
Genome Size of Life Forms of Araceae—A New Piece in the C-Value Puzzle


Abstract
1. Introduction
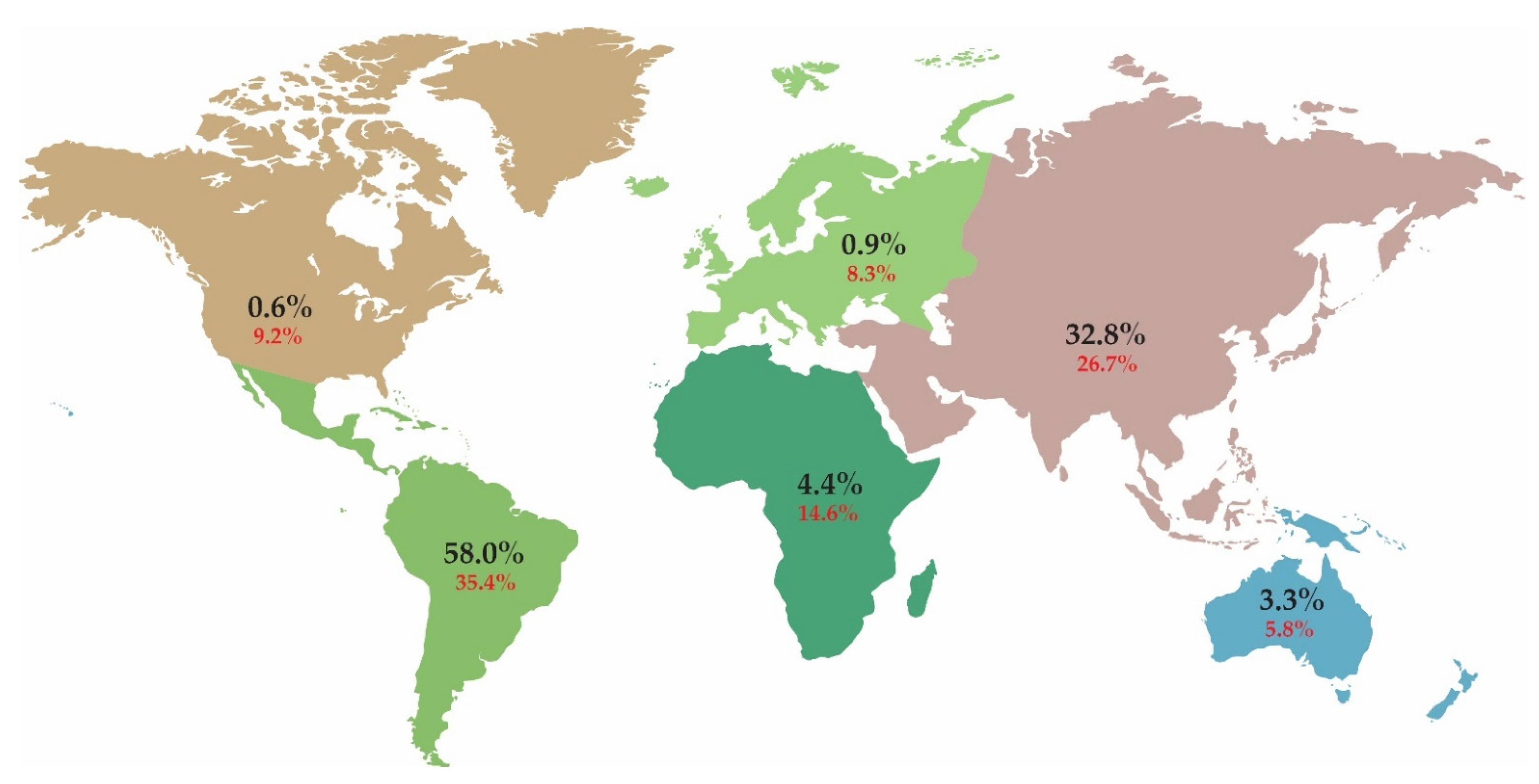
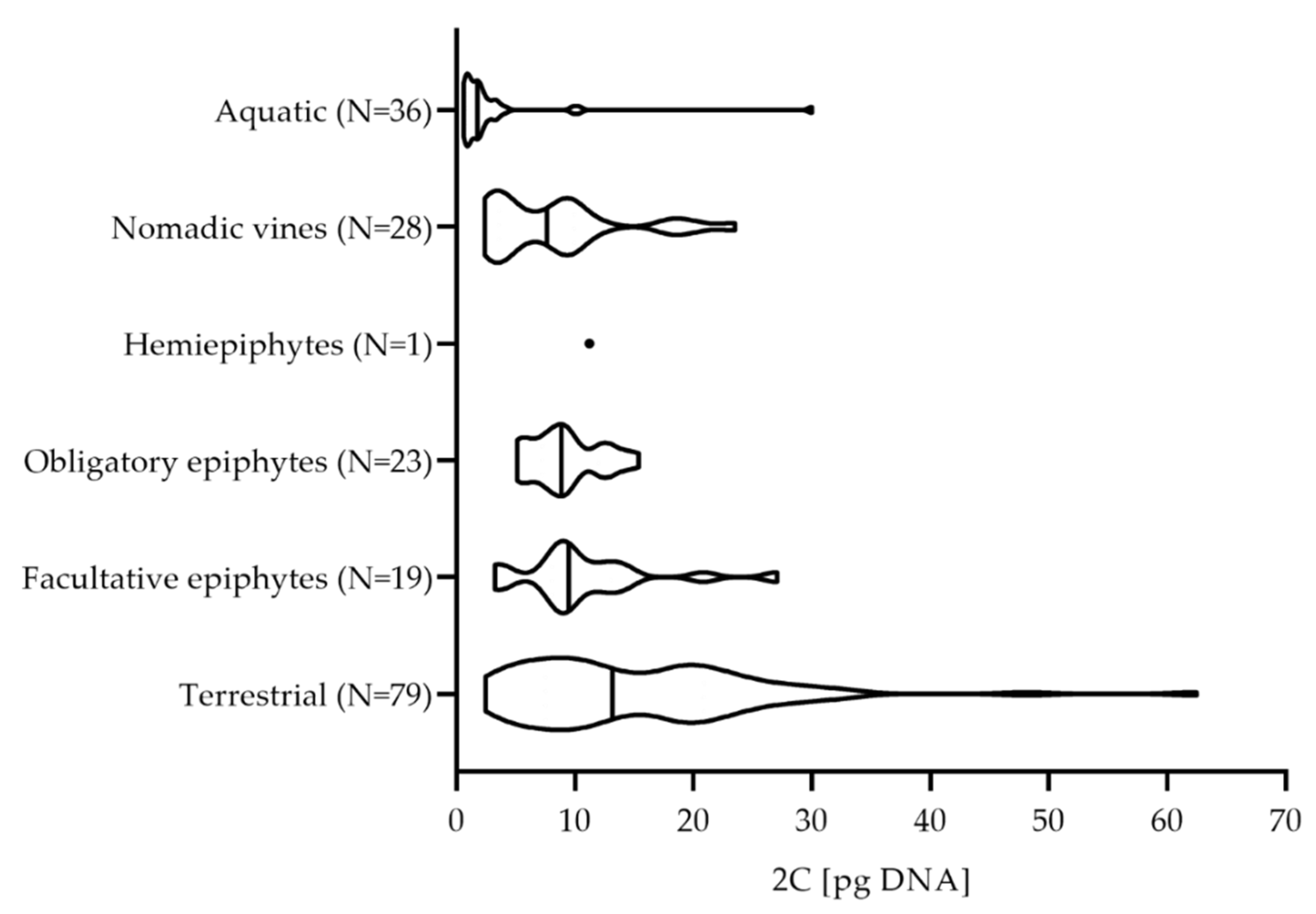
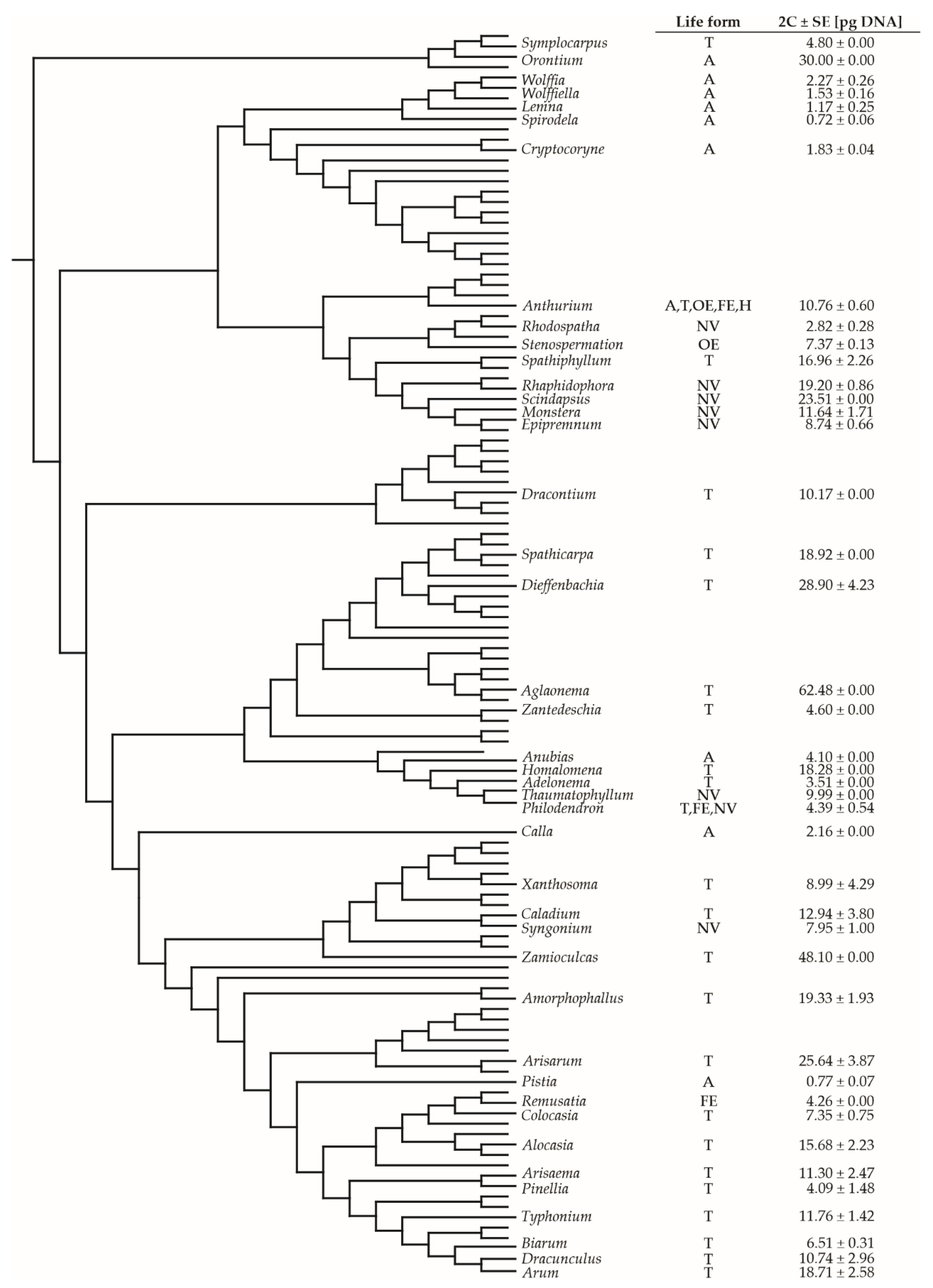
2. Results and Discussion
2.1. C-Value Estimates
2.2. Aroid C-Values Related to Plant Life Forms
3. Conclusions
4. Material and Methods
4.1. Plant Material
4.2. Genome Size Measurement
4.3. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greilhuber, J.; Doležel, J.; Lysak, M.A.; Bennett, M.D. The Origin, Evolution and Proposed Stabilization of the Terms “Genome Size” and “C-Value” to Describe Nuclear DNA Contents. Ann. Bot. 2005, 95, 255–260. [Google Scholar] [CrossRef]
- Swift, H. The Constancy of Desocyribose Nucleic Acid in Plant Nuclei. Proc. Natl. Acad. Sci. USA 1950, 36, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Barow, M.; Jovtchev, G. Endopolyploidy in Plants and Its Analysis by Flow Cytometry. In Flow Cytometry with Plant Cells; Doležel, J., Greilhuber, J., Suda, J., Eds.; Wiley: Weinheim, Germany, 2007; pp. 349–372. ISBN 978-3-527-31487-4. [Google Scholar]
- Erbrich, P. Über Endopolyploidie Und Kernstrukturen in Endospermhaustorien. Österr. Bot. Z. 1965, 112, 197–262. [Google Scholar] [CrossRef]
- Thomas, C.A., Jr. The Genetic Organization of Chromosomes. Annu. Rev. Genet. 1971, 5, 237–256. [Google Scholar] [CrossRef] [PubMed]
- Greilhuber, J.; Leitch, I.J. Genome Size and the Phenotype. In Plant Genome Diversity; Greilhuber, J., Dolezel, J., Wendel, J.F., Eds.; Springer Vienna: Vienna, Austria, 2013; Volume 2, pp. 323–344. ISBN 978-3-7091-1159-8. [Google Scholar]
- Gregory, R.T. Coincidence, Coevolution, or Causation? DNA Content, Cell Size, and the C-Value Enigma. Biol. Rev. 2001, 76, 65–101. [Google Scholar] [CrossRef]
- Orgel, L.E.; Crick, F.H.C.; Sapienza, C. Selfish DNA. Nature 1980, 288, 645–646. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.D. The Duration of Meiosis. Proc. R. Soc. Lond. B Biol. Sci. 1971, 178, 277–299. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Nuclear Volume Control by Nucleoskeletal DNA, Selection for Cell Volume and Cell Growth Rate, and the Solution of the DNA C-Value Paradox. J. Cell Sci. 1978, 36, 247–278. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Economy, Speed and Size Matter: Evolutionary Forces Driving Nuclear Genome Miniaturization and Expansion. Ann. Bot. 2005, 95, 147–175. [Google Scholar] [CrossRef]
- Bennett, M.D.; Bhandol, P.; Leitch, I.J. Nuclear DNA Amounts in Angiosperms and Their Modern Uses—807 New Estimates. Ann. Bot. 2000, 86, 859–909. [Google Scholar] [CrossRef]
- Grime, J. Plant Classification for Ecological Purposes: Is There a Role for Genome Size? Ann. Bot. 1998, 82, 117–120. [Google Scholar] [CrossRef]
- Plant DNA C-Values Database. Available online: https://cvalues.science.kew.org (accessed on 7 January 2022).
- Sessions, S.K. Genome Size. In Brenner’s Encyclopedia of Genetics; Maloy, S., Hughes, K., Eds.; Elsevier: London, UK, 2013; Volume 3, pp. 301–305. ISBN 978-0-08-096156-9. [Google Scholar]
- Lughadha, E.N.; Govaerts, R.; Belyaeva, I.; Black, N.; Lindon, H.; Allkin, R.; Magill, R.E.; Nicolson, N. Counting Counts: Revised Estimates of Numbers of Accepted Species of Flowering Plants, Seed Plants, Vascular Plants and Land Plants with a Review of Other Recent Estimates. Phytotaxa 2016, 272, 82. [Google Scholar] [CrossRef]
- Bennett, M.D.; Leitch, I.J. Nuclear DNA Amounts in Angiosperms: Targets, Trends and Tomorrow. Ann. Bot. 2011, 107, 467–590. [Google Scholar] [CrossRef] [PubMed]
- Plants of the World Online. Available online: https://powo.science.kew.org/ (accessed on 7 January 2022).
- The Überlist of Araceae, Totals for Published and Estimated Number of Species in Aroid Genera. Available online: http://www.aroid.org/genera/20201008Uberlist.pdf (accessed on 28 November 2021).
- Bogner, J. Araceae. In Monocotyledons; Eggli, U., Nyffeler, R., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 457–459. ISBN 978-3-662-56484-4. [Google Scholar]
- Mayo, S.J.; Bogner, J.; Boyce, P.C. The Genera of Araceae; Royal Botanic Gardens: Kew, UK, 1997; ISBN 978-1-900-34722-8. [Google Scholar]
- Croat, T.B. Ecology and Life Forms of Araceae. Aroideana 1988, 11, 4–52. [Google Scholar]
- Sperotto, P.; Acevedo-Rodríguez, P.; Vasconcelos, T.N.C.; Roque, N. Towards a Standardization of Terminology of the Climbing Habit in Plants. Bot. Rev. 2020, 86, 180–210. [Google Scholar] [CrossRef]
- Zotz, G. ‘Hemiepiphyte’: A Confusing Term and Its History. Ann. Bot. 2013, 111, 1015–1020. [Google Scholar] [CrossRef]
- Seifriz, W. The Plant Life of Cuba. Ecol. Monogr. 1943, 13, 375–426. [Google Scholar] [CrossRef]
- Moffett, M.W. What’s “up”? A Critical Look at the Basic Terms of Canopy Biology. Biotropica 2000, 32, 569–596. [Google Scholar] [CrossRef]
- Huber, W.; Weissenhofer, A.; Zamora, N.; Weber, A. Plant Diversity and Biogeography of the Golfo Dulce Region, Costa Rica. Staphia 2008, 88, 97–103. [Google Scholar]
- Bharathan, G.; Lambert, G.; Galbraith, D.W. Nuclear DNA Content of Monocotyledons and Related Taxa. Am. J. Bot. 1994, 81, 381–386. [Google Scholar] [CrossRef]
- Zonneveld, B.J.M.; Leitch, I.J.; Bennett, M.D. First Nuclear DNA Amounts in More than 300 Angiosperms. Ann. Bot. 2005, 96, 229–244. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Harijati, N.; Liu, E.; Jin, S.; Diao, Y.; Hu, Z. First Report on DNA Content of Three Species of Amorphophallus. J. Genet. 2020, 99, 36. [Google Scholar] [CrossRef] [PubMed]
- International Plant Names Index. Available online: https://www.ipni.org/ (accessed on 28 November 2021).
- World Checklist of Selected Plant Families. Available online: https://wcsp.science.kew.org/home.do (accessed on 28 November 2021).
- Bennett, M.D. Nuclear DNA Content and Minimum Generation Time in Herbaceous Plants. Proc. R. Soc. Lond. B Biol. Sci. 1972, 181, 109–135. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, P.; Mukherjee, S.; Sharma, A.K. Cytophotometric Estimation of in Situ DNA Content in Several Species of Araceae. Cytobios 2001, 105, 177–183. [Google Scholar] [PubMed]
- Wang, W.; Kerstetter, R.A.; Michael, T.P. Evolution of Genome Size in Duckweeds (Lemnaceae). J. Bot. 2011, 2011, 570319:1–570319:9. [Google Scholar] [CrossRef]
- Romano, L.E.; Aronne, G. The World Smallest Plants (Wolffia Sp.) as Potential Species for Bioregenerative Life Support Systems in Space. Plants 2021, 10, 1896:1–1896:11. [Google Scholar] [CrossRef]
- Sree, K.S.; Sudakaran, S.; Appenroth, K.-J. How Fast Can Angiosperms Grow? Species and Clonal Diversity of Growth Rates in the Genus Wolffia (Lemnaceae). Acta Physiol. Plant. 2015, 37, 204:1–204:7. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An Update of the Angiosperm Phylogeny Group Classification for the Orders and Families of Flowering Plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Faizullah, L.; Morton, J.A.; Hersch-Green, E.I.; Walczyk, A.M.; Leitch, A.R.; Leitch, I.J. Exploring Environmental Selection on Genome Size in Angiosperms. Trends Plant Sci. 2021, 26, 1039–1049. [Google Scholar] [CrossRef]
- Croat, T.B. A Revision of the Genus Anthurium (Araceae) of Mexico and Central America. Part I: Mexico and Middle America. Ann. Mo. Bot. Gard. 1983, 70, 211–416. [Google Scholar] [CrossRef]
- Croat, T.B. A Revision of the Genus Anthurium (Araceae) of Mexico and Central America. Part II. Panama. Monogr. Syst. Bot. Mo. Bot. Gard. 1986, 14, 204. [Google Scholar] [CrossRef]
- Croat, T.B. A Revision of Philodendron Subgenus Philodendron (Araceae) for Mexico and Central America. Ann. Mo. Bot. Gard. 1997, 84, 311–704. [Google Scholar] [CrossRef]
- Grayum, M.H. Revision of Philodendron Subgenus Pteromischum (Araceae) for Pacific and Caribbean Tropical America. Syst. Bot. Monogr. 1996, 47, 1–233. [Google Scholar] [CrossRef]
- Grayum, M.H. Araceae. In Manual de Plantas de Costa Rica. Vol. II. Gimnospermas y Monocotiledoneas. Agavaceae-Musaceae; Hammel, B.E., Grayum, M.H., Herrera, C., Zamora, N., Eds.; Missouri Botanical Garden Press: St. Louis, MO, USA, 2003; Volume 2, pp. 59–200. ISBN 1-930723-22-9. [Google Scholar]
- Cedeño-Fonseca, M.; Hay, A.; Grayum, M.H.; Blanco, M.A. Two New Endemic Species of Monstera (Araceae: Monsteroideae: Monstereae) from Golfito in Southern Costa Rica. Webbia 2020, 75, 123–132. [Google Scholar] [CrossRef]
- Weissenhofer, A.; Grayum, M.H. Araceae. In An Introductory Field Guide to the Flowering Plants of the Golfo Dulce Rain Forests, Costa Rica; Weber, A., Huber, W., Weissenhofer, A., Zamora, N., Zimmermann, G., Eds.; Catalogue of Oberösterreichisches Landesmuseum; Biologiezentrum des OÖ Landesmuseums: Linz, Austria, 2001; pp. 89–100. ISBN 3-85474-072-7. [Google Scholar]
- Bačič, T.; Frajman, B.; Dolenc Koce, J. Diversification of Luzula Sect. Luzula (Juncaceae) on the Balkan Peninsula—A Cytogenetic Approach. Folia Geobot. 2016, 51, 51–63. [Google Scholar] [CrossRef]
- Vilhar, B.; Greilhuber, J.; Dolenc Koce, J.; Temsch, E.M.; Dermastia, M. Plant Genome Size Measurement with DNA Image Cytometry. Ann. Bot. 2001, 87, 719–728. [Google Scholar] [CrossRef][Green Version]
- Greilhuber, J.; Ebert, I. Genome Size Variation in Pisum Sativum. Genome 1994, 37, 646–655. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Nauheimer, L.; Metzler, D.; Renner, S.S. Global History of the Ancient Monocot Family Araceae Inferred with Models Accounting for Past Continental Positions and Previous Ranges Based on Fossils. New Phytol. 2012, 195, 938–950. [Google Scholar] [CrossRef]
- Vasconcelos, S.; de Lourdes Soares, M.; Sakuragui, C.M.; Croat, T.B.; Oliveira, G.; Benko-Iseppon, A.M. New Insights on the Phylogenetic Relationships among the Traditional Philodendron Subgenera and the Other Groups of the Homalomena Clade (Araceae). Mol. Phylogenet. Evol. 2018, 127, 168–178. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Species | Life Form | Tissue Used for C-Value Measurement | N | 2C ± SE [pg DNA] | CV [%] |
|---|---|---|---|---|---|---|
| Adelonema | A. erythropus | Terrestrial | Leaf, unknown | 9 | 3.50 ± 0.12 | 10.7 |
| A. wendlandiia | Terrestrial | Meristem tissue, unknown, inflorescence, stem (cap) | 12 | 3.51 ± 0.08 | 7.8 | |
| Aglaonema | A. cf. marantifolium | Terrestrial | Unknown | 2 | 62.48 ± 2.70 | 6.1 |
| Alocasia | A. longilobab | Terrestrial | Leaf, stem | 3 | 27.50 ± 0.70 | 4.4 |
| A. clypeolata | Terrestrial | Young leaves, inflorescence, inflorescence (seed embryo) | 7 | 11.02 ± 0.30 | 6.7 | |
| Anthurium | A. brownii | Obligatory epiphyte | Stem | 8 | 8.45 ± 0.10 | 4.3 |
| A. clavigerumb | Obligatory epiphyte | Unknown, stem | 16 | 13.23 ± 0.15 | 4.7 | |
| A. hacumense | Obligatory epiphyte | Roots, inflorescence | 11 | 8.09 ± 0.20 | 8.2 | |
| A. hoffmaniib | Facultative epiphyte | Unknown | 9 | 7.83 ± 0.20 | 7.1 | |
| A. obtusumb | Obligatory epiphyte | Unknown, inflorescence, inflorescence (seed embryo) | 9 | 5.54 ± 0.10 | 7.7 | |
| A. ochranthum b | Terrestrial | Roots, leaf, stem, stem (cap) | 6 | 12.48 ± 0.30 | 5.3 | |
| A. cf. ravenii | Obligatory epiphyte | Unknown | 4 | 8.63 ± 0.20 | 3.5 | |
| Dieffenbachia | D. concinna | Terrestrial | Stem (bud) | 3 | 33.13 ± 0.30 | 1.7 |
| Dracontium | D. pittieri | Terrestrial | Unknown | 2 | 10.17 ± 0.10 | 1.7 |
| Monstera | M. adansonii | Nomadic vine | Stem | 3 | 10.54 ± 0.46 | 9.1 |
| M. gambensis | Nomadic vine | Stem (bud) | 3 | 9.21 ± 0.20 | 3.6 | |
| M. pinnatipartita | Nomadic vine | Leaf, unknown, stem (cap) | 8 | 12.04 ± 0.30 | 6.6 | |
| Philodendron | P. auriculatum | Facultative epiphyte | Unknown, inflorescence, stem | 10 | 3.23 ± 0.10 | 8.4 |
| P. fragrantissimum | Nomadic vine | Leaf, stem | 10 | 4.86 ± 0.10 | 7.7 | |
| P. grandipes | Terrestrial | Inflorescence, inflorescence (seed embryo), stem | 6 | 4.30 ± 0.10 | 4.8 | |
| P. mexicanum | Nomadic vine | Leaf, unknown | 9 | 3.65 ± 0.20 | 15.1 | |
| P. microstictum | Nomadic vine | Inflorescence | 8 | 3.56 ± 0.00 | 3.4 | |
| P. opacum | Nomadic vine | Leaf, inflorescence (seed embryo) | 7 | 3.07 ± 0.10 | 4.3 | |
| P. platypetiolatum | Nomadic vine | Leaf, stem | 9 | 3.55 ± 0.10 | 6.4 | |
| P. popenoei | Terrestrial | Unknown, stem | 6 | 2.90 ± 0.00 | 2.7 | |
| P. pterotum | Nomadic vine | Unknown, stem (cap, bud) | 7 | 5.12 ± 0.10 | 6.7 | |
| P. sagittifolium | Nomadic vine | Leaf (leaf embryo), stem, stem (cap) | 9 | 3.60 ± 0.00 | 3.5 | |
| P. rhodoaxis | Nomadic vine | Stem, root | 6 | 2.54 ± 0.10 | 12.8 | |
| P. sp. | Facultative epiphyte | Leaf, root | 3 | 3.28 ± 0.10 | 7.8 | |
| P. tripartitum | Nomadic vine | Leaf, stem | 7 | 3.78 ± 0.10 | 5.5 | |
| Pistia | P. stratiotesb | Aquatic | Leaf, stem, stem (cap) | 15 | 0.83 ± 0.00 | 8.7 |
| Rhodospatha | R. osaensis | Nomadic vine | Root, leaf, stem | 11 | 3.09 ± 0.27 | 28.9 |
| R. cf. osaensis | Nomadic vine | Leaf (leaf embryo) | 4 | 2.54 ± 0.00 | 3.3 | |
| Spathiphyllum | S. cf. leave | Terrestrial | Leaf | 5 | 22.12 ± 0.30 | 2.6 |
| S. silvicola | Terrestrial | Leaf, inflorescence | 6 | 18.29 ± 0.50 | 6.7 | |
| S. wendlandii | Terrestrial | Leaf, stem (node) | 6 | 20.07 ± 0.40 | 5.1 | |
| Stenospermation | S. angustifolium | Obligatory epiphyte | Unknown, inflorescence, inflorescence (seed embryo), stem | 9 | 7.49 ± 0.20 | 6.5 |
| S. cf. maranthifolium | Obligatory epiphyte | Inflorescence, stem | 9 | 7.24 ± 0.20 | 9.1 | |
| Syngonium | S. hastiferum | Nomadic vine | Roots, leaf, unknown, stem | 19 | 7.16 ± 0.20 | 9.2 |
| S. podophyllumb | Nomadic vine | Roots, leaf, stem (cap) | 10 | 5.48 ± 0.10 | 5.5 | |
| Xanthosoma | X. sagittifoliumb | Terrestrial | Leaf, stem | 9 | 4.81 ± 0.10 | 6.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocjan, D.; Dolenc Koce, J.; Etl, F.; Dermastia, M. Genome Size of Life Forms of Araceae—A New Piece in the C-Value Puzzle. Plants 2022, 11, 334. https://doi.org/10.3390/plants11030334
Kocjan D, Dolenc Koce J, Etl F, Dermastia M. Genome Size of Life Forms of Araceae—A New Piece in the C-Value Puzzle. Plants. 2022; 11(3):334. https://doi.org/10.3390/plants11030334
Chicago/Turabian StyleKocjan, Domen, Jasna Dolenc Koce, Florian Etl, and Marina Dermastia. 2022. "Genome Size of Life Forms of Araceae—A New Piece in the C-Value Puzzle" Plants 11, no. 3: 334. https://doi.org/10.3390/plants11030334
APA StyleKocjan, D., Dolenc Koce, J., Etl, F., & Dermastia, M. (2022). Genome Size of Life Forms of Araceae—A New Piece in the C-Value Puzzle. Plants, 11(3), 334. https://doi.org/10.3390/plants11030334