
Stimulating the Growth, Anabolism, Antioxidants, and Yield of Rice Plants Grown under Salt Stress by Combined Application of Bacterial Inoculants and Nano-Silicon
 ,
,  , ,
, ,  and
and
Abstract
1. Introduction
2. Results
2.1. Activity of Soil Enzymes
2.2. Physiological Characteristics
2.2.1. K/Na Ratio
2.2.2. Photosynthetic Pigments
2.2.3. Water Relations
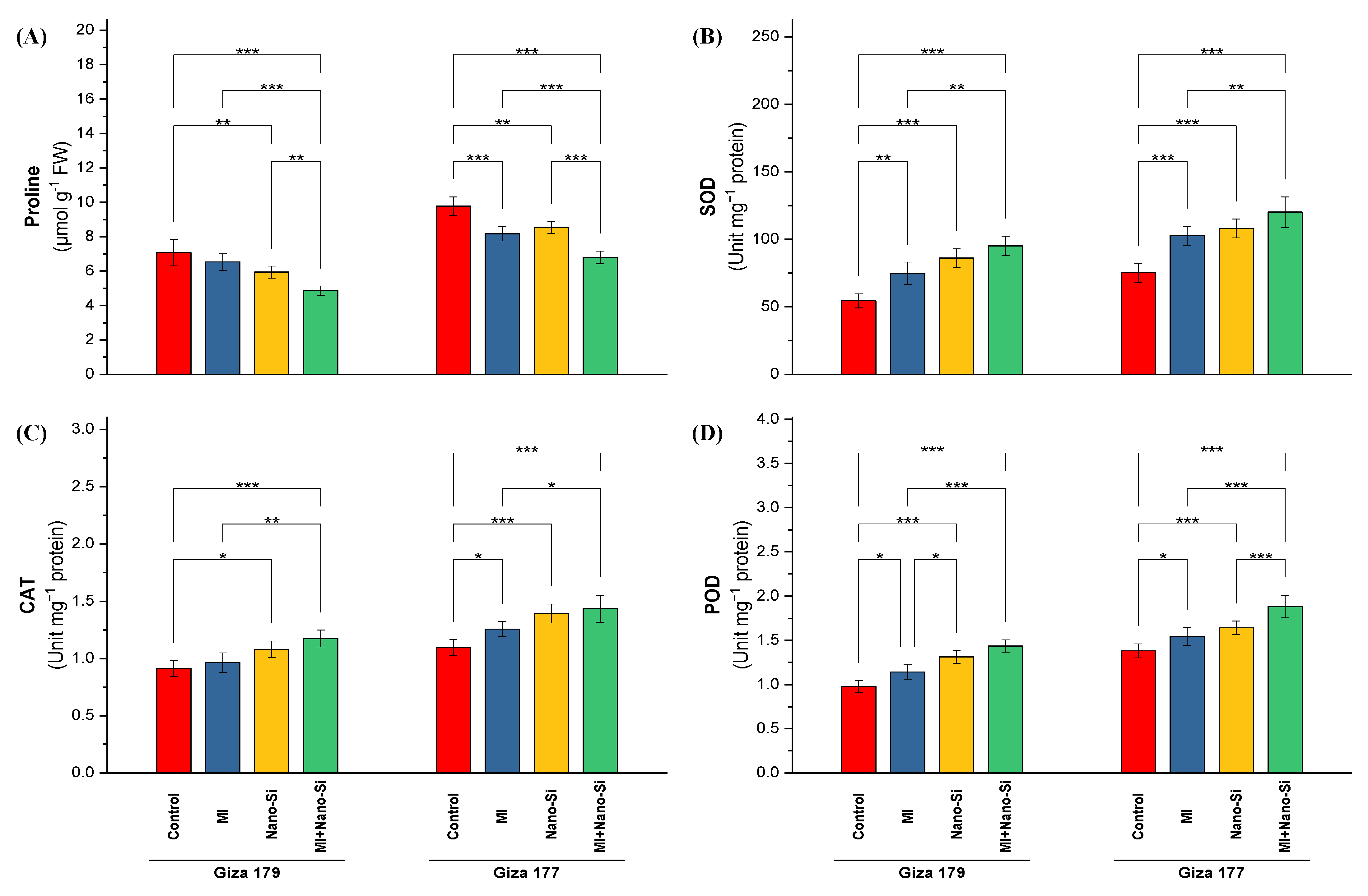
2.2.4. Oxidative Stress Indicators
2.2.5. Antioxidant Defense System
2.2.6. Vegetative Characteristics
2.2.7. Reproductive Growth Characteristics
2.2.8. Content of N, P, K, Na, and Si in Rice Grains
3. Discussion
4. Materials and Methods
4.1. Experimental Layout and Treatments
4.2. Soil Enzymes Activity
4.3. Physiological Characteristics
4.3.1. Leaf Na+ and K+ Determination
4.3.2. Chlorophylls and Carotenoid
4.3.3. Water Relations
Relative Water Content (RWC)
Stomatal Conductance
4.3.4. Oxidative Stress Indicators
Lipid Peroxidation
H2O2 Content
Electrolyte Leakage
4.3.5. Antioxidant System
Proline Content
Antioxidant Enzymes Activity
Superoxide Dismutase (SOD)
Catalase (CAT)
Peroxidase (POD)
4.3.6. Vegetative Growth Attributes
Leaf Area Index (LAI)
Root Length
4.3.7. Reproductive Growth Attributes
Crop Yield
Nutrients Content in Rice Grains
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seck, P.A.; Diagne, A.; Mohanty, S.; Wopereis, M.C.S. Crops that feed the world 7: Rice. Food Secur. 2012, 4, 7–24. [Google Scholar] [CrossRef]
- FAO. FAOSTAT Database; FAO: Rome, Italy, 2020. [Google Scholar]
- Singh, K. Microbial and enzyme activities of saline and sodic soils. Land Degrad. Develop. 2016, 27, 706–718. [Google Scholar] [CrossRef]
- Reddy, I.N.B.L.; Kim, B.-K.; Yoon, I.-S.; Kim, K.-H.; Kwon, T.-R. Salt tolerance in rice: Focus on mechanisms and approaches. Rice Sci. 2017, 24, 123–144. [Google Scholar] [CrossRef]
- Gorham, J.; Jones, R.G.W.; McDonnell, E. Some mechanisms of salt tolerance in crop plants. Plant Soil 1985, 89, 15–40. [Google Scholar] [CrossRef]
- Abdel Megeed, T.; Gharib, H.; Hafez, E.; El-Sayed, A. Effect of some plant growth regulators and biostimulants on the productivity of Sakha108 rice plant (Oryza sativa L.) under different water stress conditions. Appl. Ecol. Environ. Res 2021, 19, 2859–2878. [Google Scholar] [CrossRef]
- Salim, B.B.M.; Hikal, M.S.; Osman, H.S. Ameliorating the deleterious effects of saline water on the antioxidants defense system and yield of eggplant using foliar application of zinc sulphate. Ann. Agric. Sci. 2019, 64, 244–251. [Google Scholar] [CrossRef]
- Khush, G.S. What it will take to Feed 5.0 Billion Rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef]
- Hafez, E.M.; Abou El Hassan, W.H.; Gaafar, I.A.; Seleiman, M.F. Effect of gypsum application and irrigation intervals on clay saline-sodic soil characterization, rice water use efficiency, growth, and yield. J. Agric. Sci. 2015, 7, 208. [Google Scholar] [CrossRef]
- Hafez, E.M.; Alsohim, A.S.; Farig, M.; Omara, A.E.-D.; Rashwan, E.; Kamara, M.M. Synergistic effect of biochar and plant growth promoting rhizobacteria on alleviation of water deficit in rice plants under salt-affected soil. Agronomy 2019, 9, 847. [Google Scholar] [CrossRef]
- Kheir, A.M.S.; Abouelsoud, H.M.; Hafez, E.M.; Ali, O.A.M. Integrated effect of nano-Zn, nano-Si, and drainage using crop straw–filled ditches on saline sodic soil properties and rice productivity. Arab. J. Geosci. 2019, 12, 471. [Google Scholar] [CrossRef]
- Solyman, S.N.; Abdel-Monem, M.; Abou-Taleb, K.; Osman, H.S.; El-Sharkawy, R.M. Production of plant growth regulators by some fungi isolated under salt stress. S. Asian J. Res. Microbio. 2019, 3, 1–10. [Google Scholar] [CrossRef]
- Hafez, E.M.; Gowayed, S.M.; Nehela, Y.; Sakran, R.M.; Rady, A.M.S.; Awadalla, A.; Omara, A.E.-D.; Alowaiesh, B.F. Incorporated biochar-based soil amendment and exogenous glycine betaine foliar application ameliorate rice (Oryza sativa L.) tolerance and resilience to osmotic stress. Plants 2021, 10, 1930. [Google Scholar] [CrossRef] [PubMed]
- Osman, H.S.; Gowayed, S.M.; Elbagory, M.; Omara, A.E.; El-Monem, A.M.A.; Abd El-Razek, U.A.; Hafez, E.M. Interactive impacts of beneficial microbes and Si-Zn nanocomposite on growth and productivity of soybean subjected to water deficit under salt-affected soil conditions. Plants 2021, 10, 1396. [Google Scholar] [CrossRef] [PubMed]
- Bano, A.; Fatima, M. Salt tolerance in Zea mays (L.) following inoculation with Rhizobium and Pseudomonas. Biol. Fertil. Soils 2009, 45, 405–413. [Google Scholar] [CrossRef]
- Omara, A.E.-D.; Hafez, E.M.; Osman, H.S.; Rashwan, E.; El-Said, M.A.A.; Alharbi, K.; Abd El-Moneim, D.; Gowayed, S.M. Collaborative impact of compost and beneficial rhizobacteria on soil properties, physiological attributes, and productivity of wheat subjected to deficit irrigation in salt affected soil. Plants 2022, 11, 877. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, A.; Khan, M.A.; Lee, K.E.; Kang, S.M.; Dhungana, S.K.; Bhusal, N.; Lee, I.J. The halotolerant rhizobacterium-Pseudomonas koreensis MU2 enhances inorganic silicon and phosphorus use efficiency and augments salt stress tolerance in soybean (Glycine max L.). Microorganisms 2020, 8, 1256. [Google Scholar] [CrossRef]
- El-Shamy, M.A.; Alshaal, T.; Mohamed, H.H.; Rady, A.M.S.; Hafez, E.M.; Alsohim, A.S.; Abd El-Moneim, D. Quinoa response to application of Phosphogypsum and plant growth-promoting Rhizobacteria under water stress associated with salt-affected soil. Plants 2022, 11, 872. [Google Scholar] [CrossRef]
- Hafez, E.M.; Osman, H.S.; Gowayed, S.M.; Okasha, S.A.; Omara, A.E.-D.; Sami, R.; Abd El-Monem, A.M.; Abd El-Razek, U.A. Minimizing the adversely impacts of water deficit and soil salinity on maize growth and productivity in response to the application of plant growth-promoting rhizobacteria and silica nanoparticles. Agronomy 2021, 11, 676. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Ashfaque, F.; Chhillar, H.; Irfan, M.; Khan, N.A. The intricacy of silicon, plant growth regulators and other signaling molecules for abiotic stress tolerance: An entrancing crosstalk between stress alleviators. Plant Physiol. Biochem. 2021, 162, 36–47. [Google Scholar] [CrossRef]
- Pan, T.; Zhang, J.; He, L.; Hafeez, A.; Ning, C.; Cai, K. Silicon enhances plant resistance of rice against submergence stress. Plants 2021, 10, 767. [Google Scholar] [CrossRef]
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of silicon nanoparticles in agriculture. 3 Biotech 2019, 9, 90. [Google Scholar] [CrossRef] [PubMed]
- Alharbi, K.; Rashwan, E.; Mohamed, H.H.; Awadalla, A.; Omara, A.E.-D.; Hafez, E.M.; Alshaal, T. Application of silica nanoparticles in combination with two bacterial strains improves the growth, antioxidant capacity and production of barley irrigated with saline water in salt-affected soil. Plants 2022, 11, 2026. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-Ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. Int. 2015, 22, 15416–15431. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.M.; Osman, H.S.; El-Razek, U.A.A.; Elbagory, M.; Omara, A.E.-D.; Eid, M.A.; Gowayed, S.M. Foliar-applied potassium silicate coupled with plant growth-promoting rhizobacteria improves growth, physiology, nutrient uptake and productivity of faba bean (Vicia faba L.) irrigated with saline water in salt-affected soil. Plants 2021, 10, 894. [Google Scholar] [CrossRef] [PubMed]
- Salim, B.B.M.; Abou El-Yazied, A.; Salama, Y.A.M.; Raza, A.; Osman, H.S. Impact of silicon foliar application in enhancing antioxidants, growth, flowering and yield of squash plants under deficit irrigation condition. Ann. Agric. Sci. 2021, 66, 176–183. [Google Scholar] [CrossRef]
- Shukla, R.; Bansal, V.; Chaudhary, M.; Basu, A.; Bhonde, R.R.; Sastry, M. Biocompatibility of gold nanoparticles and their endocytotic fate inside the cellular compartment: A microscopic overview. Langmuir 2005, 21, 10644–10654. [Google Scholar] [CrossRef]
- Karunakaran, G.; Suriyaprabha, R.; Manivasakan, P.; Yuvakkumar, R.; Rajendran, V.; Prabu, P.; Kannan, N. Effect of nanosilica and silicon sources on plant growth promoting rhizobacteria, soil nutrients and maize seed germination. IET Nanobiotechnol. 2013, 7, 70–77. [Google Scholar] [CrossRef]
- Rajput, V.D.; Minkina, T.; Feizi, M.; Kumari, A.; Khan, M.; Mandzhieva, S.; Sushkova, S.; El-Ramady, H.; Verma, K.K.; Singh, A.; et al. Effects of silicon and silicon-based nanoparticles on rhizosphere microbiome, plant stress and growth. Biology 2021, 10, 791. [Google Scholar] [CrossRef]
- Chaudhary, P.; Sharma, A. Response of nanogypsum on the performance of plant growth promotory bacteria re-covered from nanocompound infested agriculture field. Env. Ecol. 2019, 37, 363–372. [Google Scholar]
- Seleiman, M.F.; Hafez, E.M. Optimizing inputs management for sustainable agricultural development. In Mitigating Environmental Stresses for Agricultural Sustainability in Egypt; Awaad, H., Abu-hashim, M., Negm, A., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 487–507. [Google Scholar]
- Hafez, E.; Farig, M. Efficacy of salicylic acid as a cofactor for ameliorating effects of water stress and enhancing wheat yield and water use efficiency in saline soil. Inter. J. Plant Prod. 2019, 13, 163–176. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, F.M.A.; Mazrou, Y.S.A.; Khalaf, A.E.A.; El-Sherif, A.M.A.; Osman, H.S.; Hafez, E.M.; Eid, M.A.M. Utilization efficiency of growth regulators in wheat under drought stress and sandy soil conditions. Agronomy 2021, 11, 1760. [Google Scholar] [CrossRef]
- Ullah, S.; Bano, A. Isolation of plant-growth-promoting rhizobacteria from rhizospheric soil of halophytes and their impact on maize (Zea mays L.) under induced soil salinity. Can. J. Microbiol. 2015, 61, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Abdelrasheed, K.G.; Mazrou, Y.; Omara, A.E.-D.; Osman, H.S.; Nehela, Y.; Hafez, E.M.; Rady, A.M.S.; El-Moneim, D.A.; Alowaiesh, B.F.; Gowayed, S.M. Soil amendment using biochar and application of K-humate enhance the growth, productivity, and nutritional value of onion (Allium cepa L.) under deficit irrigation conditions. Plants 2021, 10, 2598. [Google Scholar] [CrossRef]
- Zhu, Y.; Jiang, X.; Zhang, J.; He, Y.; Zhu, X.; Zhou, X.; Gong, H.; Yin, J.; Liu, Y. Silicon confers cucumber resistance to salinity stress through regulation of proline and cytokinins. Plant Physiol. Biochem. 2020, 156, 209–220. [Google Scholar] [CrossRef]
- Zafar-ul-Hye, M.; Muhammad, H.; Zahir, F.; Ahmad, Z.; Hussain, M.; Hussain, A. Application of ACC-deaminase containing rhizobacteria with fertilizer improves maize production under drought and salinity stress. Int. J. Agric. Biol. 2014, 16, 591–596. [Google Scholar]
- Raheem, A.; Shaposhnikov, A.; Belimov, A.A.; Dodd, I.C.; Ali, B. Auxin production by rhizobacteria was associated with improved yield of wheat (Triticum aestivum L.) under drought stress. Arch. Agron. Soil Sci. 2018, 64, 574–587. [Google Scholar] [CrossRef]
- Youssif, N.E.E.; Osman, H.S.M.; Salama, Y.A.M.; Zaghlool, S.A.M. Effect of rice straw and applications of potassium silicate, potassium humate and seaweed extract on growth and some macronutrients of sweet pepper plants under irrigation deficit. Arab Univ. J. Agric. Sci. 2018, 26, 755–773. [Google Scholar] [CrossRef]
- Kalteh, M.; Alipour, Z.T.; Ashraf, S.; Marashi Aliabadi, M.; Falah Nosratabadi, A. Effect of silica nanoparticles on basil (Ocimum basilicum) under salinity stress. J. Chem. Health Risks 2018, 4, 49–55. [Google Scholar]
- Osman, H.S.; Salim, B.B.M. Influence of exogenous application of some phytoprotectants on growth, yield and pod quality of snap bean under NaCl salinity. Ann. Agric. Sci. 2016, 61, 1–13. [Google Scholar] [CrossRef][Green Version]
- Osman, H.S.; Salim, B.B. Improving yield and quality of kohlrabi stems growing under NaCl salinity using foliar application of urea and seaweed extract. J. Hort. Sci. Ornament. Plant 2016, 8, 149–160. [Google Scholar]
- Karimi, J.; Mohsenzadeh, S. Effects of silicon oxide nanoparticles on growth and physiology of wheat seedlings. Russ. J. Plant Physiol. 2016, 63, 119–123. [Google Scholar] [CrossRef]
- Raza, A.; Salehi, H.; Rahman, M.A.; Zahid, Z.; Madadkar Haghjou, M.; Najafi-Kakavand, S.; Charagh, S.; Osman, H.S.; Albaqami, M.; Zhuang, Y.; et al. Plant hormones and neurotransmitter interactions mediate antioxidant defenses under induced oxidative stress in plants. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Danish, S.; Zafar-Ul-Hye, M.; Hussain, S.; Riaz, M.; Qayyum, M.F. Mitigation of drought stress in maize through inoculation with drought tolerant ACC deaminase containing PGPR under axenic conditions. Pak. J. Bot. 2020, 52, 49–60. [Google Scholar] [CrossRef]
- Biju, S.; Fuentes, S.; Gupta, D. Silicon improves seed germination and alleviates drought stress in lentil crops by regulating osmolytes, hydrolytic enzymes and antioxidant defense system. Plant Physiol. Biochem. 2017, 119, 250–264. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Ashraf, M. Influence of foliar application of silicon on chlorophyll fluorescence, photosynthetic pigments, and growth in water-stressed wheat cultivars differing in drought tolerance. Turk. J. Bot. 2015, 39, 625–634. [Google Scholar] [CrossRef]
- Li, B.; Tao, G.; Xie, Y.; Cai, X. Physiological effects under the condition of spraying nano-SiO2 onto the Indocalamus barbatus McClure leaves. J. Nanjing For. Univ. 2012, 36, 161–164. [Google Scholar]
- Esmaili, S.; Tavallali, V.; Amiri, B. Nano-silicon complexes enhance growth, yield, water relations and mineral composition in tanacetum parthenium under water deficit stress. Silicon 2021, 13, 2493–2508. [Google Scholar] [CrossRef]
- Khan, Z.S.; Rizwan, M.; Hafeez, M.; Ali, S.; Adrees, M.; Qayyum, M.F.; Khalid, S.; ur Rehman, M.Z.; Sarwar, M.A. Effects of silicon nanoparticles on growth and physiology of wheat in cadmium contaminated soil under different soil moisture levels. Environ. Sci. Pollut. Res. 2020, 27, 4958–4968. [Google Scholar] [CrossRef]
- Wu, Y.; Li, Y.; Zheng, C.; Zhang, Y.; Sun, Z. Organic amendment application influence soil organism abundance in saline alkali soil. Eur. J. Soil Biol. 2013, 54, 32–40. [Google Scholar] [CrossRef]
- Osman, H.S.; Rady, A.M.S.; Awadalla, A.; Omara, A.E.-D.; Hafez, E.M. Improving the antioxidants system, growth, and sugar beet quality subjected to long-term osmotic stress by phosphate solubilizing bacteria and compost tea. Inter. J. Plant Prod. 2022, 16, 119–135. [Google Scholar] [CrossRef]
- Hafez, E.; Gharib, H. Effect of exogenous application of ascorbic acid on physiological and biochemical characteristics of wheat under water stress. Inter. J. Plant Prod. 2016, 10, 579–596. [Google Scholar]
- Ibrahim, M.A.; Merwad, A.-R.M.; Elnaka, E.A. Rice (Oryza Sativa L.) tolerance to drought can be improved by silicon application. Comm. Soil Sci. Plant Anal. 2018, 49, 945–957. [Google Scholar] [CrossRef]
- Ding, Z.; Kheir, A.M.S.; Ali, O.A.M.; Hafez, E.M.; ElShamey, E.A.; Zhou, Z.; Wang, B.; Lin, X.E.; Ge, Y.; Fahmy, A.E.; et al. A vermicompost and deep tillage system to improve saline-sodic soil quality and wheat productivity. J. Environ. Manag. 2021, 277, 111388. [Google Scholar] [CrossRef]
- Hafez, E.M.; Ragab, A.Y.; Kobata, T. Water-use efficiency and ammonium-N source applied of wheat under irrigated and desiccated conditions. Int. J. Plant Soil Sci. 2014, 3, 1302–1316. [Google Scholar] [CrossRef]
- Nehela, Y.; Mazrou, Y.S.A.; Alshaal, T.; Rady, A.M.S.; El-Sherif, A.M.A.; Omara, A.E.-D.; Abd El-Monem, A.M.; Hafez, E.M. The integrated amendment of sodic-saline soils using biochar and plant growth-promoting rhizobacteria enhances maize (Zea mays L.) resilience to water salinity. Plants 2021, 10, 1960. [Google Scholar] [CrossRef]
- Nafees, M.; Fahad, S.; Shah, A.N.; Bukhari, M.A.; Maryam; Ahmed, I.; Ahmad, S.; Hussain, S. Reactive Oxygen species signaling in plants. In Plant Abiotic Stress Tolerance: Agronomic, Molecular and Biotechnological Approaches; Hasanuzzaman, M., Hakeem, K.R., Nahar, K., Alharby, H.F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 259–272. [Google Scholar]
- Akhtar, N.; Ilyas, N.; Mashwani, Z.-U.-R.; Hayat, R.; Yasmin, H.; Noureldeen, A.; Ahmad, P. Synergistic effects of plant growth promoting rhizobacteria and silicon dioxide nano-particles for amelioration of drought stress in wheat. Plant Physiol. Biochem. 2021, 166, 160–176. [Google Scholar] [CrossRef]
- Chung, W.-H. Unraveling new functions of superoxide dismutase using yeast model system: Beyond its conventional role in superoxide radical scavenging. J. Microbiol. 2017, 55, 409–416. [Google Scholar] [CrossRef]
- Ahmad, R.; Hussain, S.; Anjum, M.A.; Khalid, M.F.; Saqib, M.; Zakir, I.; Hassan, A.; Fahad, S.; Ahmad, S. Oxidative stress. In Plant Abiotic Stress Tolerance: Agronomic, Molecular and Biotechnological Approaches; Hasanuzzaman, M., Hakeem, K.R., Nahar, K., Alharby, H.F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 191–205. [Google Scholar]
- Hafez, E.M.; Omara, A.E.D.; Alhumaydhi, F.A.; El-Esawi, M.A. Minimizing hazard impacts of soil salinity and water stress on wheat plants by soil application of vermicompost and biochar. Physiol. Plant. 2020, 172, 587–602. [Google Scholar] [CrossRef]
- Asgari, F.; Majd, A.; Jonoubi, P.; Najafi, F. Effects of silicon nanoparticles on molecular, chemical, structural and ultrastructural characteristics of oat (Avena sativa L.). Plant Physiol. Biochem. 2018, 127, 152–160. [Google Scholar] [CrossRef]
- Hafez, E.; Omara, A.E.D.; Ahmed, A. The coupling effects of plant growth promoting rhizobacteria and salicylic acid on physiological modifications, yield traits, and productivity of wheat under water deficient conditions. Agronomy 2019, 9, 524. [Google Scholar] [CrossRef]
- Singh, R.P.; Jha, P.N. Analysis of fatty acid composition of PGPR Klebsiella sp. SBP-8 and its role in ameliorating salt stress in wheat. Symbiosis 2017, 73, 213–222. [Google Scholar] [CrossRef]
- Al-Enazy, A.-A.; Al-Barakah, F.; Al-Oud, S.; Usman, A. Effect of phosphogypsum application and bacteria co-inoculation on biochemical properties and nutrient availability to maize plants in a saline soil. Arch. Agron. Soil Sci. 2018, 64, 1394–1406. [Google Scholar] [CrossRef]
- Ayuso-Calles, M.; Flores-Félix, J.D.; Rivas, R. Overview of the role of rhizobacteria in plant salt stress tolerance. Agronomy 2021, 11, 1759. [Google Scholar] [CrossRef]
- Ismail, L.M.; Soliman, M.I.; Abd El-Aziz, M.H.; Abdel-Aziz, H.M.M. Impact of silica ions and nano silica on growth and productivity of pea plants under salinity stress. Plants 2022, 11, 494. [Google Scholar] [CrossRef]
- Hassan, I.F.; Ajaj, R.; Gaballah, M.S.; Ogbaga, C.C.; Kalaji, H.M.; Hatterman-Valenti, H.M.; Alam-Eldein, S.M. Foliar application of nano-silicon improves the physiological and biochemical characteristics of ‘Kalamata’ olive subjected to deficit irrigation in a semi-arid climate. Plants 2022, 11, 1561. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Öhlinger, H.; Von Mersi, W. Enzymes involved in intracellular metabolism. In Methods in Soil Biology; Schinner, F., Öhlinger, R., Kandeler, E., Margesin, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1996; pp. 235–245. [Google Scholar]
- Temminghoff, E.E.; Houba, V.J. Plant Analysis Procedures; Springer: Berlin/Heidelberg, Germany, 2004; p. 179. [Google Scholar]
- Peng, Y.; Liu, E. A comparative study of methods of extracting chlorophyll. Acta Agric. Univ. Pekin. 1992, 18, 247–250. [Google Scholar]
- Weatherley, P.E. Studies in the water relations of the cotton plant. I. The field measurement of water deficits in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Du, Z.; Bramlage, W.J. Modified thiobarbituric acid assay for measuring lipid oxidation in sugar-rich plant tissue extracts. J. Agric. Food Chem. 1992, 40, 1566–1570. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Naghashzadeh, M. Response of relative water content and cell membrane stability to mycorrhizal biofertilizer in maize. Electron. J. Biol. 2014, 10, 68–72. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Osman, H.S. Enhancing antioxidant–yield relationship of pea plant under drought at different growth stages by exogenously applied glycine betaine and proline. Ann. Agric. Sci. 2015, 60, 389–402. [Google Scholar] [CrossRef]
- Osman, H.S.; Salim, B.B.M. Enhancing antioxidants defense system of snap bean under NaCl salinity using foliar application of salicylic acid, spermidine and glycine betaine. Am. Eurasian J. Agric. Environ. Sci. 2016, 16, 1200–1210. [Google Scholar]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Analyt. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; pp. 121–126. [Google Scholar]
- Vetter, J.L.; Steinberg, M.P.; Nelson, A.I. Enzyme assay, quantitative determination of peroxidase in sweet corn. J. Agric. Food Chem. 1958, 6, 39–41. [Google Scholar] [CrossRef]
- Evans, G.C. The Quantitative Analysis of Plant Growth; University of California Press: Oakland, CA, USA, 1972. [Google Scholar]
- Adhikari, B.B.; Mehera, B.; Haefele, S.M. Selection of drought tolerant rice varieties for the western mid hills of Nepal. J. Inst. Agric. Anim. Sci. 2015, 33–34, 195–206. [Google Scholar] [CrossRef][Green Version]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Sumner, M.E. Methods of Soil Analysis: Part 3 Chemical Methods; Soil Science Society of America, Inc., American Society of Agronomy, Inc.: Madison, WI, USA, 1996. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2005. [Google Scholar]
- Novozamsky, I.; van Eck, R.; Houba, V.J.G. A rapid determination of silicon in plant material. Comm. Soil Sci. Plant Anal. 1984, 15, 205–211. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Treatments | Urease (mg N-NH4+ g−1 Soil h−1) | Dehydrogenase (mg TPF g−1 DM Soil d−1) | ||
|---|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | ||
| Giza 179 | Control | 103.3 ± 2.4 de | 111.5 ± 3.6 d | 53.2 ± 3.1 g | 56.8 ± 1.5 f |
| MI | 155.1 ± 1.8 b | 170 ± 2.2 b | 104 ± 2.1 c | 108.8 ± 1.8 c | |
| Nano-Si | 137.1 ± 3.7 c | 147.9 ± 2.6 c | 84.2 ± 1.2 d | 88.6 ± 1.6 d | |
| MI + nano-Si | 202.8 ± 2.4 a | 212 ± 3.5 a | 139.6 ± 4.1 a | 149 ± 3.3 a | |
| Giza 177 | Control | 73 ± 1.7 f | 81.9 ± 2.1 f | 30.9 ± 2 h | 46.2 ± 2.4 g |
| MI | 106.3 ± 3.4 d | 110.3 ± 2.7 de | 77.2 ± 0.7 e | 90.5 ± 1.6 d | |
| Nano-Si | 96.3 ± 2.2 e | 104 ± 3.6 e | 63.1 ± 0.8 f | 76.9 ± 2.6 e | |
| MI + nano-Si | 140.8 ± 2.4 c | 146.9 ± 1.6 c | 110.7 ± 1.9 b | 117 ± 1.4 b | |
| Cultivars | *** | ** | ** | ** | |
| Treatments | *** | *** | *** | *** | |
| Cultivars X Treatments | *** | *** | * | *** | |
| Cultivar | Treatments | Na (mg g−1 DW) | K (mg g−1 DW) | K/Na (Ratio) | |||
|---|---|---|---|---|---|---|---|
| 2020 | 2021 | 2020 | 2021 | 2020 | 2021 | ||
| Giza 179 | Control | 2.6 ± 0.1 cd | 2.74 ± 0.09 d | 0.88 ± 0.15 d | 0.93 ± 0.08 d | 0.34 ± 0.05 de | 0.35 ± 0.02 d |
| MI | 2.18 ± 0.1 ef | 2.18 ± 0.15 f | 1.23 ± 0.12 bc | 1.34 ± 0.04 c | 0.56 ± 0.03 c | 0.62 ± 0.06 c | |
| Nano-Si | 1.96 ± 0.11 fg | 2.04 ± 0.1 f | 1.45 ± 0.06 b | 1.56 ± 0.08 b | 0.74 ± 0.01 b | 0.77 ± 0.07 b | |
| MI + nano-Si | 1.65 ± 0.1 g | 1.78 ± 0.04 g | 1.77 ± 0.12 a | 1.91 ± 0.14 a | 1.08 ± 0.11 a | 1.07 ± 0.1 a | |
| Giza 177 | Control | 3.52 ± 0.11 a | 3.68 ± 0.1 a | 0.22 ± 0.07 f | 0.35 ± 0.06 f | 0.06 ± 0.02 g | 0.1 ± 0.01 f |
| MI | 3.14 ± 0.32 b | 3.27 ± 0.14 b | 0.57 ± 0.07 e | 0.69 ± 0.09 e | 0.18 ± 0.04 fg | 0.21 ± 0.02 ef | |
| Nano-Si | 2.93 ± 0.11 bc | 3.06 ± 0.09 c | 0.81 ± 0.04 de | 0.83 ± 0.04 de | 0.28 ± 0.02 ef | 0.27 ± 0.01 de | |
| MI + nano-Si | 2.34 ± 0.11 de | 2.45 ± 0.17 e | 1.05 ± 0.1 cd | 1.20 ± 0.06 c | 0.45 ± 0.03 cd | 0.49 ± 0.05 c | |
| Cultivars | * | ** | ** | ** | *** | ** | |
| Treatments | *** | *** | *** | *** | *** | *** | |
| Cultivars X Treatments | ns | *** | ns | ns | *** | *** | |
| Season | Cultivar | Treatments | N (g kg−1 DW) | P (g kg−1 DW) | K (g kg−1 DW) | Na (g kg−1 DW) | Si (g kg−1 DW) |
|---|---|---|---|---|---|---|---|
| 2020 | Giza 179 | Control | 1.30 ± 0.04 ce | 0.41 ± 0.04 cd | 0.51 ± 0.05 e | 0.44 ± 0.04 cd | 0.33 ± 0.02 d |
| MI | 1.46 ± 0.08 ac | 0.54 ± 0.03 cd | 0.81 ± 0.04 c | 0.32 ± 0.04 de | 0.48 ± 0.04 bc | ||
| Si-NPs | 1.49 ± 0.07 ab | 0.77 ± 0.06 b | 1.05 ± 0.08 b | 0.24 ± 0.02 e | 0.51 ± 0.04 b | ||
| MI + Si-NPs | 1.53 ± 0.04 a | 1.01 ± 0.09 a | 1.33 ± 0.08 a | 0.19 ± 0.01 e | 0.58 ± 0.03 a | ||
| Giza 177 | Control | 1.18 ± 0.07 e | 0.15 ± 0.06 e | 0.29 ± 0.06 f | 0.79 ± 0.05 a | 0.22 ± 0.04 e | |
| MI | 1.27 ± 0.05 de | 0.39 ± 0.02 d | 0.55 ± 0.03 de | 0.66 ± 0.03 ab | 0.33 ± 0.01 d | ||
| Si-NPs | 1.32 ± 0.07 ce | 0.41 ± 0.03 cd | 0.63 ± 0.02 de | 0.59 ± 0.01 b | 0.44 ± 0.01 c | ||
| MI + Si-NPs | 1.35 ± 0.04 bd | 0.56 ± 0.02 c | 0.72 ± 0.06 cd | 0.53 ± 0.03 bc | 0.46 ± 0.03 bc | ||
| Cultivars | * | *** | ** | ** | ** | ||
| Treatments | *** | *** | *** | *** | *** | ||
| Cultivars X Treatments | ns | ** | *** | * | * | ||
| 2021 | Giza 179 | Control | 1.29 ± 0.06 bc | 0.38 ± 0.03 d | 0.54 ± 0.05 de | 0.46 ± 0.02 d | 0.35 ± 0.02 d |
| MI | 1.45 ± 0.08 ab | 0.51 ± 0.04 c | 0.88 ± 0.04 c | 0.28 ± 0.01 e | 0.47 ± 0.02 c | ||
| Si-NPs | 1.48 ± 0.08 ab | 0.89 ± 0.03 b | 1.14 ± 0.08 b | 0.22 ± 0.02 e | 0.52 ± 0.03 b | ||
| MI + Si-NPs | 1.56 ± 0.07 a | 1.08 ± 0.05 a | 1.44 ± 0.08 a | 0.20 ± 0.03 e | 0.59 ± 0.04 a | ||
| Giza 177 | Control | 1.21 ± 0.07 c | 0.15 ± 0.02 e | 0.38 ± 0.06 e | 0.79 ± 0.03 a | 0.26 ± 0.02 e | |
| MI | 1.30 ± 0.06 bc | 0.35 ± 0.05 d | 0.64 ± 0.03 d | 0.63 ± 0.01 b | 0.35 ± 0.01 d | ||
| Si-NPs | 1.32 ± 0.07 bc | 0.41 ± 0.04 d | 0.67 ± 0.02 cd | 0.58 ± 0.01 bc | 0.45 ± 0.01 c | ||
| MI + Si-NPs | 1.35 ± 0.05 ac | 0.55 ± 0.04 c | 0.74 ± 0.06 cd | 0.51 ± 0.02 cd | 0.47 ± 0.02 c | ||
| Cultivars | ** | *** | *** | *** | ** | ||
| Treatments | ** | *** | *** | *** | *** | ||
| Cultivars X Treatments | ns | *** | *** | * | * | ||
| Soluble Cations (meq L−1) | Soluble Anions (meq L−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Season | O.M (%) | EC (dS m−1) | pH | Na+ | K+ | Mg2+ | Ca2+ | Cl− | HCO3− | SO4− |
| 2020 | 1.30 | 7.36 | 8.22 | 64.14 | 0.42 | 8.36 | 10.14 | 59.56 | 8.34 | 7.45 |
| 2021 | 1.32 | 7.24 | 8.12 | 65.03 | 0.40 | 7.24 | 9.98 | 60.32 | 9.26 | 6.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alharbi, K.; Osman, H.S.; Rashwan, E.; Hafez, E.M.; Omara, A.E.-D. Stimulating the Growth, Anabolism, Antioxidants, and Yield of Rice Plants Grown under Salt Stress by Combined Application of Bacterial Inoculants and Nano-Silicon. Plants 2022, 11, 3431. https://doi.org/10.3390/plants11243431
Alharbi K, Osman HS, Rashwan E, Hafez EM, Omara AE-D. Stimulating the Growth, Anabolism, Antioxidants, and Yield of Rice Plants Grown under Salt Stress by Combined Application of Bacterial Inoculants and Nano-Silicon. Plants. 2022; 11(24):3431. https://doi.org/10.3390/plants11243431
Chicago/Turabian StyleAlharbi, Khadiga, Hany S. Osman, Emadeldeen Rashwan, Emad M. Hafez, and Alaa El-Dein Omara. 2022. "Stimulating the Growth, Anabolism, Antioxidants, and Yield of Rice Plants Grown under Salt Stress by Combined Application of Bacterial Inoculants and Nano-Silicon" Plants 11, no. 24: 3431. https://doi.org/10.3390/plants11243431
APA StyleAlharbi, K., Osman, H. S., Rashwan, E., Hafez, E. M., & Omara, A. E.-D. (2022). Stimulating the Growth, Anabolism, Antioxidants, and Yield of Rice Plants Grown under Salt Stress by Combined Application of Bacterial Inoculants and Nano-Silicon. Plants, 11(24), 3431. https://doi.org/10.3390/plants11243431