
- Kocer, A.; Albayrak, S. Determination of forage yield and quality of pea (Pisum sativum L.) mixtures with oat and barley. Turk. J. Field Crops 2012, 17, 96–99. [Google Scholar]
- Najafi, F.; Khavari-Nejad, R.A.; Rastgar-jazii, F.; Sticklen, M. Growth and some physiological attributes of pea (Pisum sativum L.) as affected by salinity. Pak. J. Biol. Sci. 2007, 10, 2752–2755. [Google Scholar]
- Agami, R.A. Alleviating the adverse effects of NaCl stress in maize seedlings by pretreating seeds with salicylic acid and 24-epibrassinolide. South Afr. J. Bot. 2013, 88, 171–177. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Jang, K.H.; Farooq, M.; Lee, D.J. Morphological and physiological evaluation of Korean rice genotypes for salt resistance. Int. J. Agric. Biol. 2012, 14, 970–974. [Google Scholar]
- Lüchli, A.; Grattan, S.R. Plant growth and development under salinity stress. In Advances in Molecular Breeding Toward Drought and Salt Tolerant Crops; Jenks, M.A., Hasegawa, P.M., Jain, S.M., Eds.; Springer Nature: Cham, Switzerland, 2007; pp. 1–32. [Google Scholar]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genomics 2014, 2014, 701596. [Google Scholar] [CrossRef]
- Shalata, A.; Neumann, P.M. Exogenous ascorbic acid (vitamin C) increase resistance to salt stress and reduces lipid peroxidation. J. Exp. Bot. 2001, 52, 2207–2211. [Google Scholar] [CrossRef] [PubMed]
- Younis, M.E.; Hasaneen, M.N.A.; Kazamel, A.M.S. Plant growth, metabolism and adaptation in relation to stress conditions. XXVII. Can ascorbic acid modify the adverse effects of NaCl and mannitol on amino acids, nucleic acids and protein patterns in Vicia faba seedlings? Protoplasma 2009, 235, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Hemantaranjan, A.; Patel, P.K. Salicylic acid improves salinity tolerance in field pea (Pisum sativum L.) by intensifying antioxidant defense system and preventing salt-induced nitrate reductase (NR) activity loss. Legume Res. 2015, 38, 202–208. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, R.K.; Mahata, K.R.; Singh, D.P. Differential responses of antioxidant system and photosynthetic characteristics in four rice cultivars differing in sensitivity to NaCl stress. Acta Physiol. Plant. 2013, 35, 2915–2926. [Google Scholar] [CrossRef]
- Meena, V.D.; Dotaniya, M.L.; Coumar, V.; Rajendiran, S.; Kundu, S.; Subba Rao, A. A Case for silicon fertilization to improve crop yields in tropical soils. Biol. Sci. 2014, 84, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Kafi, M.; Rahimi, Z. Effect of salinity and silicon on root characteristics, growth, water status, proline content and ion accumulation of purslane (Portulaca oleracea L.). Soil Sci. Plant Nutr. 2011, 57, 341–347. [Google Scholar] [CrossRef]
- Ahmad, R.; Zaheer, S.H.; Ismail, S. Role of silicon in salt tolerance of wheat (Tritium aestivum L.). Plant Sci. 1992, 85, 43–50. [Google Scholar] [CrossRef]
- Liang, Y.C.; Shen, Q.R.; Shen, Z.G.; Ma, T.S. Effects of silicon on salinity tolerance of two barley cultivars. J. Plant Nutr. 1996, 19, 173–183. [Google Scholar] [CrossRef]
- Liang, Y.C. Effects of Si on leaf ultrastructure, chlorophyll content and photosynthetic activity in barley under salt stress. Pedosphere 1998, 8, 289–296. [Google Scholar]
- Liang, Y.C.; Zhang, W.H.; Chen, Q.; Ding, R.X. Effects of silicon on tonoplast Hþ –ATPase and Hþ-PPase activity, fatty acid composition and fluidity in roots of salt stressed barley (Hordeum vulgare L.). Environ. Exp. Bot. 2005, 53, 29–37. [Google Scholar] [CrossRef]
- Yeo, A.R.; Flowers, S.A.; Rao, G.; Welfare, K.; Senanayake, N.; Flowers, T.J. Silicon reduces sodium uptake in rice (Oryza sativa L.) in saline conditions and this is accounted for by a reduction in the transpirational bypass flow. Plant Cell Environ. 1999, 22, 559–565. [Google Scholar] [CrossRef]
- Zhu, Z.J.; Wei, G.Q.; Li, J.; Qian, Q.Q.; Yu, J.Q. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Al-Aghabary, K.; Zhu, Z.; Shi, Q.H. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. J. Plant Nutr. 2004, 27, 2101–2115. [Google Scholar] [CrossRef]
- Gunes, A.; Inal, A.; Bagci, E.G.; Pilbeam, D.J. Silicon-mediated changes of some physiological and enzymatic parameters symptomatic for oxidative stress in spinach and tomato grown in sodic-B toxic soil. Plant Soil 2007, 290, 103–114. [Google Scholar] [CrossRef]
- Hashemi, A.; Abdolzadeh, A.; Sadeghipour, H.R. Beneficial effects of silicon nutrition in alleviating salinity stress in hydroponically grown canola, Brassica napus L., plants. Soil Sci. Plant Nutr. 2010, 56, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef]
- Dhiman, P.; Rajora, N.; Bhardwaj, S.; Sudhakaran, S.S.; Kumar, A.; Raturi, G.; Chakraborty, K.; Gupta, O.P.; Devanna, B.N.; Tripathi, D.K.; et al. Fascinating role of silicon to combat salinity stress in plants: An updated overview. Plant Physiol. Biochem. 2021, 162, 110–123. [Google Scholar] [CrossRef]
- Monica, R.C.; Cremonini, R. Nanoparticles and higher plants. Caryology 2009, 62, 61–165. [Google Scholar] [CrossRef] [Green Version]
- Cheng, G.; Chang, T.; Qin, Q.; Huang, H.; Zhu, Y. Mechanical properties of silicon carbide nanowires: Effect of size-dependent defect density. Nano Lett. 2014, 14, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Torney, F.; Trewyn, B.G.; Lin, V.S.Y.; Wang, K. Mesoporous silica nanoparticles deliver DNA and chemicals into plants. Nat. Nanotechnol. 2007, 2, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Almutairi, K.F.; Alotaibi, M.; Shami, A.; Alhammad, B.A.; Battaglia, M.L. Nano-fertilization as an emerging fertilization technique: Why can modern agriculture benefit from its use? Plants 2021, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bao-Shan, L.; Chun-hui, L.; Li-jun, F.; Shu-chun, Q.; Min, Y. Effects of TMS (nanostructured silicon dioxide) on growth of Changbai larch seedlings. J. For. Res. 2004, 15, 138–140. [Google Scholar] [CrossRef]
- Badawy, S.A.; Zayed, B.A.; Bassiouni, S.M.A.; Mahdi, A.H.A.; Majrashi, A.; Ali, E.F.; Seleiman, M.F. Influence of nano silicon and nano selenium on root characters, growth, ion selectivity, yield, and yield components of rice (Oryza sativa L.) under salinity conditions. Plants 2021, 10, 1657. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–668. [Google Scholar] [CrossRef] [Green Version]
- Hanafy, A.A.H.; Harb, E.M.; Higazy, M.A.; Morgan, S.H. Effect of silicon and boron foliar applications on wheat plants grown under saline soil conditions. Inter. J. Agric. Res. 2008, 3, 1–26. [Google Scholar]
- Abdul Qados, A.M.S.; Moftah, A.E. Influence of Silicon and Nano-Silicon on Germination, Growth and Yield of Faba Bean (Vicia faba L.) Under Salt Stress Conditions. Am. J. Exp. Agric. 2015, 5, 509–524. [Google Scholar]
- Ghassemi-Golezani, K.; Nikpour-Rashidabad, N.; Zehtab-Salmasi, S. Physiological performance of pinto bean cultivars under salinity. Inter. J. Plant Anim. Environ. Sci. 2012, 2, 223–228. [Google Scholar]
- Farhoudi, R.; Tafti, M.M. Effect of salt stress on seedlings growth and ions homeostasis of soybean (Glycine max) cultivars. Adv. Environ. Biol. 2011, 5, 2522–2526. [Google Scholar]
- Zhani, K.; Elouer, M.A.; Aloui, H.; Hannachi, C. Selection of a salt tolerant Tunisian cultivar of chili pepper (Capsicum frutescens). EurAsian J. BioSci. 2012, 6, 47–59. [Google Scholar] [CrossRef]
- Kanwal, H.; Ashraf, M.; Hameed, M. Water relations and ionic composition in the seedlings of some newly developed and candidate cultivars of wheat (Triticum aestivum L.) under saline conditions. Pak. J. Bot. 2013, 45, 1221–1227. [Google Scholar]
- Ghezal, N.; Rinez, I.; Sbai, H.; Saad, I.; Farooqd, M.; Rinez, A.; Zribi, I.; Haouala, R. Improvement of Pisum sativum salt stress tolerance by bio-priming their seeds using Typha angustifolia leaves aqueous extract. S. Afr. J. Bot. 2016, 105, 240–250. [Google Scholar] [CrossRef]
- Taize, L.; Zeiger, E. Plant Physiology, 4th ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2006. [Google Scholar]
- Epstein, E. Silicon. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- Agurie, S.; Agata, W.; Kubota, F.; Kaufman, P.B. Physiological role of silicon in photosynthesis and dry matter production in rice plants. Crop Sci. 1992, 61, 200–206. [Google Scholar]
- Hossain, M.T.; Ryuji, M.; Soga, K.; Wakabayashi, K.; Kamisaka, S.; Fuji, S.; Yamamoto, R.; Hoson, T. Growth promotion and increase in cell wall extensibility by silicon in rice and some Poaceae seedlings. J. Plant Res. 2002, 115, 23–27. [Google Scholar] [CrossRef]
- Matichenkov, V.V.; Bocharnikova, E.A.; Ammosova, J.M. The influence of silicon fertilizers on the plants and soils. Agrochemica 2001, 12, 30–37. [Google Scholar]
- Rodrigues, F.A.; Vale, F.X.R.; Korndorfer, G.H.; Prabhu, A.S.; Datnoff, L.E.; Oliveira, A.M.A.; Zambolim, L. Influence of silicon on sheath blight of rice in Brazil. Crop Prot. 2003, 22, 23–29. [Google Scholar] [CrossRef]
- Ma, J.F. Role of silicon in enhancing the resistance of plants to biotic and abiotic stresses. Soil Sci. Plant Nutr. 2004, 50, 11–18. [Google Scholar] [CrossRef]
- Ali, A.; Basra, S.M.A.; Ahmad, R.; Wahid, A. Optimizing silicon application to improve salinity tolerance in wheat. Soil Environ. 2009, 28, 136–144. [Google Scholar]
- Romero-Aranda, M.R.; Jurado, O.; Cuartero, J. Silicon alleviates the deleterious salt effect on tomato plant growth by improving plant water status. J. Plant Physiol. 2006, 163, 847–855. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.Z.; Liu, Y.H. Effects of silicon on growth of maize seedlings under salt stress. Agro-Environ. Prot. 2001, 20, 38–40. [Google Scholar]
- Tahir, M.A.; Rammatullah, A.; Ashraf, T.M.; Kanwal, S.; Maqsood, M.A. Beneficial effects of silicon in wheat (Triticum aestivum) under salinity stress. Pak. J. Bot. 2006, 38, 1715–1722. [Google Scholar]
- Bradbury, M.; Ahmad, R. The effect of silicon on the growth of Prosopis juliflora growing in saline soil. Plant Soil. 1990, 125, 71–74. [Google Scholar] [CrossRef]
- Kamenidou, S.; Cavins, T.J.; Marek, S. Evaluation of silicon as a nutritional supplement for greenhouse zinnia production. Sci. Hort. 2009, 119, 297–301. [Google Scholar] [CrossRef]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Cuartero, J.; Yeo, A.R.; Flowers, T.J. Selection of donors for salt-tolerance in tomato using physiological traits. New Phytol. 1992, 121, 63–69. [Google Scholar] [CrossRef]
- Gong, H.; Chen, K.; Chen, G.; Wang, S.; Zhang, C. Effects of silicon on growth of wheat under drought. J. Plant Nutr. 2003, 26, 1055–1063. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, L.; Higgs, D. Effect of silicon on plant growth and mineral nutrition of maize grown under water-stress conditions. J. Plant Nutr. 2006, 29, 1469–1480. [Google Scholar] [CrossRef]
- Shahzade, M. Growth-Related Changes in Subcellularion and Protein Patterns in Maize and Field Bean Leaves under Salt Stress. Master’s Thesis, Institute of Plant Nutrition and Soil Science, University of Kiel, Kiel, Germany, 2012. [Google Scholar]
- Oertli, J.J. Extracellular salt accumulation, a possible mechanism of salt injury in plants. Soil Sci. 1968, 12, 461–469. [Google Scholar]
- Flowers, T.J.; Flowers, S.A.; Hajibagheri, M.A.; Yeo, A. Ion accumulation in the cells walls of rice plants growing under saline conditions: Evidence for Oertli hypothesis. Plant Cell Environ. 1991, 14, 319–325. [Google Scholar] [CrossRef]
- Barros, L.U.; da Cunha, C.F.; Miguens, F.C. Structure, morphology, and composition of silicon biocomposites in the palm tree Syagrus coronata (Mart.). Becc Protoplasma 2002, 220, 89–96. [Google Scholar]
- Richmond, K.E.; Sussman, M. Got silicon? The non-essential benefical plant nutrient. Curr. Opin. Plant Biol. 2003, 6, 268–272. [Google Scholar] [CrossRef]
- Smith, A.M.; Duan, H.W.; Mohs, A.M.; Nie, S.M. Bioconjugated quantum dots for in vivo molecular and cellular imaging. Adv. Drug Deliv. Rev. 2008, 60, 1226–1240. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Q.; Song, H. Calcium protects Trifolium repens L. plants against cadmium stress. Plant Cell Rep. 2009, 28, 1341–1349. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: Areview. Ecotoxicol. Environ. Saf. 2015, 119, 186–197. [Google Scholar] [CrossRef]
- Suriyaprabha, R.; Karunakaran, G.; Yuvakkumar, R.; Rajendran, V.; Kannan, N. Silica nano particles for increased silica availability in maize (Zea mays L.) seeds under hydroponic conditions. Curr. Nanosci. 2012, 8, 902–908. [Google Scholar] [CrossRef]
- Abbas, W.; Ashraf, M.; Akram, N.A. Alleviation of salt induced adverse effects in eggplant (Solanum melongena L.) by glycine betaine and sugar beet extracts. Sci. Hortic. 2010, 125, 188–195. [Google Scholar] [CrossRef]
- Gay, F.; Maraval, I.; Roques, S.; Gunata, Z.; Boulanger, R.; Audebert, A.; Mestres, C. Effect of salinity on yield and 2-acetyl-1-pyrroline content in the grains of three fragrant rice cultivars (Oryza sativa L.) in Camargue (France). Field Crops Res. 2010, 117, 154–160. [Google Scholar] [CrossRef]
- Kardoni, F.; Mosavi, S.J.S.; Parande, S.; Torbaghan, M.E. Effect of salinity stress and silicon application on yield and component yield of Faba bean (Vicia faba). Inter. J. Agric. Crop Sci. 2013, 6, 814–818. [Google Scholar]
- Miyake, Y.; Takahashi, E. Effect of silicon on the growth and fruit production of strawberry plants in a solution culture. Soil Sci. Plant Nutr. 1986, 32, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Kobraee, S.; Shamsi, K.; Rasekhi, B. Soybean production under water deficit conditions. Ann. Biol. Res. 2011, 2, 423–434. [Google Scholar]
- Ahmad, A.; Afzal, M.; Ahmad, A.U.H.; Tahir, M. Effect of foliar application of silicon on yield and quality of rice (Oryza sativa L). Cercetări. Agron. Moldova. 2013, 3, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Parande, S.; Zamani, G.R.; Zahan, M.H.S.; Ghaderi, M.G. Effects of silicon application on the yield and component of yield in the common bean (Phaseolus vulgaris) under salinity stress. Inter. J. Agron. Plant Prod. 2013, 4, 1574–1579. [Google Scholar]
- Jaberzadeh, A.; Moaveni, P.; Moghadam, H.R.; Zahedi, H. Influence of Bulk and Nanoparticles Titanium Foliar Application on some Agronomic Traits, Seed Gluten and Starch Contents of Wheat Subjected to Water Deficit Stress. Not Bot. Horti. Agrobio. 2013, 41, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M. Biotechnological approach of improving plant salt tolerance using antioxidants as markers. Biotechnol. Adv. 2009, 27, 84–93. [Google Scholar] [CrossRef]
- Anjum, S.A.; Wang, L.; Farooq, M.; Khan, I.; Xue, L. Methyl jasmonate-induced alteration in lipid peroxidation, antioxidative defence system and yield in soybean under drought. J. Agron. Crop Sci. 2011, 197, 296–301. [Google Scholar] [CrossRef]
- Shahid, M.A.; Pervez, M.A.; Balal, R.M.; Mattson, N.S.; Rashid, A.; Ahmad, R.; Ayyub, C.M.; Abbas, T. Brassinosteroid (24-epibrassinolide) enhances growth and alleviates the deleterious effects induced by salt stress in pea (Pisum sativum L.). Aust. J. Crop Sci. 2011, 5, 500–510. [Google Scholar]
- Noreen, Z.; Ashraf, M. Assessment of variation in antioxidative defense system in salt- treated pea (Pisum sativum) cultivars and its putative use as salinity tolerance markers. J. Plant Physiol. 2009, 166, 1764–1774. [Google Scholar] [CrossRef]
- Rouhier, N.; Jacquot, J.P. Getting sick may help plants overcome abiotic stress. New Phytol. 2008, 180, 738–741. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Shim, J.K.; Kim, D.H.; Lee, K.Y.; Lee, I.J. Silicon application to rice root zone influenced the phytohormonal and antioxidant responses under salinity stress. J. Plant Growth Regul. 2014, 33, 137–149. [Google Scholar] [CrossRef]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Shahzad, R.; Lee, I.J. Silicon-mediated mitigation of wounding stress acts by up-regulating the rice antioxidant system. Cereal Res. Commun. 2016, 44, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Torabi, F.; Majd, A.; Enteshari, S. The effect of silicon on alleviation of salt stress in borage (Borago officinalis L.). Soil Sci. Plant Nut. 2015, 61, 788–798. [Google Scholar] [CrossRef] [Green Version]
- Mateos-Naranjo, E.; Gallé, A.; Florez-Sarasa, I.; Perdomo, J.A.; Galmés, J.; Ribas-Carbó, M.; Flexas, J. Assessment of the role of silicon in the Cu tolerance of the C 4 grass Spartina densiflora. J. Plant Physiol. 2015, 178, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Haliem, M.E.; Hegazy, H.S.; Hassan, N.S.; Naguib, D.M. Effect of silica ions and nano silica on rice plants under salinity stress. Ecol. Eng. 2017, 99, 282–289. [Google Scholar] [CrossRef]
- Vatansever, R.; Ozyigit, I.I.; Filiz, E.; Gozukara, N. Genome wide exploration of silicon (Si) transporter genes, Lsi1 and Lsi2 in plants; insights into Si-accumulation status/capacity of plants. BioMetals 2017, 30, 185–200. [Google Scholar] [CrossRef]
- Yamanaka, S.; Takeda, H.; Komatsubara, S.; Ito, F.; Usami, H.; Togawa, E.; Yoshino, K. Structures and physiological functions of silica bodies in the epidermis of rice plants. Appl. Phys. Lett. 2009, 95, 123–703. [Google Scholar] [CrossRef] [Green Version]
- Gong, H.J.; Randall, D.P.; Flowers, T.J. Silicon deposition in the root reduces sodium uptake in rice (Oryza sativa L.) seedlings by reducing bypass flow. Plant Cell Environ. 2006, 29, 1970–1979. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef]
- Trenholm, L.E.; Datnoff, L.E.; Nagara, R.T. Influence of silicon on drought and shade tolerance of St. Augustine grass. Hort. Technol. 2004, 14, 487–490. [Google Scholar] [CrossRef] [Green Version]
- Arshi, A.; Ahmad, A.; Aref, I.M.; Iqbal, M. Comparative studies on antioxidant enzyme action and ion accumulation in soybean cultivars under salinity stress. J. Environ. Biol. 2012, 33, 9–20. [Google Scholar] [PubMed]
- Hertwig, B.; Streb, P.; Feierabend, J. Light dependence of catalase synthesis and degradation in leaves and the influence of interfering stress conditions. Plant Physiol. 1992, 100, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Alscher, R.G.; Donahue, J.L.; Cramer, C.L. Reactive oxygen species and antioxidants: Relationship in green cells. Physiol. Plant. 1997, 100, 224–233. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Qureshi, M.I.; Abdin, M.Z.; Qadir, S.; Iqbal, M. Lead-induced oxidative stress and metabolic alterations in Cassia angustifolia Vahl. Biol. Plant. 2007, 51, 121–128. [Google Scholar] [CrossRef]
- Liang, Y.C.; Chen, Q.; Liu, Q.; Zhang, W.H.; Ding, R.X. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef]
- Noreen, Z.; Ashraf, M.; Akram, N.A. Salt-induced regulation of some key antioxidant enzymes and physiobiochemical phenomena in five diverse cultivars of turnip (Brassica rapa L.). J. Agron. Crop Sci. 2010, 196, 273–285. [Google Scholar]
- Kim, Y.H.; Khan, A.L.; Waqas, M.; Lee, I.J. Silicon regulates antioxidant activities of crop plants under abiotic-induced oxidative stress: A review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [Green Version]
- Beadle, C.L. Growth analysis. In Photosynthesis and Production in a Changing Environment, A Field and Laboratory Manual; Hall, D.C., Scurlock, J.M.O., Bolhar-Nordenkampf, H.R., Leegod, R.C., Long, S.P., Eds.; Chapman & Hall: London, UK, 1993; pp. 36–46. [Google Scholar]
- Ray, S.; Choudhuri, M.A. Regulation of flag leaf senescence in rice by nutrients and its impacts on yield. RISO 1980, 29, 9–14. [Google Scholar]
- Motsara, M.R.; Roy, R.N. Guide to Laboratory Establishment for Plant Nutrient Analysis; Food and Agriculture Organization of the United Nations: Rome, Italy, 2008; pp. 80–90. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta DeltaC (T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Shaheen, R. Stimulation of antioxidant system and lipid peroxidation by abiotic stresses in leaves of Momordica charantia. Braz. J. Plant Physiol. 2007, 19, 149–161. [Google Scholar] [CrossRef] [Green Version]
- Nishikimi, M.; Roa, N.A.; Yogi, K. Measurement of superoxide dismutase. Biochem. Biophys. Res. Comm. 1972, 46, 849–854. [Google Scholar] [CrossRef]
- Devi, P. Principles and Methods in Plant Molecular Biology, Biochemistry and Genetics, 1st ed.; Agrobios: Jodhpur, India, 2002. [Google Scholar]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Treatment | Shoot Length (cm plant−1) | Shoot Fresh Weight (g plant−1) | Shoot Dry Weight (g plant−1) | Shoot Water Content (g plant−1) | Root Length (cm plant−1) | Root Fresh Weight (g plant−1) | Root Dry Weight (g plant−1) | Root Water Content (g plant−1) | No of Leaves/Plant | Leaf Area (cm2 plant−1) |
|---|---|---|---|---|---|---|---|---|---|---|
| Control | 26.60 bcd | 4.76 bcd | 0.89 cdefg | 3.87 abcdef | 3.45 ab | 0.20 abcd | 0.03 bc | 0.17 ab | 6.00 b | 57.0 l |
| Si | 28.53 ab | 6.28 ab | 1.07 bc | 5.21 ab | 4.67 ab | 0.25 ab | 0.03 bc | 0.21 a | 7.00 ab | 70.62 e |
| NSi | 30.40 a | 7.01 a | 1.68 a | 5.33 a | 4.83 ab | 0.26 a | 0.05 a | 0.21 a | 8.00 a | 68.98 f |
| 100 mM | 22.25 fghi | 4.37 cde | 0.88 cdefg | 3.49 bcdef | 3.80 ab | 0.14 cdef | 0.03 cde | 0.11 bcd | 7.00 ab | 55.04 m |
| 100 mM + Si | 26.20 bcde | 5.78 abc | 1.02 bcd | 4.76 abc | 3.50 ab | 0.18 bcde | 0.03 bc | 0.14 abc | 8.00 a | 60.09 i |
| 100 mM + NSi | 27.30 bc | 6.39 ab | 1.30 b | 5.09 abc | 3.15 b | 0.21 abc | 0.04 ab | 0.17 ab | 8.00 a | 61.12 h |
| 150 mM | 22.00 fghi | 4.22 cde | 0.68 defgh | 3.54 bcdef | 4.80 ab | 0.12 def | 0.03 cde | 0.09 cd | 7.00 ab | 49.94 n |
| 150 mM + Si | 26.00 bcde | 5.57 abc | 0.91 cdef | 4.66 abcd | 4.00 ab | 0.16 cdef | 0.03 bc | 0.12 bcd | 8.00 a | 88.65 a |
| 150 mM + NSi | 24.50 cdef | 4.01 cde | 0.62 efgh | 3.39 cdef | 3.97 ab | 0.19 bcde | 0.03 bcd | 0.15 abc | 8.00 a | 78.89 b |
| 200 mM | 21.00 ghi | 2.95 e | 0.51 h | 2.44 ef | 5.00 ab | 0.09 f | 0.02 de | 0.07 d | 7.00 ab | 36.95 o |
| 200 mM + Si | 23.27 efgh | 3.55 de | 0.57 fgh | 2.98 def | 4.25 ab | 0.14 cdef | 0.03 cde | 0.11 bcd | 8.00 a | 73.88 d |
| 200 mM + NSi | 23.30 efgh | 5.68 abc | 0.95 cde | 4.73 abc | 4.53 ab | 0.14 cdef | 0.02 cde | 0.11 bcd | 7.00 ab | 76.49 c |
| 250 mM | 19.50 i | 2.74 e | 0.53 gh | 2.21 f | 5.63 a | 0.11 ef | 0.02 e | 0.09 cd | 7.00 ab | 59.18 k |
| 250 mM + Si | 20.50 hi | 3.29 de | 0.57 fgh | 2.72 ef | 4.90 ab | 0.13 def | 0.03 cde | 0.10 bcd | 8.00 a | 61.37 g |
| 250 mM + NSi | 24.00 defg | 4.88 bcd | 0.83 cdefgh | 4.05 abcde | 4.90 ab | 0.13 def | 0.02 cde | 0.10 bcd | 7.00 ab | 59.23 j |
| Treatment | Shoot Length (cm plant−1) | Shoot Fresh Weight (g plant−1) | Shoot Dry Weight (g plant−1) | Shoot Water Content (g plant−1) | Root Length (cm plant−1) | Root Fresh Weight (g plant−1) | Root Dry Weight (g plant−1) | Root Water Content (g plant−1) | No of Leaves/Plant | Leaf Area (cm2 plant−1) | No of Flowers/Plant |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 32.60 ab | 5.50 ef | 0.93 efg | 4.57 fg | 4.75 ab | 0.22 a | 0.04 e | 0.18 a | 10.00 ab | 109.06 b | 1.00 a |
| Si | 34.00 ab | 7.69 c | 1.51 bc | 6.18 cde | 6.25 a | 0.26 a | 0.06 b | 0.20 a | 10.00 ab | 88.34 f | 2.00 a |
| NSi | 34.10 ab | 13.38 a | 2.23 a | 11.14 a | 6.37 a | 0.27 a | 0.08 a | 0.19 a | 11.00 a | 87.65 g | 2.00 a |
| 100 mM | 31.13 abcd | 6.98 cde | 1.39 bc | 5.59 cdef | 4.90 ab | 0.19 a | 0.04 e | 0.15 a | 10.00 ab | 104.01 c | 1.00 a |
| 100 mM + Si | 31.85 abc | 7.97 c | 1.34 bc | 6.63 c | 3.60 b | 0.22 a | 0.05 cd | 0.17 a | 10.00 ab | 62.00 n | 2.00 a |
| 100 mM + NSi | 36.20 a | 11.28 b | 1.63 b | 9.65 b | 4.20 ab | 0.25 a | 0.05 bc | 0.20 a | 10.00 ab | 68.38 j | 2.00 a |
| 150 mM | 31.00 abcd | 7.25 cd | 1.19 cdef | 6.06 cde | 5.25 ab | 0.23 a | 0.04 de | 0.19 a | 10.00 ab | 89.51 d | 1.00 a |
| 150 mM + Si | 31.33 abcd | 7.66 c | 1.27 cd | 6.40 cd | 4.40 ab | 0.25 a | 0.05 cde | 0.20 a | 10.00 ab | 68.14 k | 2.00 a |
| 150 mM + NSi | 33.00 ab | 10.73 b | 1.63 b | 9.10 b | 4.50 ab | 0.20 a | 0.05 cd | 0.15 a | 10.00 ab | 89.13 e | 2.00 a |
| 200 mM | 25.40 def | 4.98 f | 0.87 fg | 4.10 g | 5.30 ab | 0.15 a | 0.04 e | 0.11 a | 9.00 b | 76.71 h | 1.00 a |
| 200 mM + Si | 25.60 def | 4.65 f | 0.89 fg | 3.75 g | 4.90 ab | 0.14 a | 0.04 de | 0.10 a | 9.00 b | 67.06 l | 2.00 a |
| 200 mM + NSi | 28.87 bcde | 8.04 c | 1.25 cde | 6.80 c | 5.35 ab | 0.17 a | 0.05 cde | 0.12 a | 10.00 ab | 135.04 a | 2.00 a |
| 250 mM | 21.40 f | 4.70 f | 0.85 g | 3.85 g | 6.20 a | 0.14 a | 0.04 e | 0.11 a | 9.00 b | 66.33 m | 2.00 a |
| 250 mM + Si | 23.00 ef | 5.89 def | 0.85 g | 5.05 defg | 5.30 ab | 0.37 a | 0.04 e | 0.33 a | 10.00 ab | 72.85 i | 2.00 a |
| 250 mM + NSi | 26.25 cdef | 5.82 def | 0.99 defg | 4.83 efg | 5.55 ab | 0.13 a | 0.04 e | 0.09 a | 10.00 ab | 56.54 o | 2.00 a |
| Treatment | Shoot Length (cm plant−1) | Plant Height (cm plant−1) | Pod Length (cm plant−1) | Pod Weight (g plant−1) | Crop Yield (g plant−1) | No of Pods /Plant | No of Seeds /Pod | Seed Weight (g pod−1) | Total Seed Yield (g plant−1) | Straw Yield (g plant−1) | Harvest Index | Crop Index | Mobilization Index |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 36.00 abc | 42.35 ab | 10.20 ab | 6.13 c | 7.94 abcd | 3.00 bc | 7.00 ab | 3.53 abc | 3.37 abcd | 6.81 abc | 0.48 ab | 0.32 ab | 0.05 a |
| Si | 34.00 abcd | 37.90 abcd | 10.10 abc | 8.09 b | 9.02 abc | 3.00 bc | 7.00 ab | 3.97 a | 4.10 a | 6.98 abc | 0.59 a | 0.37 a | 0.05 a |
| NSi | 39.55 a | 45.10 a | 10.35 a | 9.91 a | 11.47 a | 5.00 a | 8.00 a | 3.92 ab | 3.95 ab | 8.62 a | 0.46 abc | 0.31 ab | 0.04 a |
| 100 mM | 33.60 abcd | 37.70 abcd | 9.03 bcd | 4.35 de | 7.89 abcd | 3.00 bc | 6.00 ab | 2.02 def | 2.54 abcde | 6.33 abcd | 0.40 abc | 0.29 abc | 0.05 a |
| 100 mM + Si | 34.83 abc | 38.17 abcd | 10.10 abc | 7.82 b | 8.44 abcd | 3.00 bc | 7.00 ab | 2.75 bcd | 3.59 abc | 6.93 abc | 0.46 abc | 0.30 ab | 0.04 a |
| 100 mM + NSi | 37.00 ab | 41.75 ab | 9.80 abc | 7.97 b | 10.17 ab | 4.00 ab | 7.00 ab | 3.74 ab | 2.95 abcde | 8.18 a | 0.37 abc | 0.27 abc | 0.03 a |
| 150 mM | 32.95 abcd | 36.70 bcd | 8.50 de | 3.46 e | 5.70 cde | 1.00 d | 6.00 ab | 1.51 defg | 1.55 def | 4.91 cde | 0.31 abc | 0.23 bc | 0.05 a |
| 150 mM + Si | 34.13 abcd | 37.97 abcd | 9.85 abc | 6.14 c | 6.55 bcde | 3.00 bc | 7.00 ab | 2.39 cde | 2.42 abcde | 6.14 abcd | 0.47 abc | 0.30 ab | 0.06 a |
| 150 mM + NSi | 35.00 abc | 41.4 abc | 9.05 bcd | 5.65 cd | 7.18 bcd | 3.00 bc | 7.00 ab | 2.48 cde | 2.38 abcde | 8.10 ab | 0.29 bc | 0.23 bc | 0.03 a |
| 200 mM | 29.50 cd | 35.65 bcd | 8.30 de | 3.42 e | 4.88 de | 1.00 d | 6.00 ab | 1.02 fg | 1.44 ef | 4.02 de | 0.36 abc | 0.26 abc | 0.06 a |
| 200 mM + Si | 33.25 abcd | 37.00 bcd | 9.05 bcd | 4.84 cde | 5.64 cde | 3.00 bc | 7.00 ab | 2.05 def | 2.05 cdef | 5.59 bcd | 0.37 abc | 0.27 abc | 0.05 a |
| 200 mM + NSi | 32.10 abcd | 35.80 bcd | 8.90 cde | 4.74 cde | 5.72 cde | 3.00 bc | 6.00 ab | 2.04 def | 2.18 bcdef | 6.06 abcd | 0.45 abc | 0.29 abc | 0.07 a |
| 250 mM | 26.93 d | 33.50 d | 7.70 e | 1.91 f | 2.82 e | 1.00 d | 5.00 b | 0.49 g | 0.49 f | 2.68 e | 0.20 c | 0.16 c | 0.07 a |
| 250 mM + Si | 29.00 cd | 36.50 bcd | 9.00 bcd | 3.88 e | 4.99 de | 2.00 cd | 6.00 ab | 1.61 defg | 1.83 cdef | 3.93 de | 0.47 abc | 0.32 ab | 0.08 a |
| 250 mM + NSi | 29.67 bcd | 34.00 cd | 8.47 de | 3.51 e | 5.35 cde | 3.00 bc | 6.00 ab | 1.31 efg | 1.73 def | 4.43 cde | 0.39 abc | 0.28 abc | 0.06 a |
| Treatment | Na+ | K+ | Na+/K+ Ratio | |||
|---|---|---|---|---|---|---|
| Root | Shoot | Root | Shoot | Root | Shoot | |
| Control | 9.80 k | 7.97 l | 12.96 f | 21.84 c | 0.76 j | 0.36 l |
| Si | 10.20 j | 8.90 k | 11.06 h | 24.89 a | 0.92 h | 0.36 l |
| NSi | 9.80 k | 7.97 l | 16.16 d | 19.78 e | 0.61 m | 0.40 k |
| 100 mM | 12.19 h | 17.21 f | 6.80 l | 10.50 k | 1.79 d | 1.64 d |
| 100 mM + Si | 10.20 j | 13.20 i | 10.50 i | 12.90 j | 0.97 g | 1.02 g |
| 100 mM + NSi | 9.80 k | 11.20 j | 13.20 e | 15.87 h | 0.74 jk | 0.71 j |
| 150 mM | 20.24 c | 18.21 e | 9.80 j | 8.32 l | 2.07 c | 2.19 c |
| 150 mM + Si | 12.19 h | 15.22 g | 10.50 i | 13.22 i | 1.16 e | 1.15 f |
| 150 mM + NSi | 11.19 i | 14.23 h | 16.20 c | 19.40 f | 0.69 l | 0.73 j |
| 200 mM | 26.23 b | 19.21 d | 11.80 g | 3.80 m | 2.22 b | 5.06 b |
| 200 mM + Si | 13.24 f | 23.23 b | 16.20 c | 19.33 g | 0.82 i | 1.20 e |
| 200 mM + NSi | 12.80 g | 19.22 d | 17.88 b | 23.41 b | 0.72 kl | 0.82 h |
| 250 mM | 32.24 a | 21.22 c | 9.06 k | 2.90 n | 3.56 a | 7.32 a |
| 250 mM + Si | 19.22 d | 26.23 a | 17.88 b | 21.54 d | 1.07 f | 1.22 e |
| 250 mM + NSi | 16.20 e | 19.22 d | 22.25 a | 24.90 a | 0.73 jk | 0.77 i |
| Target Gene | Forward Primer 5′–3′ | Reverse Primer 5′–3′ |
|---|---|---|
| SOD | GCGACATTCTTCCGGCTTTC | GCCCAGCCTGAACCAAATTG |
| POD | CTAGTTGCTCTTTCCGGTGC | GGTTTGTTCCACTTCCACCC |
| CAT | TCACAGGGATGAGGAGGTCA | TGGATCGGTGTCGGATAAAGC |
| PsLecRLK | TGAAGGATGGGAAGTGGAAG | CTCGACCGGTTTTGATCACT |
| Actin | GCTGTCCTCTCCCTCTATGCA | GCCGAGGTGGTGAACATATACC |
| Tubulin | GTACACTGGTGAAGGCATGGA | ACTGCTGAACACACTTACACG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ismail, L.M.; Soliman, M.I.; Abd El-Aziz, M.H.; Abdel-Aziz, H.M.M. Impact of Silica Ions and Nano Silica on Growth and Productivity of Pea Plants under Salinity Stress. Plants 2022, 11, 494. https://doi.org/10.3390/plants11040494
Ismail LM, Soliman MI, Abd El-Aziz MH, Abdel-Aziz HMM. Impact of Silica Ions and Nano Silica on Growth and Productivity of Pea Plants under Salinity Stress. Plants. 2022; 11(4):494. https://doi.org/10.3390/plants11040494
Chicago/Turabian StyleIsmail, Lamiaa M., Magda I. Soliman, Mohammed H. Abd El-Aziz, and Heba M. M. Abdel-Aziz. 2022. "Impact of Silica Ions and Nano Silica on Growth and Productivity of Pea Plants under Salinity Stress" Plants 11, no. 4: 494. https://doi.org/10.3390/plants11040494
APA StyleIsmail, L. M., Soliman, M. I., Abd El-Aziz, M. H., & Abdel-Aziz, H. M. M. (2022). Impact of Silica Ions and Nano Silica on Growth and Productivity of Pea Plants under Salinity Stress. Plants, 11(4), 494. https://doi.org/10.3390/plants11040494