
Severe Shoot Trimming and Crop Size as Tools to Modulate Cv. Merlot Berry Composition

 ,
,  , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Vineyard Site
2.2. Experimental Design
2.3. Leaf Area, Cluster Exposure, and Yield Components
2.4. Berry Sampling and Fruit Composition
2.5. Statistical Analysis
3. Results
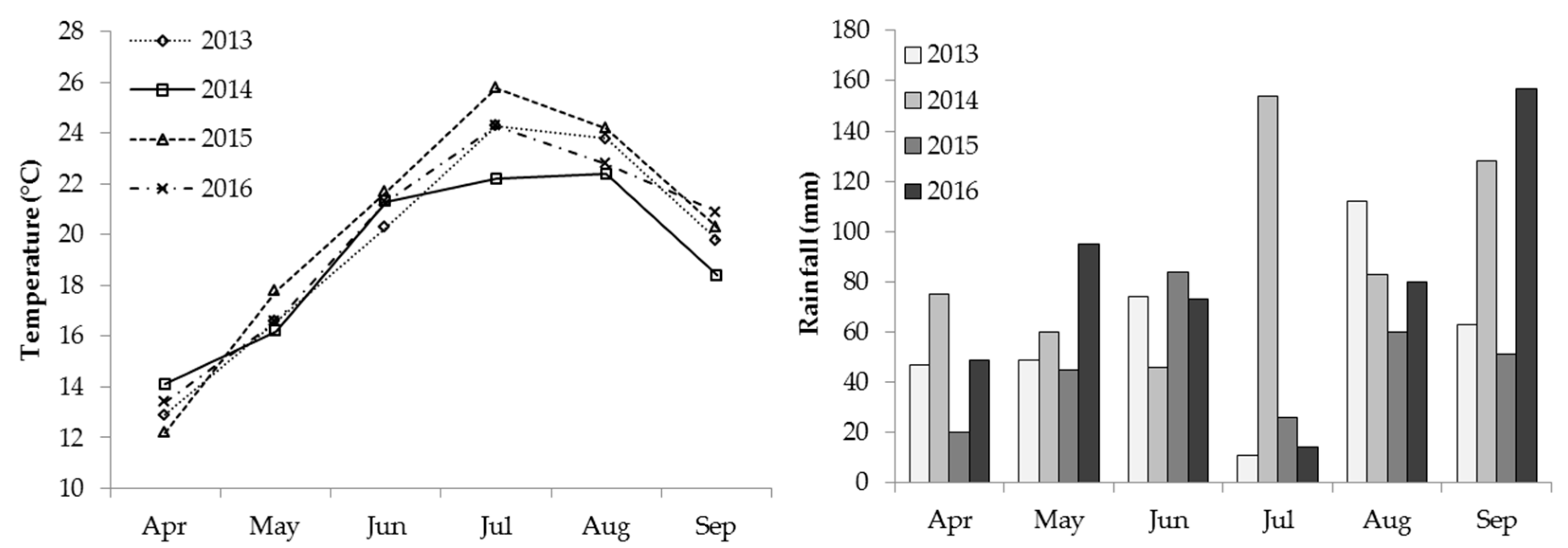
3.1. Meteorological Conditions
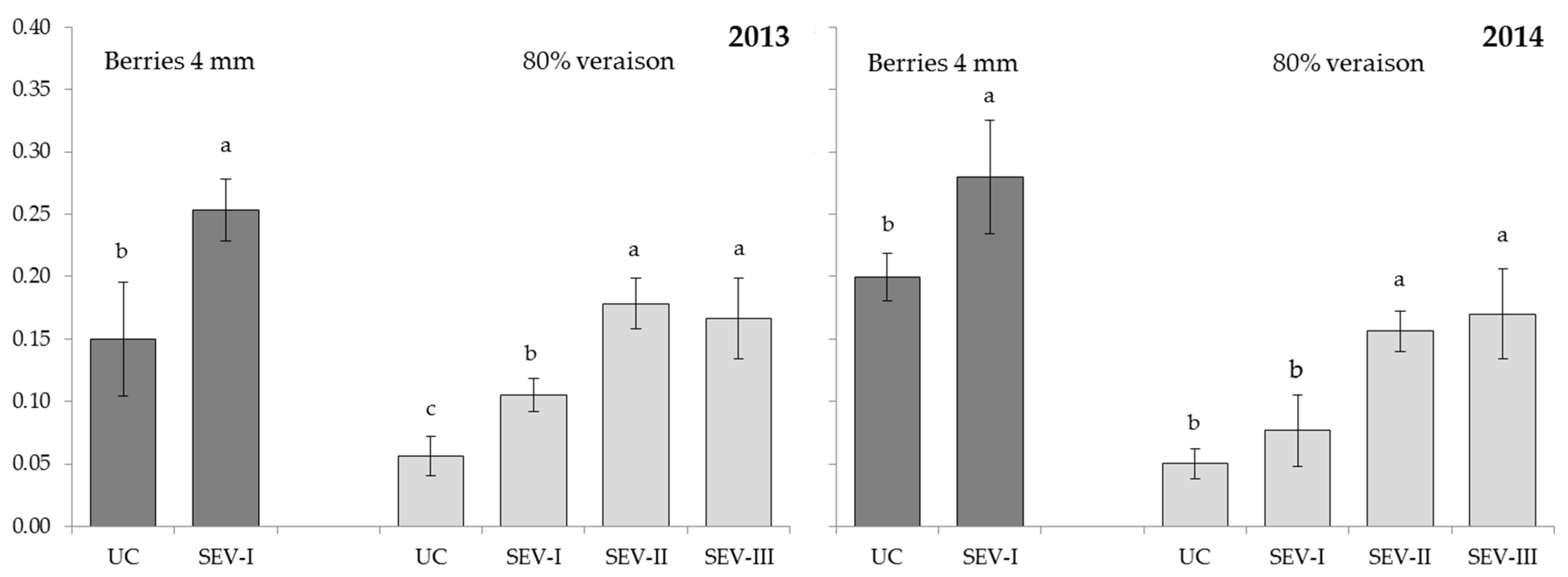
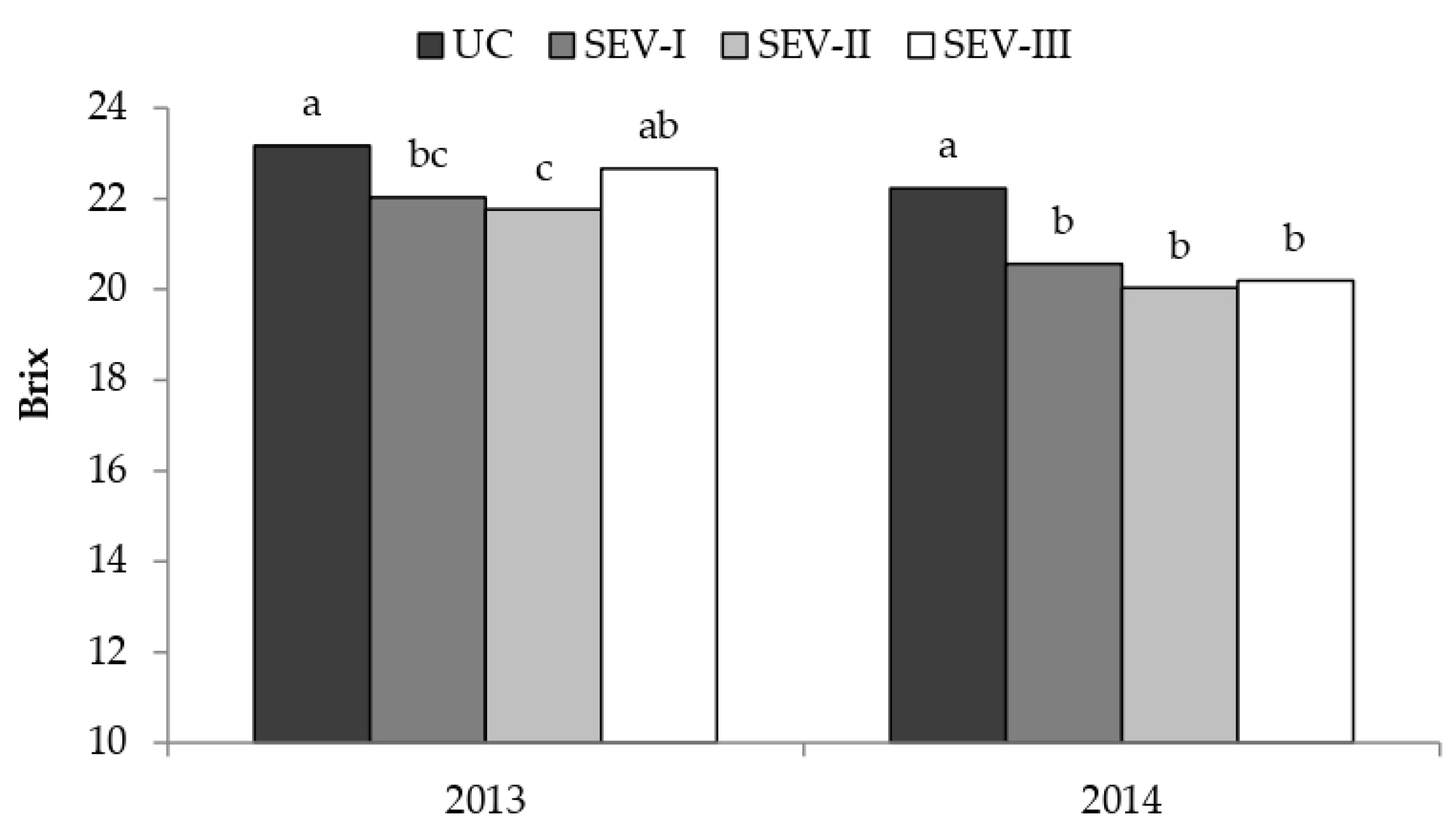
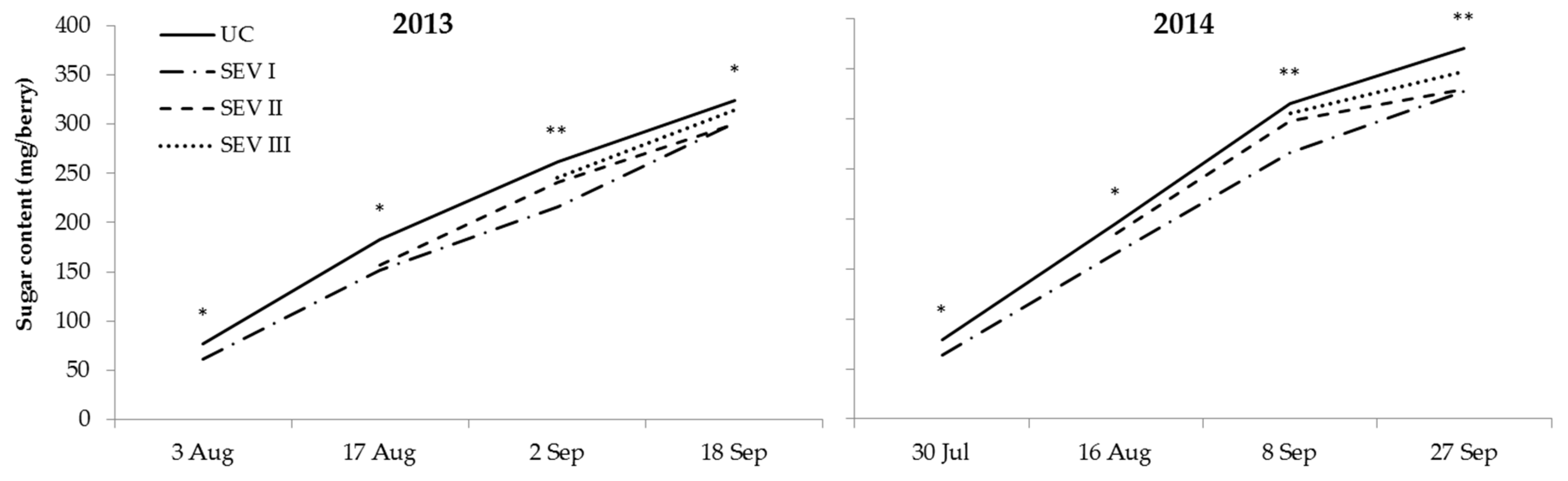
3.2. Study of Severe Shoot Trimming at Different Phenological Stages
3.3. Study of Severe Shoot Trimming Combined with Crop Size
4. Discussion
4.1. Study of Severe Shoot Trimming at Different Phenological Stages
4.2. Study of Severe Shoot Trimming Combined with Crop Size
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Van Leeuwen, C.; Destrac-Irvine, A.; Dubernet, M.; Duchêne, E.; Gowdy, M.; Marguerit, E.; Pieri, P.; Parker, A.; De Resseguier, L.; Ollat, N. An update on the impact of climate change in viticulture and potential adaptations. Agronomy 2019, 9, 514. [Google Scholar] [CrossRef]
- Gambetta, G.; Kurtural, S.K. Global warming and wine quality: Are we close to the tipping point? OENO One 2021, 55, 353–361. [Google Scholar] [CrossRef]
- Cameron, W.; Petrie, P.R.; Barlow, E.; Howell, K.; Jarvis, C.; Fuentes, S. A comparison of the effect of temperature on grapevine phenology between vineyards. OENO One 2021, 55, 301–320. [Google Scholar] [CrossRef]
- Bucher, T.; Deroover, K.; Stockley, C. Low-alcohol wine: A narrative review on consumer perception and behaviour. Beverages 2018, 4, 82. [Google Scholar] [CrossRef]
- Gouot, J.C.; Smith, J.P.; Holzapfel, B.P.; Walker, A.R.; Barril, C. Grape berry flavonoids: A review of their biochemical responses to high and extreme high temperatures. J. Exp. Bot. 2019, 70, 397–423. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Zheng, W.; de Toda, F.M. Current viticultural techniques to mitigate the effects of global warming on grape and wine quality: A comprehensive review. Food Res. Int. 2021, 139, 109946. [Google Scholar] [CrossRef]
- Peterlunger, E.; Sivilotti, P.; Colussi, V. Water stress increased polyphenolic quality in Merlot grapes. In Proceedings of the VII International Symposium on Grapevine Physiology and Biotechnology 689, Davis, CA, USA, 21–25 June 2004; pp. 293–300. [Google Scholar] [CrossRef]
- Guilpart, N.; Metay, A.; Gary, C. Grapevine bud fertility and number of berries per bunch are determined by water and nitrogen stress around flowering in the previous year. Eur. J. Agron. 2014, 54, 9–20. [Google Scholar] [CrossRef]
- Fraga, H.; Molitor, D.; Leolini, L.; Santos, J.A. What is the impact of heatwaves on European viticulture? A modelling assessment. Appl. Sci. 2020, 10, 3030. [Google Scholar] [CrossRef]
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.-T.; Correia, C.; Moriondo, M.; Leolini, L.; Dibari, C.; Costafreda-Aumedes, S. A review of the potential climate change impacts and adaptation options for European viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Cataldo, E.; Fucile, M.; Mattii, G.B. A Review: Soil management, sustainable strategies and approaches to improve the quality of modern viticulture. Agronomy 2021, 11, 2359. [Google Scholar] [CrossRef]
- Marín, D.; Armengol, J.; Carbonell-Bejerano, P.; Escalona, J.; Gramaje, D.; Hernández-Montes, E.; Intrigliolo, D.S.; Martínez-Zapater, J.M.; Medrano, H.; Mirás-Avalos, J.M. Challenges of viticulture adaptation to global change: Tackling the issue from the roots. Aust. J. Grape Wine Res. 2021, 27, 8–25. [Google Scholar] [CrossRef]
- Neethling, E.; Barbeau, G.; Coulon-Leroy, C.; Quénol, H. Spatial complexity and temporal dynamics in viticulture: A review of climate-driven scales. Agric. For. Meteorol. 2019, 276, 107618. [Google Scholar] [CrossRef]
- Bubola, M.; Sivilotti, P.; Janjanin, D.; Poni, S. Early leaf removal has a larger effect than cluster thinning on grape phenolic composition in cv. Teran. Am. J. Enol. Vitic. 2017, 68, 234–242. [Google Scholar] [CrossRef]
- Bubola, M.; Lukić, I.; Radeka, S.; Sivilotti, P.; Grozić, K.; Vanzo, A.; Bavčar, D.; Lisjak, K. Enhancement of Istrian Malvasia wine aroma and hydroxycinnamate composition by hand and mechanical leaf removal. J. Sci. Food Agric. 2019, 99, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Mucalo, A.; Budić-Leto, I.; Lukšić, K.; Maletić, E.; Zdunić, G. Early Defoliation Techniques Enhance Yield Components, Grape and Wine Composition of cv. Trnjak (Vitis vinifera L.) in Dalmatian Hinterland Wine Region. Plants 2021, 10, 551. [Google Scholar] [CrossRef] [PubMed]
- Iorio, D.; Gambacorta, G.; Tarricone, L.; Vilanova, M.; Paradiso, V.M. Effect of Early Basal Leaf Removal on Phenolic and Volatile Composition and Sensory Properties of Aglianico Red Wines. Plants 2022, 11, 591. [Google Scholar] [CrossRef]
- Poni, S.; Casalini, L.; Bernizzoni, F.; Civardi, S.; Intrieri, C. Effects of early defoliation on shoot photosynthesis, yield components, and grape composition. Am. J. Enol. Vitic. 2006, 57, 397–407. [Google Scholar] [CrossRef]
- Poni, S.; Zamboni, M.; Vercesi, A.; Garavani, A.; Gatti, M. Effects of early shoot trimming of varying severity on single high-wire trellised pinot noir grapevines. Am. J. Enol. Vitic. 2014, 65, 493–498. [Google Scholar] [CrossRef]
- Martínez de Toda, F.; Sancha, J.C.; Zheng, W.; Balda, P. Leaf area reduction by trimming, a growing technique to restore the anthocyanins: Sugars ratio decoupled by the warming climate. Vitis 2014, 53, 189–192. [Google Scholar] [CrossRef]
- Filippetti, I.; Movahed, N.; Allegro, G.; Valentini, G.; Pastore, C.; Colucci, E.; Intrieri, C. Effect of post-veraison source limitation on the accumulation of sugar, anthocyanins and seed tannins in Vitis vinifera cv. Sangiovese berries. Aust. J. Grape Wine Res. 2015, 21, 90–100. [Google Scholar] [CrossRef]
- Parker, A.K.; Hofmann, R.W.; Van Leeuwen, C.; McLachlan, A.R.; Trought, M.C. Manipulating the leaf area to fruit mass ratio alters the synchrony of total soluble solids accumulation and titratable acidity of grape berries. Aust. J. Grape Wine Res. 2015, 21, 266–276. [Google Scholar] [CrossRef]
- Bondada, B.; Covarrubias, J.I.; Tessarin, P.; Boliani, A.C.; Marodin, G.; Rombolà, A.D. Postveraison shoot trimming reduces cluster compactness without compromising fruit quality attributes in organically grown Sangiovese grapevines. Am. J. Enol. Vitic. 2016, 67, 206–211. [Google Scholar] [CrossRef]
- Bobeica, N.; Poni, S.; Hilbert, G.; Renaud, C.; Gomès, E.; Delrot, S.; Dai, Z. Differential responses of sugar, organic acids and anthocyanins to source-sink modulation in Cabernet Sauvignon and Sangiovese grapevines. Front. Plant Sci. 2015, 6, 382. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, P.; Collins, C.; De Bei, R. Leaf removal applied to a sprawling canopy to regulate fruit ripening in Cabernet Sauvignon. Plants 2021, 10, 1017. [Google Scholar] [CrossRef] [PubMed]
- Herrera, J.; Bucchetti, B.; Sabbatini, P.; Comuzzo, P.; Zulini, L.; Vecchione, A.; Peterlunger, E.; Castellarin, S.D. Effect of water deficit and severe shoot trimming on the composition of Vitis vinifera L. Merlot grapes and wines. Aust. J. Grape Wine Res. 2015, 21, 254–265. [Google Scholar] [CrossRef]
- Palliotti, A.; Tombesi, S.; Silvestroni, O.; Lanari, V.; Gatti, M.; Poni, S. Changes in vineyard establishment and canopy management urged by earlier climate-related grape ripening: A review. Sci. Hortic. 2014, 178, 43–54. [Google Scholar] [CrossRef]
- Bubola, M.; Rusjan, D.; Lukić, I. Crop level vs. leaf removal: Effects on Istrian Malvasia wine aroma and phenolic acids composition. Food Chem. 2020, 312, 126046. [Google Scholar] [CrossRef]
- De Bei, R.; Wang, X.; Papagiannis, L.; Fuentes, S.; Gilliham, M.; Tyerman, S.; Collins, C. Shoot thinning of Semillon in a hot climate did not improve yield and berry and wine quality. OENO One 2020, 54, 469–484. [Google Scholar] [CrossRef]
- Poni, S.; Bernizzoni, F.; Presutto, P.; Rebucci, B. Performance of Croatina under short-cane mechanical hedging: A successful case of adaptation. Am. J. Enol. Vitic. 2004, 55, 379–388. [Google Scholar] [CrossRef]
- Coombe, B.G. Growth stages of the grapevine: Adoption of a system for identifying grapevine growth stages. Aust. J. Grape Wine Res. 1995, 1, 104–110. [Google Scholar] [CrossRef]
- Smart, R.; Robinson, M. Sunlight into Wine: A Handbook for Winegrape Canopy Management; Winetitles: Ashford, Australia, 1991. [Google Scholar]
- OIV—International Organization of Vine and Wine. Compendium of International Methods of Wine and Musts Analysis; OIV—International Organization of Vine and Wine: Paris, France, 2012. [Google Scholar]
- Previtali, P.; Dokoozlian, N.; Capone, D.; Wilkinson, K.; Ford, C. Exploratory study of sugar and C6 compounds in single berries of grapevine (Vitis vinifera L.) cv. Cabernet Sauvignon throughout ripening. Aust. J. Grape Wine Res. 2021, 27, 194–205. [Google Scholar] [CrossRef]
- Iland, P.; Bruer, N.; Edwards, G.; Weeks, S.; Wilkes, E. Chemical Analysis of Grapes and Wine: Techniques and Concepts; Patrick Iland Wine Promotions Pty Ltd.: Campbelltown, Australia, 2004. [Google Scholar]
- Tessarin, P.; Parpinello, G.P.; Rombolà, A.D. Physiological and Enological Implications of Postveraison Trimming in an Organically-Managed Vineyard (cv. Sangiovese). Am. J. Enol. Vitic. 2018, 69, 59–69. [Google Scholar] [CrossRef]
- Palliotti, A.; Panara, F.; Silvestroni, O.; Lanari, V.; Sabbatini, P.; Howell, G.S.; Gatti, M.; Poni, S. Influence of mechanical postveraison leaf removal apical to the cluster zone on delay of fruit ripening in Sangiovese (Vitis vinifera L.) grapevines. Aust. J. Grape Wine Res. 2013, 19, 369–377. [Google Scholar] [CrossRef]
- De Bei, R.; Wang, X.; Papagiannis, L.; Cocco, M.; O’Brien, P.; Zito, M.; Ouyang, J.; Fuentes, S.; Gilliham, M.; Tyerman, S. Postveraison leaf removal does not consistently delay ripening in Sémillon and Shiraz in a hot Australian climate. Am. J. Enol. Vitic. 2019, 70, 398–410. [Google Scholar] [CrossRef]
- Valentini, G.; Allegro, G.; Pastore, C.; Colucci, E.; Filippetti, I. Post-veraison trimming slow down sugar accumulation without modifying phenolic ripening in Sangiovese vines. J. Sci. Food Agric. 2019, 99, 1358–1365. [Google Scholar] [CrossRef]
- Ollat, N.; Gaudillere, J. The effect of limiting leaf area during stage I of berry growth on development and composition of berries of Vitis vinifera L. cv. Cabernet Sauvignon. Am. J. Enol. Vitic. 1998, 49, 251–258. [Google Scholar] [CrossRef]
- Schubert, A.; Lovisolo, C.; Restagno, M. Net Photosynthesus of Grapevine Leaves of Different Age Exposed to High or Low Light Intensities. Adv. Hortic. Sci. 1996, 10, 163–166. [Google Scholar]
- Kliewer, W.M.; Dokoozlian, N.K. Leaf area/crop weight ratios of grapevines: Influence on fruit composition and wine quality. Am. J. Enol. Vitic. 2005, 56, 170–181. [Google Scholar] [CrossRef]
- Caccavello, G.; Giaccone, M.; Scognamiglio, P.; Mataffo, A.; Teobaldelli, M.; Basile, B. Vegetative, yield, and berry quality response of aglianico to shoot-trimming applied at three stages of berry ripening. Am. J. Enol. Vitic. 2019, 70, 351–359. [Google Scholar] [CrossRef]
- Bubola, M.; Sivilotti, P.; Diklić, K.; Užila, Z.; Palčić, I.; Plavša, T. Manipulation of ‘Teran’ grape composition with severe shoot trimming and cluster thinning. Acta Hortic. 2017, 1188, 91–96. [Google Scholar] [CrossRef]
- Abad, F.J.; Marin, D.; Loidi, M.; Miranda, C.; Royo, J.B.; Urrestarazu, J.; Santesteban, L.G. Evaluation of the incidence of severe trimming on grapevine (Vitis vinifera L.) water consumption. Agric. Water Manag. 2019, 213, 646–653. [Google Scholar] [CrossRef]
- Poni, S.; Gatti, M.; Bernizzoni, F.; Civardi, S.; Bobeica, N.; Magnanini, E.; Palliotti, A. Late leaf removal aimed at delaying ripening in cv. Sangiovese: Physiological assessment and vine performance. Aust. J. Grape Wine Res. 2013, 19, 378–387. [Google Scholar] [CrossRef]
- Buesa, I.; Caccavello, G.; Basile, B.; Merli, M.C.; Poni, S.; Chirivella, C.; Intrigliolo, D.S. Delaying berry ripening of Bobal and Tempranillo grapevines by late leaf removal in a semi-arid and temperate-warm climate under different water regimes. Aust. J. Grape Wine Res. 2019, 25, 70–82. [Google Scholar] [CrossRef]
- Persic, M.; Mikulic-Petkovsek, M.; Halbwirth, H.; Solar, A.; Veberic, R.; Slatnar, A. Red walnut: Characterization of the phenolic profiles, activities and gene expression of selected enzymes related to the phenylpropanoid pathway in pellicle during walnut development. J. Agric. Food Chem. 2018, 66, 2742–2748. [Google Scholar] [CrossRef] [PubMed]
- Torres, N.; Martínez-Lüscher, J.; Porte, E.; Kurtural, S.K. Optimal ranges and thresholds of grape berry solar radiation for flavonoid biosynthesis in warm climates. Front. Plant Sci. 2020, 11, 931. [Google Scholar] [CrossRef]
- Silvestroni, O.; Lanari, V.; Lattanzi, T.; Palliotti, A.; Vanderweide, J.; Sabbatini, P. Canopy management strategies to control yield and grape composition of Montepulciano grapevines. Aust. J. Grape Wine Res. 2019, 25, 30–42. [Google Scholar] [CrossRef]
- Bubola, M.; Persurić, Đ.; Ganić, K.K. Impact of cluster thinning on productive characteristics and wine phenolic composition of cv. Merlot. J. Food Agric. Environ. 2011, 9, 36–39. [Google Scholar]
- Gatti, M.; Bernizzoni, F.; Civardi, S.; Poni, S. Effects of Cluster Thinning and Preflowering Leaf Removal on Growth and Grape Composition in cv. Sangiovese. Am. J. Enol. Vitic. 2012, 63, 325–332. [Google Scholar] [CrossRef]
- Sivilotti, P.; Falchi, R.; Vanderweide, J.; Sabbatini, P.; Bubola, M.; Vanzo, A.; Lisjak, K.; Peterlunger, E.; Herrera, J.C. Yield reduction through cluster or selective berry thinning similarly modulates anthocyanins and proanthocyanidins composition in Refosco dal peduncolo rosso (Vitis vinifera L.) grapes. Sci. Hortic. 2020, 264, 109166. [Google Scholar] [CrossRef]
- Alba, V.; Natrella, G.; Gambacorta, G.; Crupi, P.; Coletta, A. Effect of over crop and reduced yield by cluster thinning on phenolic and volatile compounds of grapes and wines of ‘Sangiovese’ trained to Tendone. J. Sci. Food Agric. 2022, 102, 7155–7163. [Google Scholar] [CrossRef]
- Poni, S.; Gatti, M.; Palliotti, A.; Dai, Z.; Duchêne, E.; Truong, T.-T.; Ferrara, G.; Matarrese, A.M.S.; Gallotta, A.; Bellincontro, A. Grapevine quality: A multiple choice issue. Sci. Hortic. 2018, 234, 445–462. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Area of Primary Shoot (m2) | Lateral Leaf Area/Primary Shoot (m2) | Total Leaf Area/Shoot (m2) | Leaf Area/Vine (m2) | Leaf Area/Yield (m2/kg) | |
|---|---|---|---|---|---|
| Treatments | |||||
| UC | 0.163 a b | 0.127 a | 0.290 a | 4.02 a | 1.59 a |
| SEV-I | 0.082 b | 0.081 b | 0.163 b | 2.33 b | 0.92 b |
| SEV-II | 0.085 b | 0.051 c | 0.136 c | 1.92 c | 0.77 bc |
| SEV-III | 0.087 b | 0.044 c | 0.131 c | 1.84 c | 0.70 c |
| Sign. a | *** | *** | *** | *** | *** |
| Years | |||||
| 2013 | 0.100 | 0.070 | 0.170 | 2.47 | 1.01 |
| 2014 | 0.109 | 0.073 | 0.183 | 2.44 | 0.94 |
| Sign. | ns | ns | ns | ns | ns |
| Treat × year c | ns | ns | ns | ns | ns |
| Yield/Vine (kg) | Clusters/Vine | Cluster Weight (g) | Shoots/Vine | Clusters/Shoot | Berry Weight (g) | Berries/Cluster | |
|---|---|---|---|---|---|---|---|
| Treatments | |||||||
| UC | 2.58 | 18.9 | 137 | 14.0 | 1.35 | 1.64 | 85 |
| SEV-I | 2.57 | 19.0 | 136 | 14.2 | 1.34 | 1.58 | 86 |
| SEV-II | 2.52 | 19.2 | 132 | 14.3 | 1.35 | 1.62 | 82 |
| SEV-III | 2.64 | 19.3 | 138 | 14.1 | 1.38 | 1.66 | 84 |
| Sign. a | ns | ns | ns | ns | ns | ns | ns |
| Years | |||||||
| 2013 | 2.47 | 20.0 | 124 | 14.6 | 1.37 | 1.48 | 85 |
| 2014 | 2.68 | 18.2 | 148 | 13.7 | 1.33 | 1.77 | 84 |
| Sign. | ns | ns | * | ns | ns | ** | ns |
| Treat × year b | ns | ns | ns | ns | ns | ns | ns |
| Soluble Solids (Brix) | Titratable Acidity (g/L) | pH | Total Anthocyanins (mg/g) | Total Anthocyanins (mg/Berry) | Total Phenolics (mg/g) | Total Phenolics (mg/Berry) | |
|---|---|---|---|---|---|---|---|
| Treatments | |||||||
| UC | 22.7 a b | 6.7 | 3.36 | 0.84 a | 1.34 a | 1.95 | 3.17 |
| SEV-I | 21.3 b | 6.8 | 3.33 | 0.66 b | 1.02 b | 1.82 | 2.85 |
| SEV-II | 20.9 b | 6.9 | 3.32 | 0.69 b | 1.10 b | 1.84 | 2.95 |
| SEV-III | 21.4 b | 6.8 | 3.31 | 0.78 a | 1.28 a | 1.87 | 3.07 |
| Sign. a | ** | ns | ns | * | * | ns | ns |
| Years | |||||||
| 2013 | 22.4 | 6.7 | 3.35 | 0.90 | 1.34 | 2.06 | 3.04 |
| 2014 | 20.8 | 6.9 | 3.31 | 0.59 | 1.04 | 1.68 | 2.98 |
| Sign. | * | ns | ns | ** | * | * | ns |
| Treat × year c | * | ns | ns | ns | ns | ns | ns |
| Season 2015 | Season 2016 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot Trimming | Crop Size | Int. b | Shoot Trimming | Crop Size | Int. | |||||||||
| HC | SST | Sign. a | LCS | HCS | Sign. | Sign. | HC | SST | Sign. | LCS | HCS | Sign. | Sign. | |
| LA of primary shoot (m2) | 0.204 | 0.098 | *** | 0.164 | 0.137 | ns | ns | 0.191 | 0.092 | *** | 0.156 | 0.128 | * | ns |
| Lateral LA/primary shoot (m2) | 0.101 | 0.043 | *** | 0.080 | 0.064 | ns | ns | 0.136 | 0.052 | *** | 0.113 | 0.075 | * | ns |
| Total LA/shoot (m2) | 0.305 | 0.141 | *** | 0.244 | 0.201 | * | ns | 0.327 | 0.144 | *** | 0.269 | 0.203 | ** | * |
| % of laterals | 33 | 31 | ns | 32 | 33 | ns | ns | 41 | 36 | ns | 41 | 36 | ns | ns |
| Total LA/vine (m2) | 4.45 | 2.06 | *** | 2.99 | 3.52 | * | ns | 4.71 | 2.16 | *** | 3.16 | 3.72 | ns | ns |
| LA/yield (m2/kg) | 1.70 | 0.82 | *** | 1.27 | 1.25 | ns | ns | 1.80 | 0.86 | *** | 1.41 | 1.25 | ns | ns |
| Canopy gaps (%) | 4.7 | 4.3 | ns | 5.2 | 3.8 | ns | ns | 3.8 | 3.5 | ns | 4.2 | 3.2 | ns | ns |
| Leaf layer number | 2.32 | 2.29 | ns | 2.13 | 2.48 | ns | ns | 2.51 | 2.44 | ns | 2.38 | 2.57 | ns | ns |
| PAR c (% ambient) | 8 | 15 | *** | 12 | 11 | ns | ns | 7 | 13 | ** | 10 | 9 | ns | ns |
| Season 2015 | Season 2016 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot Trimming | Crop Size | Int. b | Shoot Trimming | Crop Size | Int. | |||||||||
| HC | SST | Sign. a | LCS | HCS | Sign. | Sign. | HC | SST | Sign. | LCS | HCS | Sign. | Sign. | |
| Yield/vine (kg) | 2.61 | 2.53 | ns | 2.36 | 2.78 | * | ns | 2.65 | 2.59 | ns | 2.29 | 2.96 | * | ns |
| Clusters/vine | 22.0 | 22.2 | ns | 19.4 | 24.9 | ** | ns | 24.2 | 23.7 | ns | 19.8 | 28.1 | *** | ns |
| Cluster weight (kg) | 119 | 115 | ns | 122 | 112 | * | ns | 111 | 111 | ns | 116 | 106 | ns | ns |
| Shoots/vine | 14.7 | 15.1 | ns | 12.2 | 17.7 | *** | ns | 15.0 | 15.2 | ns | 11.9 | 18.2 | *** | ns |
| Clusters/shoot | 1.51 | 1.5 | ns | 1.59 | 1.42 | * | ns | 1.63 | 1.57 | ns | 1.66 | 1.54 | ns | ns |
| Berry weight | 1.49 | 1.44 | ns | 1.48 | 1.45 | ns | ns | 1.55 | 1.54 | ns | 1.59 | 1.50 | ns | ns |
| Berries/cluster | 80 | 81 | ns | 83 | 78 | ns | ns | 71 | 72 | ns | 73 | 71 | ns | ns |
| Season 2015 | Season 2016 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shoot Trimming | Crop Size | Int. b | Shoot Trimming | Crop Size | Int. | |||||||||
| HC | SST | Sign. a | LCS | HCS | Sign. | Sign. | HC | SST | Sign. | LCS | HCS | Sign. | Sign. | |
| Soluble solids (Brix) | 23.2 | 21.4 | *** | 22.5 | 22.1 | ns | ns | 22.8 | 21.4 | * | 22.7 | 21.5 | * | ns |
| Titratable acidity (g/L) | 6.4 | 6.7 | ns | 6.5 | 6.6 | ns | ns | 6.8 | 6.8 | ns | 6.9 | 6.8 | ns | ns |
| pH | 3.37 | 3.36 | ns | 3.39 | 3.34 | ns | ns | 3.26 | 3.29 | ns | 3.28 | 3.27 | ns | ns |
| Total anthocyanins (mg/g) | 0.97 | 0.91 | ns | 0.95 | 0.92 | ns | ns | 0.83 | 0.78 | ns | 0.85 | 0.76 | * | ns |
| Total anthocyanins (mg/berry) | 1.44 | 1.30 | ns | 1.41 | 1.34 | ns | ns | 1.29 | 1.22 | ns | 1.35 | 1.16 | * | ns |
| Total phenolics (mg/g) | 2.20 | 2.08 | ns | 2.14 | 2.15 | ns | ns | 2.06 | 1.99 | ns | 2.08 | 1.97 | ns | ns |
| Total phenolics (mg/berry) | 3.28 | 3.00 | ns | 3.17 | 3.11 | ns | ns | 3.20 | 3.08 | ns | 3.30 | 2.98 | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bubola, M.; Persic, M.; Rossi, S.; Bestulić, E.; Zdunić, G.; Plavša, T.; Radeka, S. Severe Shoot Trimming and Crop Size as Tools to Modulate Cv. Merlot Berry Composition. Plants 2022, 11, 3571. https://doi.org/10.3390/plants11243571
Bubola M, Persic M, Rossi S, Bestulić E, Zdunić G, Plavša T, Radeka S. Severe Shoot Trimming and Crop Size as Tools to Modulate Cv. Merlot Berry Composition. Plants. 2022; 11(24):3571. https://doi.org/10.3390/plants11243571
Chicago/Turabian StyleBubola, Marijan, Martina Persic, Sara Rossi, Ena Bestulić, Goran Zdunić, Tomislav Plavša, and Sanja Radeka. 2022. "Severe Shoot Trimming and Crop Size as Tools to Modulate Cv. Merlot Berry Composition" Plants 11, no. 24: 3571. https://doi.org/10.3390/plants11243571
APA StyleBubola, M., Persic, M., Rossi, S., Bestulić, E., Zdunić, G., Plavša, T., & Radeka, S. (2022). Severe Shoot Trimming and Crop Size as Tools to Modulate Cv. Merlot Berry Composition. Plants, 11(24), 3571. https://doi.org/10.3390/plants11243571