

Exogenous Phytohormones and Fertilizers Enhance Jatropha curcas L. Growth through the Regulation of Physiological, Morphological, and Biochemical Parameters

 ,
,  ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Phytohormones and Fertilizer Enhances J. curcas Growth Vigor
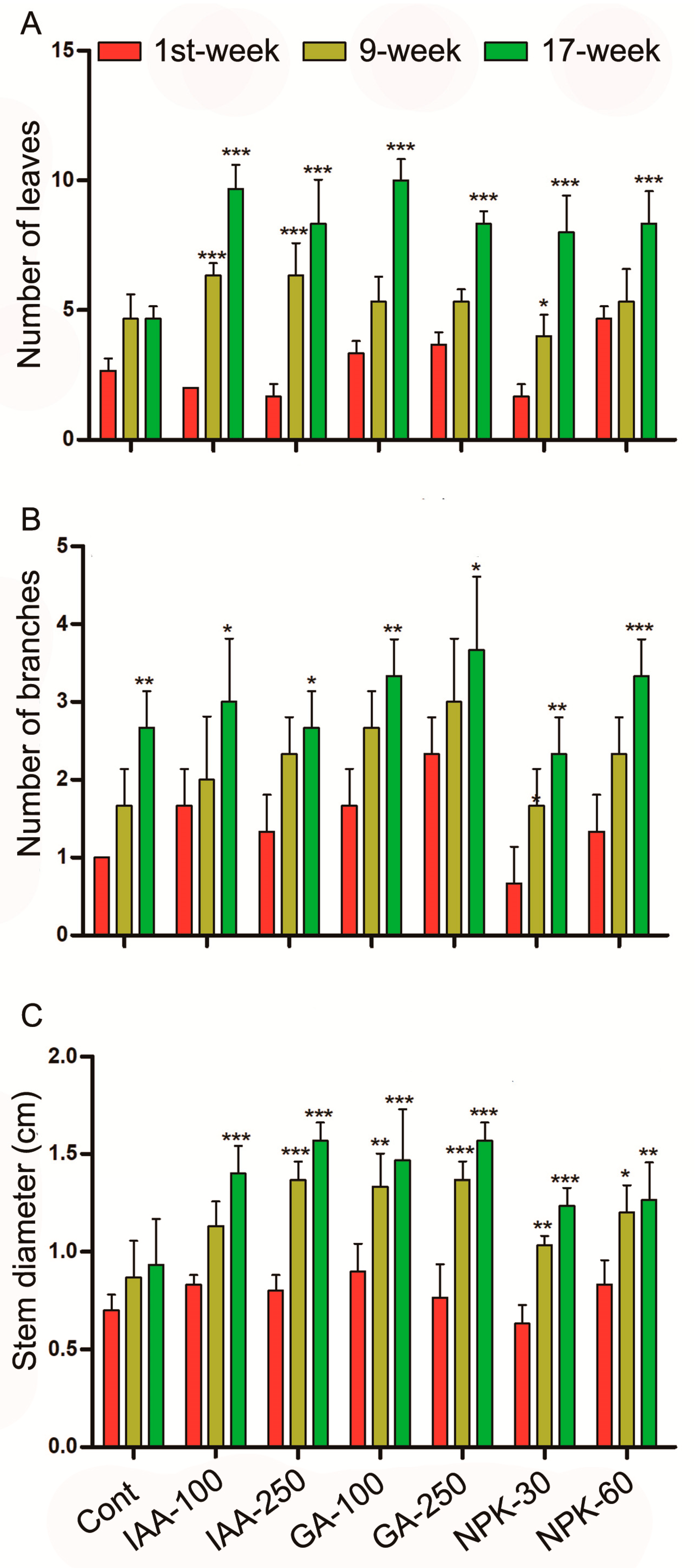
2.2. Phytohormones and Fertilizer Treatment Promotes Leaf and Branch Number and Stem Diameter
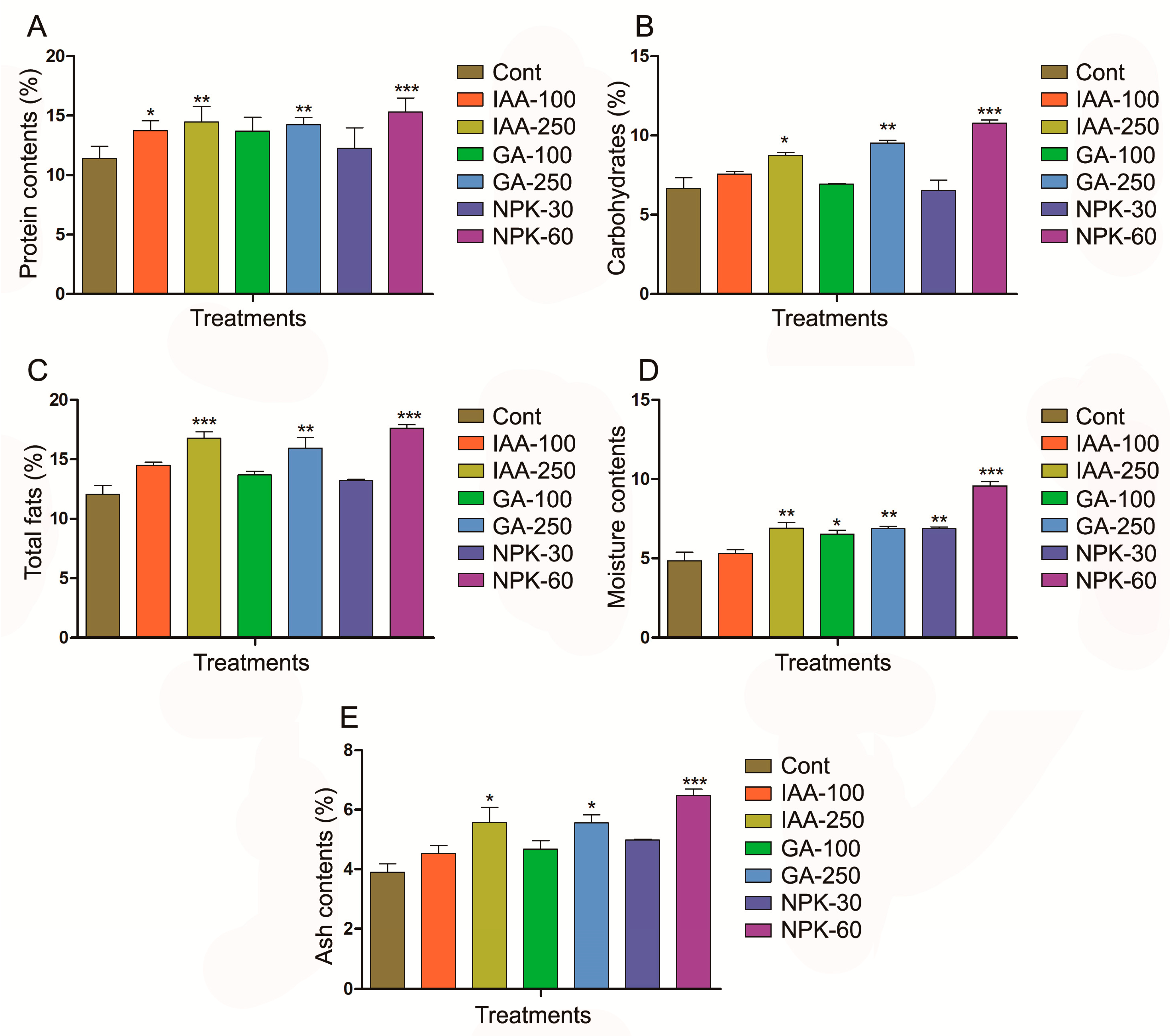
2.3. Phytohormones and Fertilizers Promote J. curcas Plant Growth by Regulating Proximate Compositions
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Material Used
4.2. Parameters Studied
4.3. Proximate Composition Analysis
4.4. Soil Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lustosa Sobrinho, R.; Zoz, T.; Finato, T.; Oliveira, C.E.d.S.; Neto, S.S.d.O.; Zoz, A.; Alaraidh, I.A.; Okla, M.K.; Alwasel, Y.A.; Beemster, G. Jatropha curcas L. as a Plant Model for Studies on Vegetative Propagation of Native Forest Plants. Plants 2022, 11, 2457. [Google Scholar] [CrossRef] [PubMed]
- Holl, M.; Gush, M.; Hallowes, J.; Versfeld, D. Jatropha curcas in South Africa: An Assessment of Its Water Use and Bio-Physical Potential; Pretoria South Africa Water Research Commission: Pretoria, South Africa, 2007.
- Neupane, D.; Bhattarai, D.; Ahmed, Z.; Das, B.; Pandey, S.; Solomon, J.K.; Qin, R.; Adhikari, P. Growing Jatropha (Jatropha curcas L.) as a potential second-generation biodiesel feedstock. Inventions 2021, 6, 60. [Google Scholar] [CrossRef]
- Salimon, J.; Abdullah, R. Physicochemical properties of Malaysian Jatropha curcas seed oil. Sains Malays. 2008, 37, 379–382. [Google Scholar]
- Abou Kheira, A.A.; Atta, N.M. Response of Jatropha curcas L. to water deficits: Yield, water use efficiency and oilseed characteristics. Biomass Bioenergy 2009, 33, 1343–1350. [Google Scholar] [CrossRef]
- Kumar, G.; Yadav, S.; Thawale, P.; Singh, S.; Juwarkar, A. Growth of Jatropha curcas on heavy metal contaminated soil amended with industrial wastes and Azotobacter–A greenhouse study. Bioresour. Technol. 2008, 99, 2078–2082. [Google Scholar] [CrossRef]
- Achten, W.M.; Verchot, L.; Franken, Y.J.; Mathijs, E.; Singh, V.P.; Aerts, R.; Muys, B. Jatropha bio-diesel production and use. Biomass Bioenergy 2008, 32, 1063–1084. [Google Scholar] [CrossRef]
- Joshi, G.; Shukla, A.; Shukla, A. Synergistic response of auxin and ethylene on physiology of Jatropha curcas L. Braz. J. Plant Physiol. 2011, 23, 66–77. [Google Scholar] [CrossRef]
- Srivastava, L.M. Plant Growth and Development: Hormones and Environment; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Leite, V.M.; Rosolem, C.A.; Rodrigues, J.D. Gibberellin and cytokinin effects on soybean growth. Sci. Agric. 2003, 60, 537–541. [Google Scholar] [CrossRef]
- Wang, Y.; Mopper, S.; Hasenstein, K.H. Effects of salinity on endogenous ABA, IAA, JA, and SA in Iris hexagona. J. Chem. Ecol. 2001, 27, 327–342. [Google Scholar] [CrossRef]
- Akbarian, M.; Modafebehzadi, N.; Bagheripour, M. Study of fertilizer (NPK) effects on yield and triglycerids in Jatropha (Jatropha curcas). Plant Ecophysiol. 2010, 2, 169–172. [Google Scholar]
- Sapeta, H.; Costa, J.M.; Lourenco, T.; Maroco, J.; van der Linde, P.; Oliveira, M.M. Drought stress response in Jatropha curcas: Growth and physiology. Environ. Exp. Bot. 2013, 85, 76–84. [Google Scholar] [CrossRef]
- Zolman, B.K.; Yoder, A.; Bartel, B. Genetic analysis of indole-3-butyric acid responses in Arabidopsis thaliana reveals four mutant classes. Genetics 2000, 156, 1323–1337. [Google Scholar] [CrossRef]
- El-Eslamboly, A.A. Effect of watermelon propagation by cuttings on vegetative growth, yield and fruit quality. Egypt. J. Agric. Res. 2014, 92, 553–579. [Google Scholar] [CrossRef]
- Sadak, M.S.; Dawood, M.G.; Bakry, B.; El-Karamany, M. Synergistic effect of indole acetic acid and kinetin on performance, some biochemical constituents and yield of faba bean plant grown under newly reclaimed sandy soil. World J. Agric. Sci. 2013, 9, 335–344. [Google Scholar]
- Ogwu, M.C.; Osas, A.R.; Edwin, O.M. Effects of Indole-3-Acetic Acid on Germination in Lead Polluted Petri Dish of Citrullus lanatus (Thunberg) Matsumura and Nakai, Cucurbitaceae. Aceh Int. J. Sci. Technol. 2015, 4, 107–113. [Google Scholar] [CrossRef]
- Zhao, T.; Deng, X.; Xiao, Q.; Han, Y.; Zhu, S.; Chen, J. IAA priming improves the germination and seedling growth in cotton (Gossypium hirsutum L.) via regulating the endogenous phytohormones and enhancing the sucrose metabolism. Ind. Crops Prod. 2020, 155, 112788. [Google Scholar] [CrossRef]
- Farooq, M.; Jan, R.; Kim, K.-M. Gravistimulation effects on Oryza sativa amino acid profile, growth pattern and expression of OsPIN genes. Sci. Rep. 2020, 10, 17303. [Google Scholar] [CrossRef] [PubMed]
- Goren, S.; Lugassi, N.; Stein, O.; Yeselson, Y.; Schaffer, A.A.; David-Schwartz, R.; Granot, D. Suppression of sucrose synthase affects auxin signaling and leaf morphology in tomato. PLoS ONE 2017, 12, e0182334. [Google Scholar] [CrossRef]
- Martens, D.; Luck, S.; Frankenberger, W., Jr. Role of plant growth regulators in vegetative spring flush, flowering, and fruit drop in avocado (Persea americana, Mill.). Circular 1994, CAS-94, 1–14. [Google Scholar]
- Sarkar, P.K.; Haque, M.S.; Abdul Karim, M. Effects of GA and IAA and their frequency of application on morphology, yield 3. Pak. J. Agron. 2002, 1, 119–122. [Google Scholar]
- El-Bassiouny, H.M.; Shukry, W.M. Cowpea growth pattern, metabolism and yield in response to IAA and biofertilizers under drought conditions. Egypt. J. Biol. 2001, 3, 117–129. [Google Scholar]
- Zhang, Y.; Li, Y.; Hassan, M.J.; Li, Z.; Peng, Y. Indole-3-acetic acid improves drought tolerance of white clover via activating auxin, abscisic acid and jasmonic acid related genes and inhibiting senescence genes. BMC Plant Biol. 2020, 20, 150. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Welti, R.; Wang, X. Quantitative analysis of major plant hormones in crude plant extracts by high-performance liquid chromatography–mass spectrometry. Nat. Protoc. 2010, 5, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, G.G.; Alhamd, M. Effect of gibberellic acid and indole 3-acetic acid on improving growth and accumulation of phytochemical composition in Balanites aegyptiaca plants. Am. J. Plant Physiol. 2011, 6, 36–43. [Google Scholar] [CrossRef][Green Version]
- Hu, Y.-X.; Tao, Y.-B.; Xu, Z.-F. Overexpression of Jatropha Gibberellin 2-oxidase 6 (JcGA2ox6) induces dwarfism and smaller leaves, flowers and fruits in Arabidopsis and Jatropha. Front. Plant Sci. 2017, 8, 2103. [Google Scholar] [CrossRef]
- Miceli, A.; Moncada, A.; Sabatino, L.; Vetrano, F. Effect of gibberellic acid on growth, yield, and quality of leaf lettuce and rocket grown in a floating system. Agronomy 2019, 9, 382. [Google Scholar] [CrossRef]
- Khan, N. Effect of gibberellic acid on carbonic anhydrase, photosynthesis, growth and yield of mustard. Biol. Plant. 1996, 38, 145–147. [Google Scholar] [CrossRef]
- Khan, N. Variation in carbonic anhydrase activity and its relationship with photosynthesis and dry mass of mustard. Photosynthetica 1994, 30, 317. [Google Scholar]
- Devies, P.; Sun, T. Gibberellin signal transduction in stem elongation and leaf growth. In Plant Hormones; Springer: Dordrecht, The Netherlands, 2004. [Google Scholar]
- Basra, A. Plant Growth Regulators in Agriculture and Horticulture: Their Role and Commercial Uses; CRC Press: Boca Raton, FL, USA, 2000. [Google Scholar]
- Aloni, R. Role of auxin and gibberellin in differentiation of primary phloem fibers. Plant Physiol. 1979, 63, 609–614. [Google Scholar] [CrossRef]
- Ullah, S.; Anwar, S.; Rehman, M.; Khan, S.; Zafar, S.; Liu, L.; Peng, D. Interactive effect of gibberellic acid and NPK fertilizer combinations on ramie yield and bast fibre quality. Sci. Rep. 2017, 7, 10647. [Google Scholar] [CrossRef]
- Ni, J.; Gao, C.; Chen, M.-S.; Pan, B.-Z.; Ye, K.; Xu, Z.-F. Gibberellin promotes shoot branching in the perennial woody plant Jatropha curcas. Plant Cell Physiol. 2015, 56, 1655–1666. [Google Scholar] [CrossRef] [PubMed]
- Koorneef, M.; Elgersma, A.; Hanhart, C.v.; van Loenen-Martinet, E.; Van Rijn, L.; Zeevaart, J. A gibberellin insensitive mutant of Arabidopsis thaliana. Physiol. Plant. 1985, 65, 33–39. [Google Scholar] [CrossRef]
- Scott, T.K.; Case, D.B.; Jacobs, W.P. Auxin-gibberellin interaction in apical dominance. Plant Physiol. 1967, 42, 1329–1333. [Google Scholar] [CrossRef] [PubMed]
- Agharkar, M.; Lomba, P.; Altpeter, F.; Zhang, H.; Kenworthy, K.; Lange, T. Stable expression of AtGA2ox1 in a low-input turfgrass (Paspalum notatum Flugge) reduces bioactive gibberellin levels and improves turf quality under field conditions. Plant Biotechnol. J. 2007, 5, 791–801. [Google Scholar] [CrossRef] [PubMed]
- Mauriat, M.; Sandberg, L.G.; Moritz, T. Proper gibberellin localization in vascular tissue is required to control auxin-dependent leaf development and bud outgrowth in hybrid aspen. Plant J. 2011, 67, 805–816. [Google Scholar] [CrossRef]
- Zawaski, C.; Busov, V.B. Roles of gibberellin catabolism and signaling in growth and physiological response to drought and short-day photoperiods in Populus trees. PLoS ONE 2014, 9, e86217. [Google Scholar] [CrossRef]
- Marth, P.C.; Audia, W.V.; Mitchell, J.W. Effects of gibberellic acid on growth and development of plants of various genera and species. Bot. Gaz. 1956, 118, 106–111. [Google Scholar] [CrossRef]
- Elfving, D.C.; Visser, D.B.; Henry, J.L. Gibberellins stimulate lateral branch development in young sweet cherry trees in the orchard. Int. J. Fruit Sci. 2011, 11, 41–54. [Google Scholar] [CrossRef]
- Choubane, D.; Rabot, A.; Mortreau, E.; Legourrierec, J.; Péron, T.; Foucher, F.; Ahcène, Y.; Pelleschi-Travier, S.; Leduc, N.; Hamama, L. Photocontrol of bud burst involves gibberellin biosynthesis in Rosa sp. J. Plant Physiol. 2012, 169, 1271–1280. [Google Scholar] [CrossRef]
- Björklund, S.; Antti, H.; Uddestrand, I.; Moritz, T.; Sundberg, B. Cross-talk between gibberellin and auxin in development of Populus wood: Gibberellin stimulates polar auxin transport and has a common transcriptome with auxin. Plant J. 2007, 52, 499–511. [Google Scholar] [CrossRef]
- Mauriat, M.; Moritz, T. Analyses of GA20ox-and GID1-over-expressing aspen suggest that gibberellins play two distinct roles in wood formation. Plant J. 2009, 58, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Denis, E.; Kbiri, N.; Mary, V.; Claisse, G.; Conde e Silva, N.; Kreis, M.; Deveaux, Y. WOX 14 promotes bioactive gibberellin synthesis and vascular cell differentiation in Arabidopsis. Plant J. 2017, 90, 560–572. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Krishnan, S.; Merewitz, E.; Xu, J.; Huang, B. Gibberellin-regulation and genetic variations in leaf elongation for tall fescue in association with differential gene expression controlling cell expansion. Sci. Rep. 2016, 6, 30258. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Hussain, I.; Nabi, G. Exogenous gibberellic acid application influences on vegetative and reproductive aspects in gladiolus. Ornam. Hortic. 2020, 26, 244–250. [Google Scholar] [CrossRef]
- Chen, W.-S.; Liu, H.-Y.; Liu, Z.-H.; Yang, L.; Chen, W.-H. Gibberellin and temperature influence carbohydrate content and flowering in Phalaenopsis [sucrose feeding]. Physiol. Plant. 1994, 90, 391–395. [Google Scholar] [CrossRef]
- Crozier, A.; Turnbull, C. Gibberellins: Biochemistry and action in extension growth. What New Plant Physiol. 1984, 15, 9–12. [Google Scholar]
- Wood, A.; Paleg, L. Alteration of liposomal membrane fluidity by gibberellic acid. Funct. Plant Biol. 1974, 1, 31–40. [Google Scholar] [CrossRef]
- Ansari, H. Effect of Some Phytohormones and NPK on Growth and Metabolism of Mustard. Ph.D. Thesis, Aligarh Muslim University, Aligarh, India, 1996. [Google Scholar]
- Estruch, J.J.; Peretó, J.G.; Vercher, Y.; Beltrán, J.P. Sucrose loading in isolated veins of Pisum sativum: Regulation by abscisic acid, gibberellic acid, and cell turgor. Plant Physiol. 1989, 91, 259–265. [Google Scholar] [CrossRef]
- Yuxi, Z.; Yanchao, Y.; Zejun, L.; Tao, Z.; Feng, L.; Chunying, L.; Shupeng, G. GA3 is superior to GA4 in promoting bud endodormancy release in tree peony (Paeonia suffruticosa) and their potential working mechanism. BMC Plant Biol. 2021, 21, 323. [Google Scholar] [CrossRef]
- Singh, S.; Kulkarni, M.G.; Van Staden, J. Biochemical changes associated with gibberellic acid-like activity of smoke-water, karrikinolide and vermicompost leachate during seedling development of Phaseolus vulgaris L. Seed Sci. Res. 2014, 24, 63–70. [Google Scholar] [CrossRef]
- Chandio, A.S.; Depar, M.S.; Meghwar, B.L.; Waseem, M.; Kalroo, A.K.P.; Solangi, A.H.; Rind, P.A.; Ahmed, N. Nutrient Requirement of Jatropha cultivars of Diversified Origin. Int. J. Biol. Biotechnol. 2016, 13, 385–392. [Google Scholar]
- Franken, Y.; Nielsen, F. Plantation establishment and management. In The Jatropha Handbook-From Cultivation to Application (9–29); FACT Foundation: Eindhoven, The Netherlands, 2010. [Google Scholar]
- Mohapatra, S.; Panda, P.K. Effects of fertilizer application on growth and yield of Jatropha curcas L. in an aeric tropaquept of Eastern India. Not. Sci. Biol. 2011, 3, 95–100. [Google Scholar] [CrossRef]
- Patolia, J.; Ghosh, A.; Chikara, J.; Chaudhary, D.; Parmar, D.; Bhuva, H. Response of Jatropha curcas grown on wasteland to N and P fertilization. In Proceedings of the Expert seminar on Jatropha curcas L. Agronomy and Genetics, Wageningen, The Netherlands, 26–28 March 2007; pp. 26–28. [Google Scholar]
- Duarah, I.; Deka, M.; Saikia, N.; Deka Boruah, H. Phosphate solubilizers enhance NPK fertilizer use efficiency in rice and legume cultivation. 3 Biotech 2011, 1, 227–238. [Google Scholar] [CrossRef]
- Yan, L.; Sunoj, V.J.; Short, A.W.; Lambers, H.; Elsheery, N.I.; Kajita, T.; Wee, A.K.; Cao, K.F. Correlations between allocation to foliar phosphorus fractions and maintenance of photosynthetic integrity in six mangrove populations as affected by chilling. New Phytol. 2021, 232, 2267–2282. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, R.A.; Hussein, O.S.; Abdelkader, A.F.; Farag, I.A.; Hassan, Y.E.; Ibrahim, M. Metabolic activities and molecular investigations of the ameliorative impact of some growth biostimulators on chilling-stressed coriander (Coriandrum sativum L.) plant. BMC Plant Biol. 2021, 21, 361. [Google Scholar] [CrossRef]
- Umami, N.; Abdiyansah, A.; Agus, A. Effects of different doses of NPK fertilization on growth and productivity of Cichorium intybus. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Moscow, Russia, 27 May–6 June 2019; p. 012097. [Google Scholar]
- Goenadi, D. Pupuk dan Teknologi Pemupukan Berbasis Hayati Dari Cawan Petri ke Lahan Petani; Yayasan John Hi-Tech Idetama Jkt.: Jakarta, Indonesia, 2006. [Google Scholar]
- Suprihati, N.D. Soil chemical and mineralogical characteristics and its relationship with the fertilizers requirement for rice (Oriza sativa), maize (Zea mays), and soybean (Glycine max). Bul. Agron 2005, 33, 40–47. [Google Scholar]
- Hillel, D.; Hatfield, J.L. Encyclopedia of Soils in the Environment; Elsevier: Amsterdam, The Netherlands, 2005; Volume 3. [Google Scholar]
- Fixen, P. Nutrient management following conservation reserve program. Better Crops 1996, 80, 16–19. [Google Scholar]
- Lynch, J.; Läuchli, A.; Epstein, E. Vegetative growth of the common bean in response to phosphorus nutrition. Crop Sci. 1991, 31, 380–387. [Google Scholar] [CrossRef]
- Khan, S.; Anwar, S.; Kuai, J.; Ullah, S.; Fahad, S.; Zhou, G. Optimization of nitrogen rate and planting density for improving yield, nitrogen use efficiency, and lodging resistance in oilseed rape. Front. Plant Sci. 2017, 8, 532. [Google Scholar] [CrossRef]
- Huang, C.; Wei, G.; Luo, Z.; Xu, J.; Zhao, S.; Wang, L.; Jie, Y. Effects of nitrogen on ramie (Boehmeria nivea) hybrid and its parents grown under field conditions. J. Agric. Sci. 2014, 6, 230. [Google Scholar] [CrossRef]
- Almazan, A.M.; Adeyeye, S.O. Fat and fatty acid concentrations in some green vegetables. J. Food Compos. Anal. 1998, 11, 375–380. [Google Scholar] [CrossRef]
- Paquot, C.; Hautfenne, A. Standard Methods for the Analysis of Oils, Fats and Derivatives; Blackwell Scientific Publications Oxford: Hoboken, NJ, USA, 1987; Volume 73. [Google Scholar]
- Latimer, G. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 2016. [Google Scholar]
- Mæhre, H.K.; Dalheim, L.; Edvinsen, G.K.; Elvevoll, E.O.; Jensen, I.-J. Protein determination—Method matters. Foods 2018, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Marcel, A.; Bievenu, M.J. Proximate, mineral and phytochemical analysis of the leaves of H. myriantha and Urera trinervis. Pak. J. Biol. Sci. PJBS 2012, 15, 536–541. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample No | pH | Electric Conductivity ds/m | CaCO3 % | Organic Manure % | Nitrogen % | Texture |
|---|---|---|---|---|---|---|
| 1 | 7.51 | 0.55 | 14 | 0.69 | 0.0345 | Clay loam |
| 2 | 7.31 | 0.67 | 16 | 0.759 | 0.0379 | Clay loam |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jan, R.; Khan, M.; Adnan, M.; Asaf, S.; Asif, S.; Kim, K.-M.; Murad, W. Exogenous Phytohormones and Fertilizers Enhance Jatropha curcas L. Growth through the Regulation of Physiological, Morphological, and Biochemical Parameters. Plants 2022, 11, 3584. https://doi.org/10.3390/plants11243584
Jan R, Khan M, Adnan M, Asaf S, Asif S, Kim K-M, Murad W. Exogenous Phytohormones and Fertilizers Enhance Jatropha curcas L. Growth through the Regulation of Physiological, Morphological, and Biochemical Parameters. Plants. 2022; 11(24):3584. https://doi.org/10.3390/plants11243584
Chicago/Turabian StyleJan, Rahmatullah, Murtaza Khan, Muhammad Adnan, Sajjad Asaf, Saleem Asif, Kyung-Min Kim, and Waheed Murad. 2022. "Exogenous Phytohormones and Fertilizers Enhance Jatropha curcas L. Growth through the Regulation of Physiological, Morphological, and Biochemical Parameters" Plants 11, no. 24: 3584. https://doi.org/10.3390/plants11243584
APA StyleJan, R., Khan, M., Adnan, M., Asaf, S., Asif, S., Kim, K.-M., & Murad, W. (2022). Exogenous Phytohormones and Fertilizers Enhance Jatropha curcas L. Growth through the Regulation of Physiological, Morphological, and Biochemical Parameters. Plants, 11(24), 3584. https://doi.org/10.3390/plants11243584