
Evaluation of Barley Cultivars for Competitive Traits in Southern New South Wales





Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description and Experimental Design
2.2. Crop Establishment
2.3. Crop Assessments and Data Collection
2.4. Data Analysis
3. Results
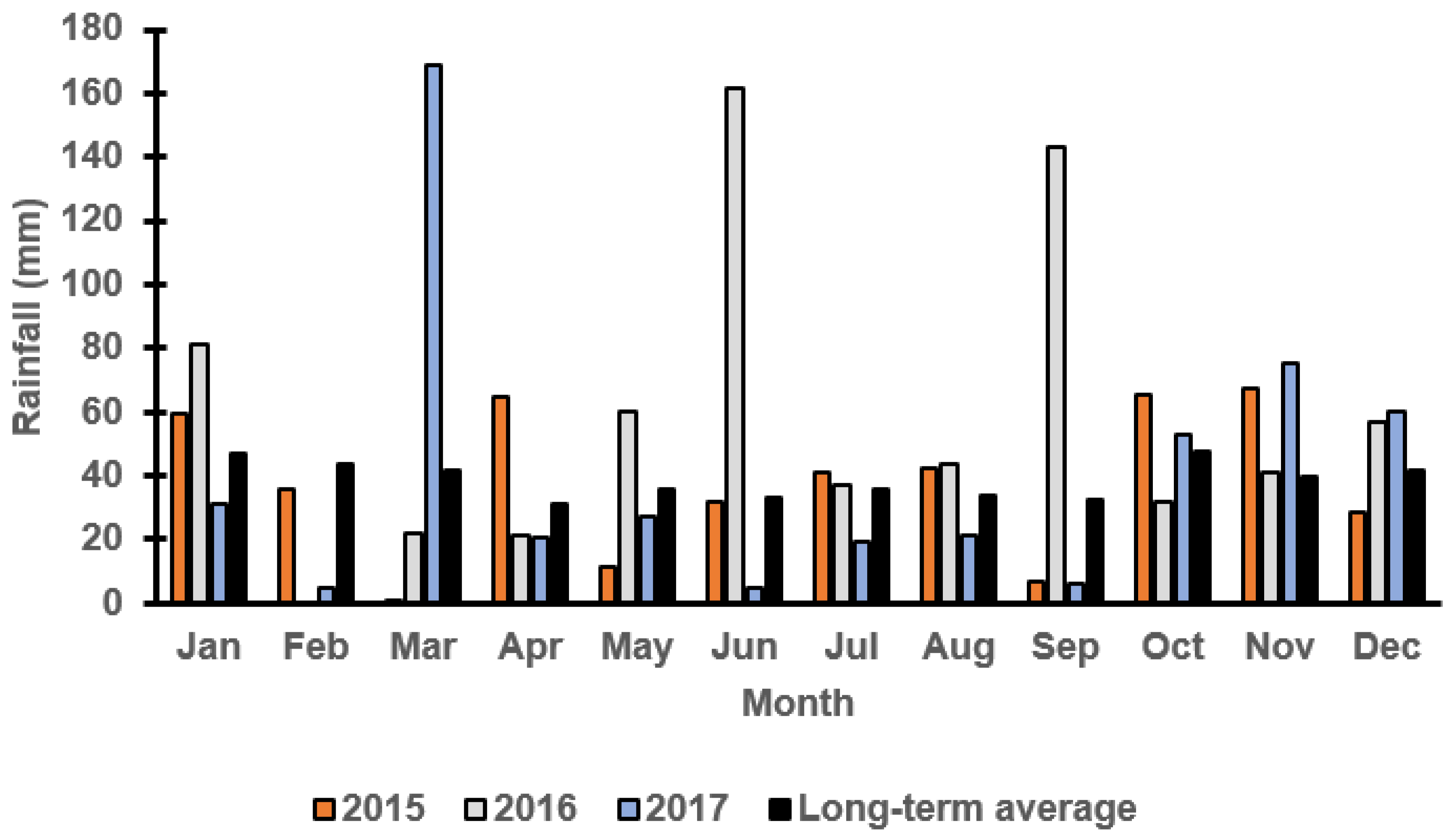
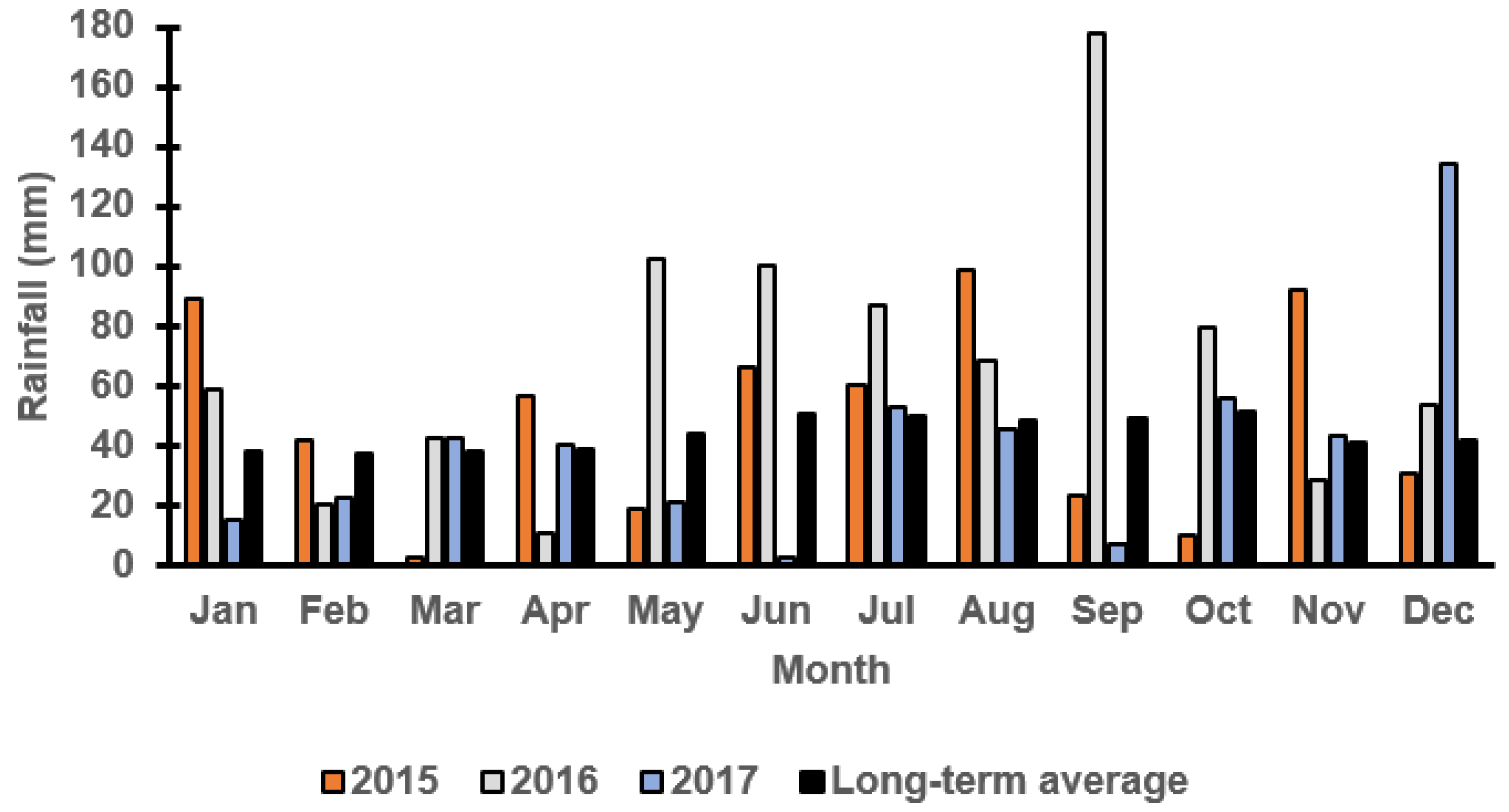
3.1. Monthly Rainfall at Both Locations
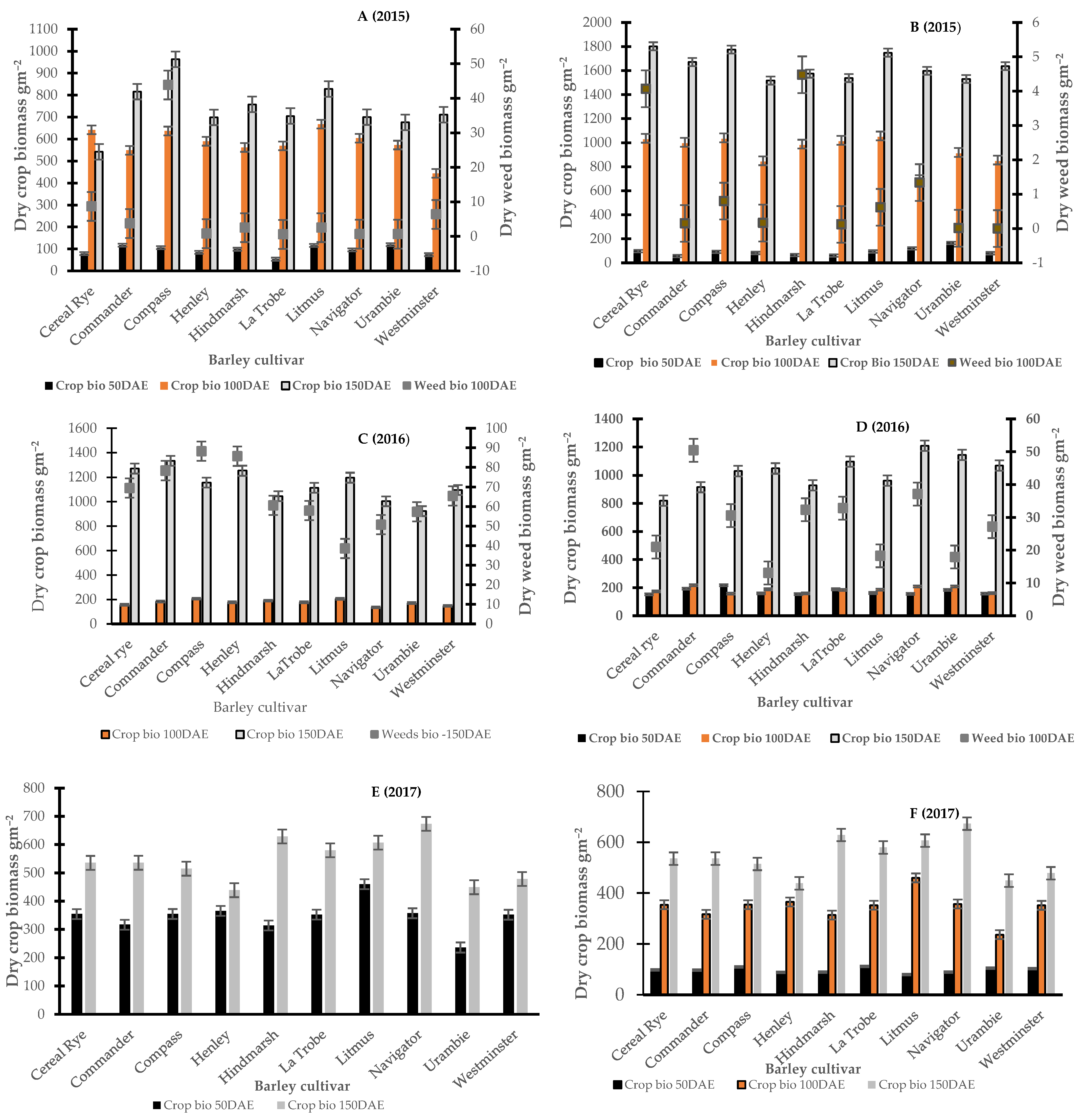
3.2. Early Competitive Ability of Barley to Suppress Weeds Based on Biomass Accumulation
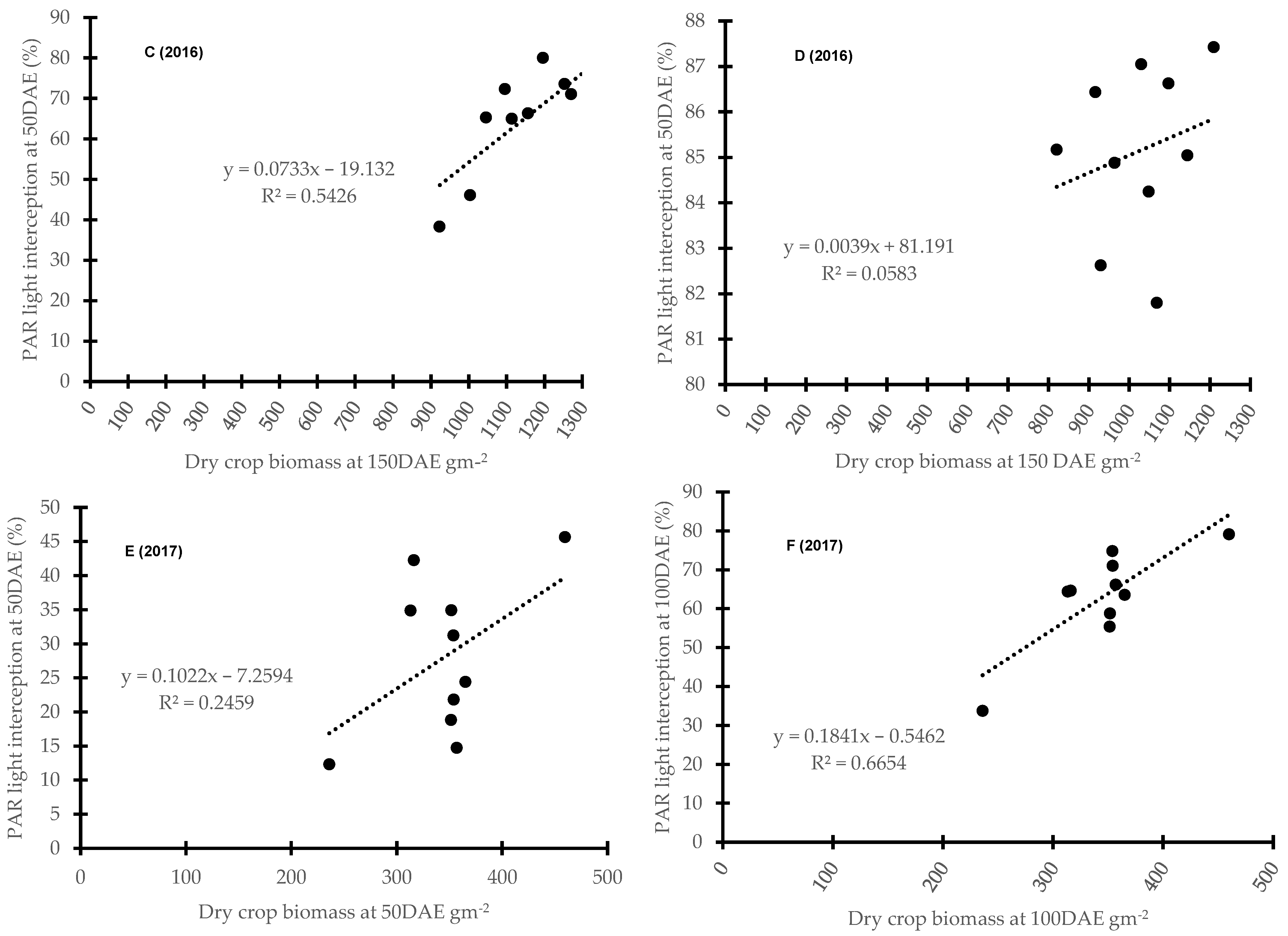
3.3. Barley Canopy Architecture Traits Contributing to Weed Suppression at Different Growth Stages
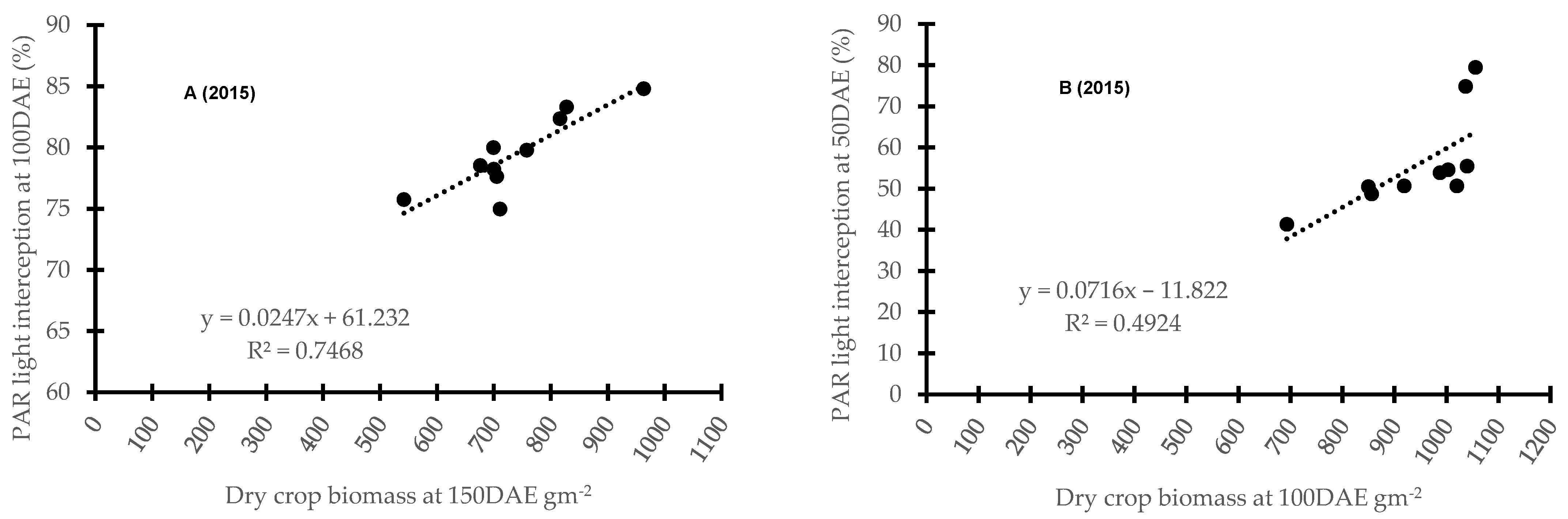
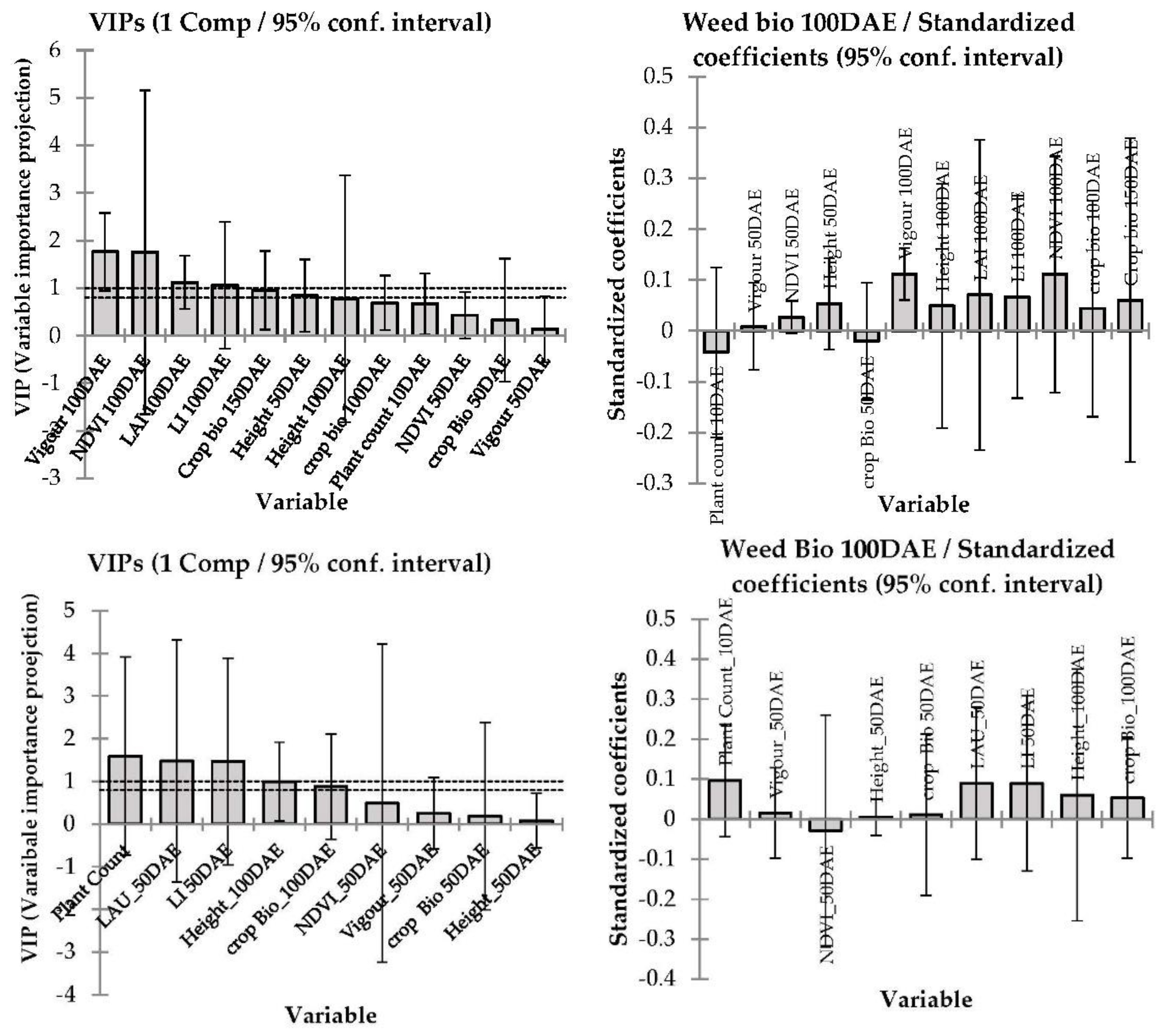
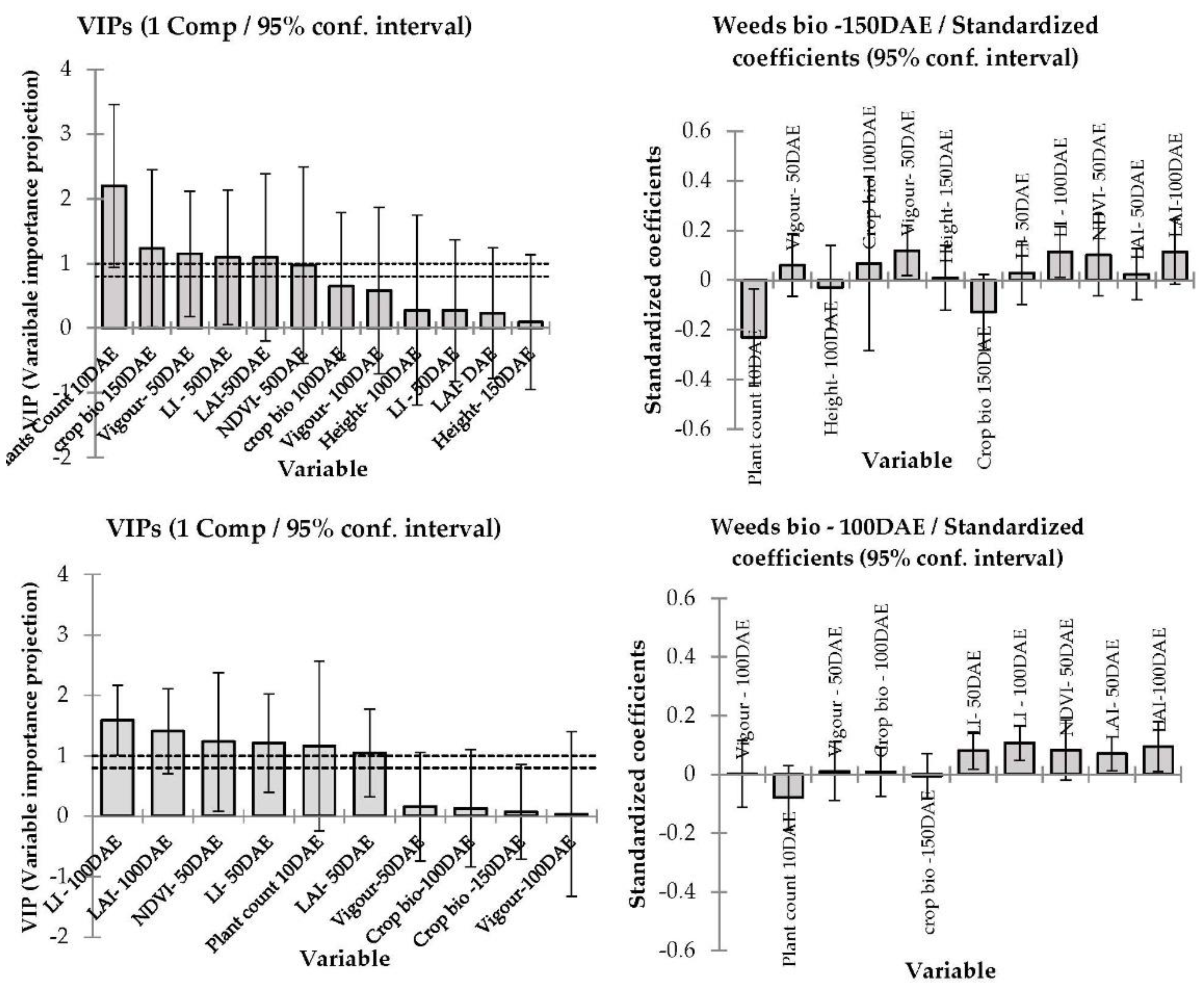
3.4. Correlation between Barley Cultivar Selected Canopy Traits and Competitive Ability against Weeds
3.5. Barley Cultivar Grain Yield and Tolerance against Weeds
4. Discussion
4.1. Early Competitive Ability of Barley to Suppress Weeds Based on Biomass Accumulation
4.2. Barley Canopy Architecture Traits Contributing to Weed Suppression at Different Growth Stages
4.3. Modelling—Correlation between Barley Canopy Traits and Competitive Ability against Weeds
4.4. Barley Cultivar Grain Yield and Tolerance against Weeds
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jabran, K.; Mahajan, G.; Sardana, V.; Chauhan, B.S. Allelopathy for weed control in agricultural systems. Crop Prot. 2015, 72, 57–65. [Google Scholar] [CrossRef]
- Mwendwa, J.M.; Brown, W.B.; Weidenhamer, J.D.; Weston, P.A.; Quinn, J.C.; Wu, H.; Weston, L.A. Evaluation of commercial wheat cultivars for canopy architecture, early vigour, weed suppression, and yield. Agronomy 2020, 10, 983. [Google Scholar] [CrossRef]
- Wu, H. Integrating Belowground Non-Chemical Approaches for Future Weed Management. In Weed and Pest Control: Molecular Biology, Practices and Environmental Impact; Travlos, I.S., Bilalis, D., Chachalis, D., Eds.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2016. [Google Scholar]
- Heap, I. International Herbicide-Resistant Weed Database. Available online: www.weedscience.com (accessed on 30 April 2021).
- Bajwa, A.A.; Walsh, M.; Chauhan, B.S. Weed management using crop competition in Australia. Crop Prot. 2017, 95, 8–13. [Google Scholar] [CrossRef]
- Llewellyn, R.; Ronning, D.; Ouzman, J.; Walker, S.; Mayfield, A.; Clarke, M. Impact of Weeds on Australian Grain Production: The Cost of Weeds to Australian Grain Growers and the Adoption of Weed Management and Tillage Practices; Grains Research and Development Corporation and CSIRO: Canberra, Australia, 2016. [Google Scholar]
- Mwendwa, J.M.; Brown, W.B.; Wu, H.; Weston, P.A.; Weidenhamer, J.D.; Quinn, J.C.; Weston, L.A. The weed suppressive ability of selected Australian grain crops; case studies from the Riverina region in New South Wales. Crop Prot. 2018, 103, 9–19. [Google Scholar] [CrossRef]
- Andrew, I.K.S.; Storkey, J.; Sparkes, D.L. A review of the potential for competitive cereal cultivars as a tool in integrated weed management. Weed Res. 2015, 55, 239–248. [Google Scholar] [CrossRef]
- Goldberg, D. Components of Resource Competition in Plant Communities. In Perspectives in Plant Competition; Grace, J.B., Tilman, D., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 27–49. [Google Scholar]
- Tilman, D. Constraints and tradeoffs: Toward a predictive theory of competition and succession. Oikos 1990, 58, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Didon, U.M.E. Variation between barley cultivars in early response to weed competition. J. Agron. Crop Sci. 2002, 188, 176–184. [Google Scholar] [CrossRef]
- Lemerle, D.; Verbeek, B.; Cousens, R.D.; Coombes, N.E. The potential for selecting wheat varieties strongly competitive against weeds. Weed Res. 1996, 36, 505–513. [Google Scholar] [CrossRef]
- Huel, D.G.; Hucl, P. Genotypic variation for competitive ability in spring wheat. Plant Breed. 1996, 115, 325–329. [Google Scholar] [CrossRef]
- Seavers, G.P.; Wright, K.J. Crop canopy development and structure influence weed suppression. Weed Res. 1999, 39, 319–328. [Google Scholar] [CrossRef]
- Wicks, G.A.; Ramsel, R.E.; Nordquist, P.T.; Schmidt, J.W.; Challaiah, R.E. Impact of wheat cultivars on establishment and suppression of summer annual weeds. Agron. J. 1986, 78, 59–62. [Google Scholar]
- Christensen, S. Weed suppression ability of spring barley varieties. Weed Res. 1995, 35, 241–247. [Google Scholar] [CrossRef]
- Liebman, M.; Davis, A.S. Integration of soil, crop and weed management in low-external-input farming systems. Weed Res. 2000, 40, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Hodgdon, E.A.; Warren, N.D.; Smith, R.G.; Sideman, R.G. In-season and carry-over effects of cover crops on productivity and weed suppression. Agron. J. 2016, 108, 1624–1635. [Google Scholar] [CrossRef]
- Hansen, P.K.; Rasmussen, I.A.; Holst, N.; Andreasen, C. Tolerance of four spring barley (Hordeum vulgare) varieties to weed harrowing. Weed Res. 2007, 47, 241–251. [Google Scholar] [CrossRef] [Green Version]
- Bertholdsson, N.-O.; Andersson, S.C.; Merker, A. Allelopathic potential of Triticum spp., Secale spp. and Triticosecale spp. and use of chromosome substitutions and translocations to improve weed suppression ability in winter wheat. Plant Breed. 2012, 131, 75–80. [Google Scholar] [CrossRef]
- Worthington, M.; Reberg-Horton, C. Breeding cereal crops for enhanced weed suppression: Optimising allelopathy and competitive ability. J. Chem. Ecol. 2013, 39, 213–231. [Google Scholar] [CrossRef]
- Bertholdsson, N.O. Variation in allelopathic activity over 100 years of barley selection and breeding. Weed Res. 2004, 44, 78–86. [Google Scholar] [CrossRef]
- Dhima, K.; Vasilakoglou, I.; Gatsis, T.; Eleftherohorinos, I. Competitive interactions of fifty barley cultivars with Avena sterilis and Asperugo procumbens. Field Crops Res. 2010, 117, 90–100. [Google Scholar] [CrossRef]
- Bertholdsson, N.O. Early vigour and allelopathy–two useful traits for enhanced barley and wheat competitiveness against weeds. Weed Res. 2005, 45, 94–102. [Google Scholar] [CrossRef]
- Mwendwa, J.M.; Weston, P.A.; Weidenhamer, J.D.; Fomsgaard, I.S.; Wu, H.; Gurusinghe, S.; Weston, L.A. Metabolic profiling of benzoxazinoids in the roots and rhizosphere of commercial winter wheat genotypes. Plant Soil. 2021, 466, 467–489. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Singh, R.G.; Mahajan, G. Ecology and management of weeds under conservation agriculture: A review. Crop Prot. 2012, 38, 57–65. [Google Scholar] [CrossRef]
- Mwendwa, J.M.; Brown, W.B.; Weston, P.A.; Haque, K.S.; Preston, C.; Weston, L.A. Evaluation of selected commercial oilseed rape cultivars for early vigour, weed suppression and yield in southern New South Wales. Weed Res. 2020, 60, 450–463. [Google Scholar] [CrossRef]
- Rebetzke, G.; Van Herwaarden, A.; Chenu, K.; Moeller, C.; Biddulph, B.; Richards, R.; Rattey, A. Protocols for Experimental Plot Sampling, Handling and Processing of Cereal Experiments: Standardised Methods for Use in Large Agronomic, Physiological and Genetic Field Studies; Prometheus Wiki; CSIRO Publishing: Collingwood, Australia, 2012; pp. 1–46. Available online: http://prometheuswiki.publish.csiro.au/tiki-index.php (accessed on 18 April 2021).
- Matthews, P.; McCaffery, D.; Jenkins, L. Winter Crop Variety Sowing Guide 2015; NSW Department of Primary Industries: Orange, Australia, 2015. [Google Scholar]
- VSN International. GenStat for Windows, 19th ed.; VSN International: Hemel Hempstead, UK, 2018. [Google Scholar]
- Mevik, B.H.; Wehrens, R. The PLS Package: Principal Component and Partial Least Squares Regression in R. J. Stat. Softw. 2007, 18, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Akarachantachote, N.; Seree, C.; Kidakan, S. Cutoff threshold of variable importance in projection for variable selection. Int. J. Pure Appl. Math. 2014, 94, 307–322. [Google Scholar] [CrossRef]
- Coleman, R.K.; Gill, G.S.; Rebetzke, G.J. Identification of quantitative trait loci for traits conferring weed competitiveness in wheat (Triticum aestivum L.). Crop Past. Sci. 2001, 52, 1235–1246. [Google Scholar] [CrossRef]
- Paynter, B.H.; Hills, A.L. Barley and rigid ryegrass (Lolium rigidum) competition is influenced by crop cultivar and density. Weed Technol. 2009, 23, 40–48. [Google Scholar] [CrossRef]
- Worthington, M.; Reberg-Horton, S.C.; Brown-Guedira, G.; Jordan, D.; Weisz, R.; Murphy, J.P. Relative contributions of allelopathy and competitive traits to the weed suppressive ability of winter wheat lines against Italian Ryegrass. Crop Sci. 2015, 55, 57–64. [Google Scholar] [CrossRef]
- Bertholdsson, N.O. Use of multivariate statistics to separate allelopathic and competitive factors influencing weed suppression ability in winter wheat. Weed Res. 2011, 51, 273–283. [Google Scholar] [CrossRef]
- Sardana, V.; Mahajan, G.; Jabran, K.; Chauhan, B.S. Role of competition in managing weeds: An introduction to the special issue. Crop Prot. 2017, 95, 1–7. [Google Scholar] [CrossRef]
- Didon, U.M.E.; Hansson, M.L. Competition between six spring barley (Hordeum vulgare ssp. vulgare L.) cultivars and two weed flora in relation to interception of photosynthetic active radiation. Biol. Agric. Hortic. 2002, 20, 257–274. [Google Scholar]
- Rebetzke, G.; Ingvordsen, C.; Newman, P.; Weston, L.A.; French, B.; Gill, G. Delivering Weed-Competitive, Wheat Breeding Lines to Growers; GRDC Grains Research Update: Wagga Wagga, Australia, 2018; pp. 35–40. [Google Scholar]
- Bertholdsson, N.-O. Breeding spring wheat for improved allelopathic potential. Weed Res. 2010, 50, 49–57. [Google Scholar] [CrossRef]
- Watson, P.R.; Derksen, D.A.; Van Acker, R.C. The ability of 29 barley cultivars to compete and withstand competition. Weed Sci. 2006, 54, 783–792. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wagga Wagga | Condobolin | Description | |
|---|---|---|---|
| 1 | Commander | Commander | Malt |
| 2 | Compass | Compass | Malt and Grazing |
| 3 | Henley | Henley | Malt |
| 4 | Hindmarsh | Hindmarsh | Feed/malt |
| 5 | La Trobe | La Trobe | Malt |
| 6 | Litmus | Litmus | Malt |
| 7 | Navigator | Navigator | Malt |
| 8 | Urambie | Urambie | Feed and Grazing |
| 9 | Westminster | Westminster | Malt |
| 10 | Grazer (Cereal rye) | Grazer (Cereal rye) | Dual purpose (control) |
| Location | Condobolin Yield t ha−1 | Wagga Wagga Yield t ha−1 | ||||
|---|---|---|---|---|---|---|
| Cultivar/Year | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 |
| Cereal Rye | 0.4 | 2.2 | 0.9 | 2.7 | 5.7 | 1.3 |
| Commander | 1.3 | 4.3 | 1.4 | 4.0 | 6.3 | 1.1 |
| Compass | 1.2 | 4.6 | 1.8 | 4.5 | 5.4 | 1.4 |
| Henley | 1.3 | 4.5 | 1.5 | 4.3 | 5.6 | 1.1 |
| Hindmarsh | 1.4 | 3.8 | 1.5 | 5.1 | 5.7 | 1.5 |
| La Trobe | 1.3 | 4.2 | 1.6 | 4.9 | 5.5 | 1.4 |
| Litmus | 1.7 | 4.3 | 1.4 | 4.3 | 5.8 | 1.4 |
| Navigator | 1.1 | 4.7 | 1.6 | 4.4 | 6.1 | 0.8 |
| Urambie | 1.3 | 3.9 | 1.2 | 3.8 | 5.3 | 1.2 |
| Westminster | 1.1 | 3.9 | 1.3 | 3.8 | 6.1 | 1.2 |
| LSD0.05 | 0.47 | 0.70 | 0.28 | 0.50 | 1.70 | 0.13 |
| CV % | 7.4 | 12.1 | 16.8 | 9.4 | 20.6 | 9.13 |
| p-value | >0.01 | >0.01 | >0.01 | >0.01 | NS | >0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mwendwa, J.M.; Brown, W.B.; Weston, P.A.; Weston, L.A. Evaluation of Barley Cultivars for Competitive Traits in Southern New South Wales. Plants 2022, 11, 362. https://doi.org/10.3390/plants11030362
Mwendwa JM, Brown WB, Weston PA, Weston LA. Evaluation of Barley Cultivars for Competitive Traits in Southern New South Wales. Plants. 2022; 11(3):362. https://doi.org/10.3390/plants11030362
Chicago/Turabian StyleMwendwa, James M., William B. Brown, Paul A. Weston, and Leslie A. Weston. 2022. "Evaluation of Barley Cultivars for Competitive Traits in Southern New South Wales" Plants 11, no. 3: 362. https://doi.org/10.3390/plants11030362
APA StyleMwendwa, J. M., Brown, W. B., Weston, P. A., & Weston, L. A. (2022). Evaluation of Barley Cultivars for Competitive Traits in Southern New South Wales. Plants, 11(3), 362. https://doi.org/10.3390/plants11030362