
Ectomycorrhizal Fungi Modulate Pedunculate Oak’s Heat Stress Responses through the Alternation of Polyamines, Phenolics, and Osmotica Content

 ,
,  ,
,  ,
,  ,
,  , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- Parameters on the physiological level (net rate of photosynthesis, stomatal conductance, and transpiration);
- Accumulation of osmoprotectants such as glycine-betaine and proline;
- Accumulation and profiles of polyamines (putrescine, spermidine, and spermine) as important factors of both abiotic and biotic stress;
- Oxidative stress-related parameters such as the accumulation of important antioxidants (i.e., non-protein thiols, phenolics, flavonoids, and tannins) and the estimation of their total antioxidant and reducing potential by different assays (i.e., ABTS, FRAP) as well as lipid peroxidation (LP) intensity in oak leaves.
2. Results
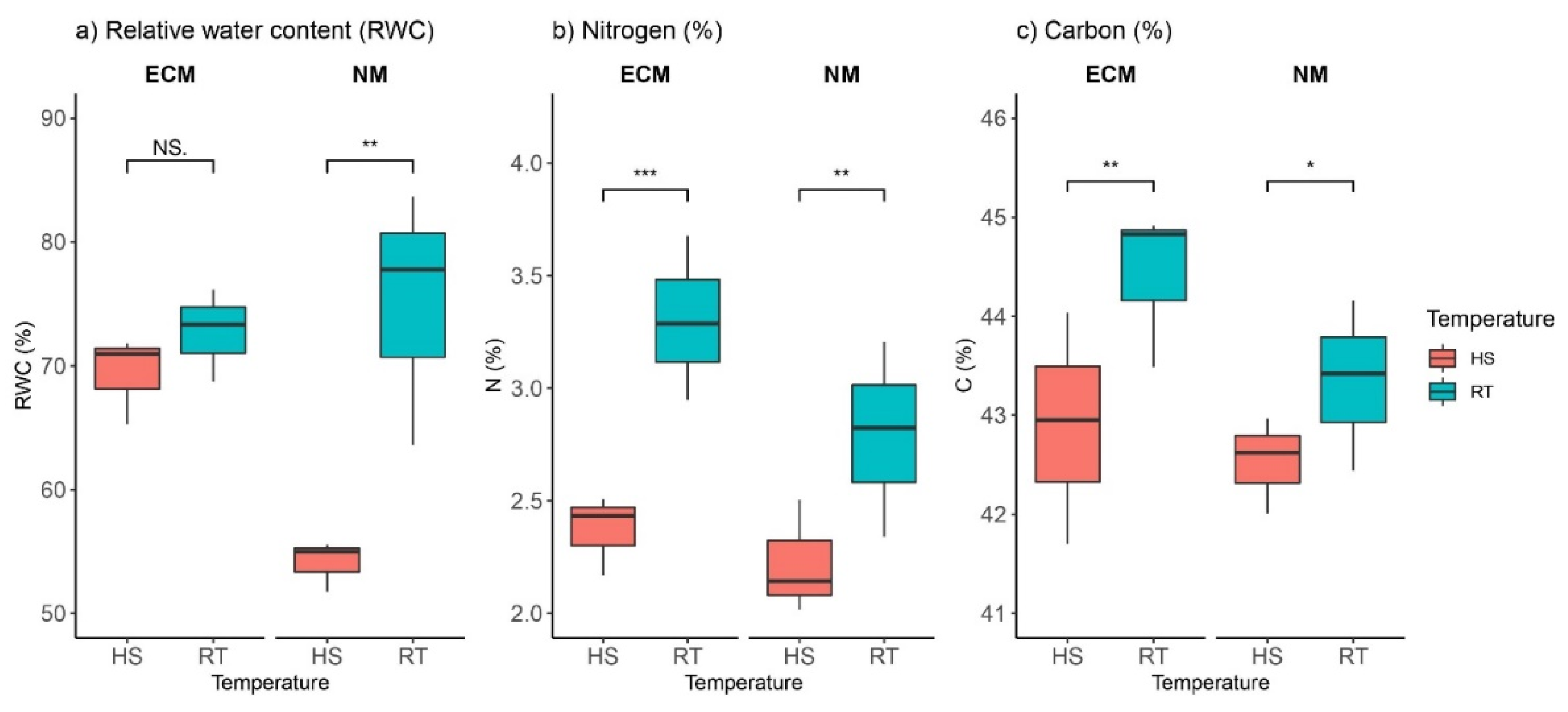
2.1. Heat and ECM Effects on Carbon, Nitrogen, and Leaf-Relative Water Contents
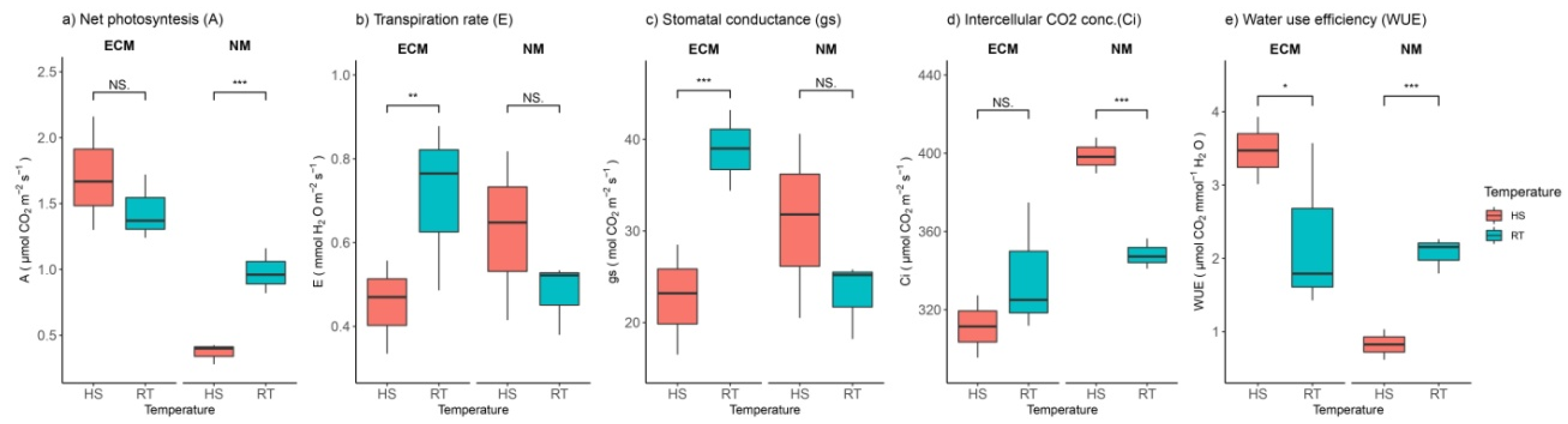
2.2. Heat and ECM Effects on Gas Exchange Parameters
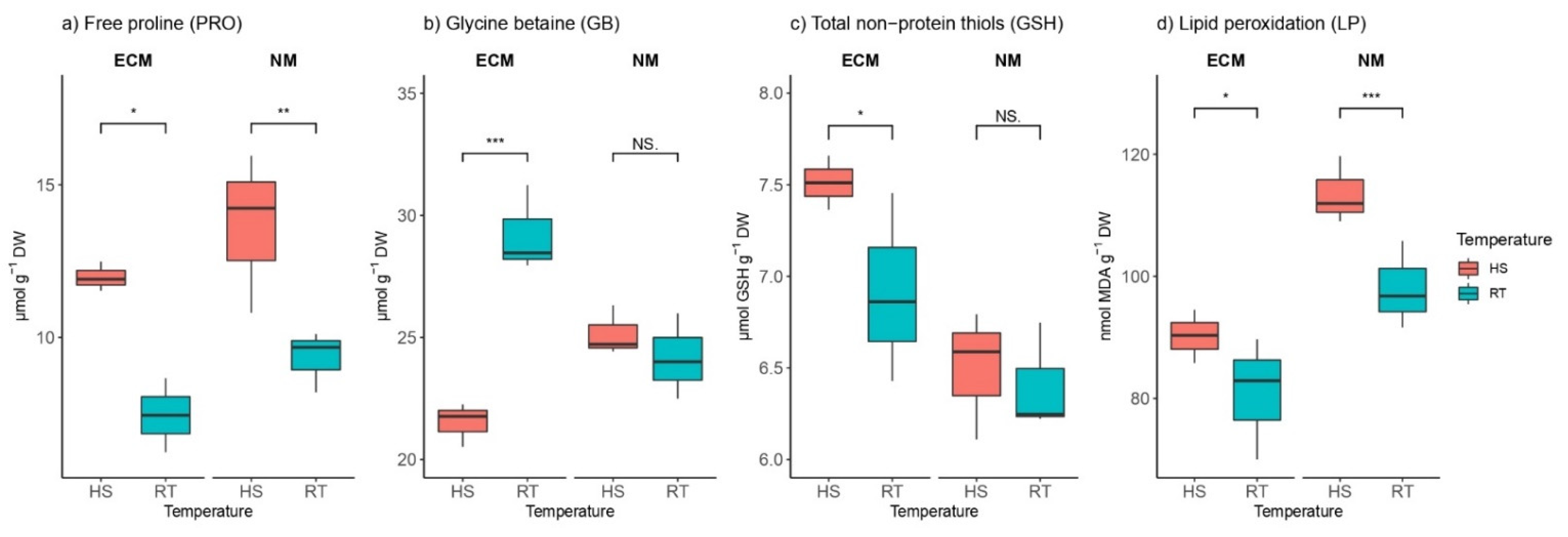
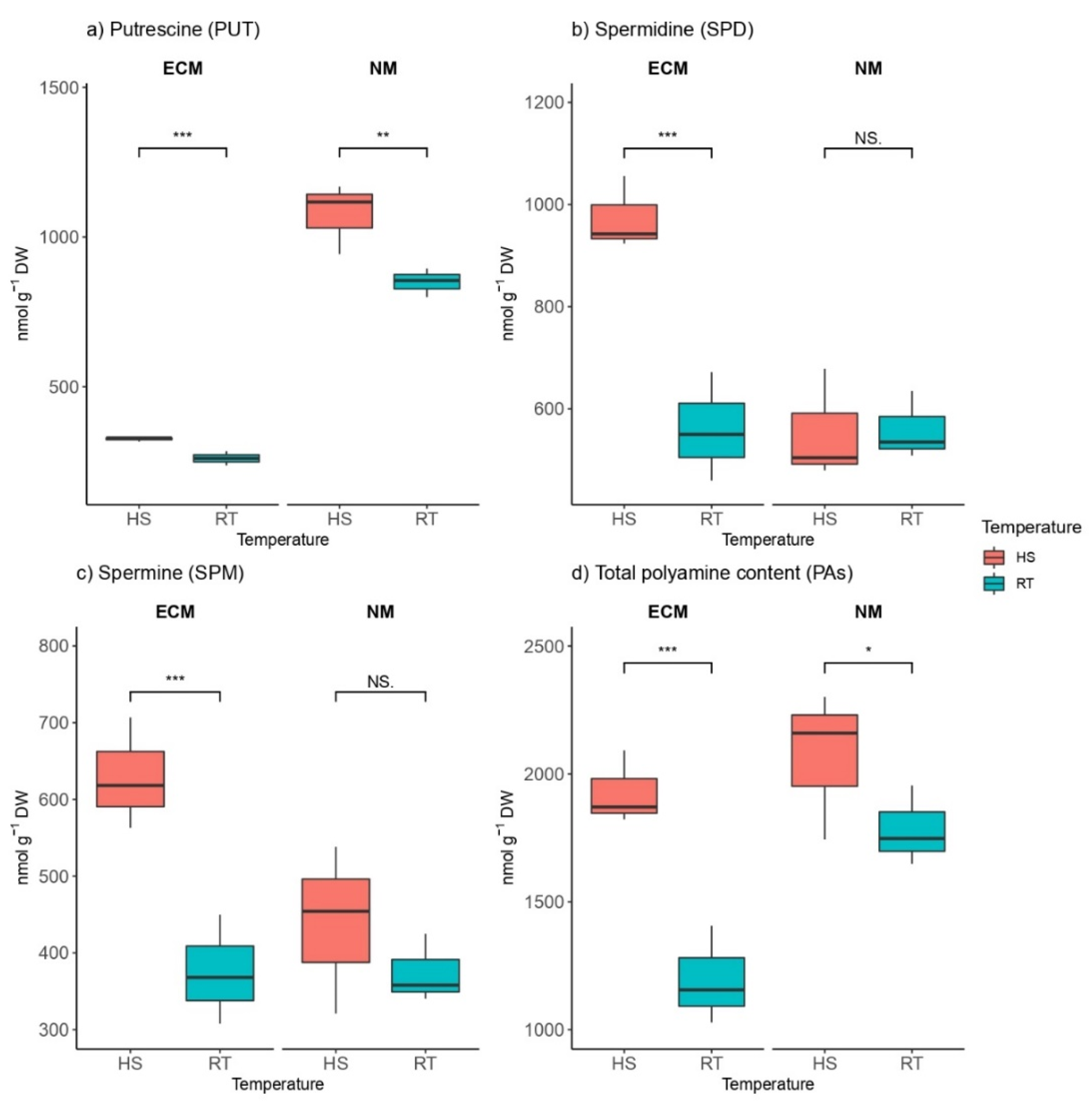
2.3. Heat Effects on Osmolyte Levels in the Presence and Absence of ECM Fungi
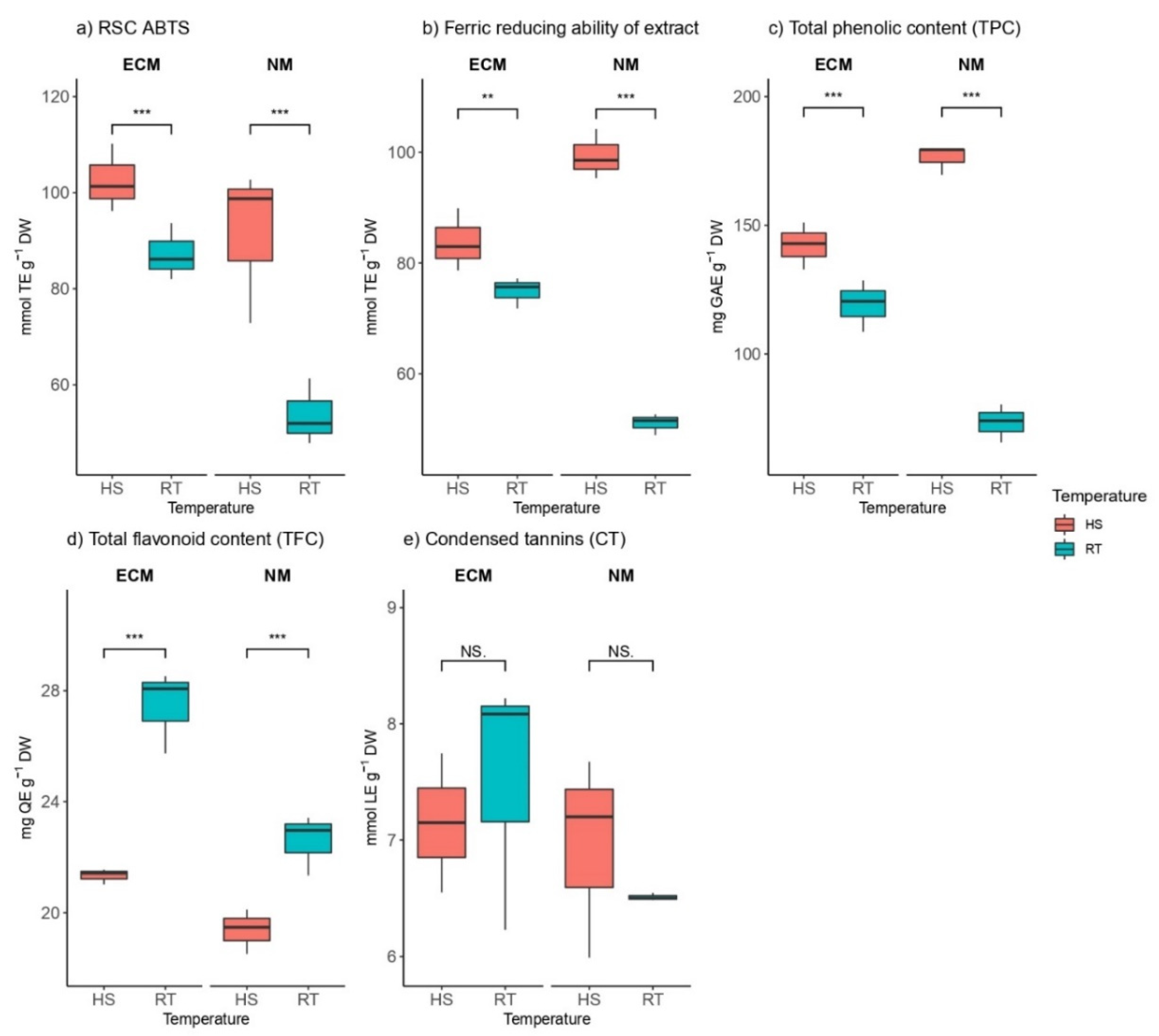
2.4. Changes of Oaks’ Phenolics and Antioxidant Properties under Heat Stress and Mycorrhization
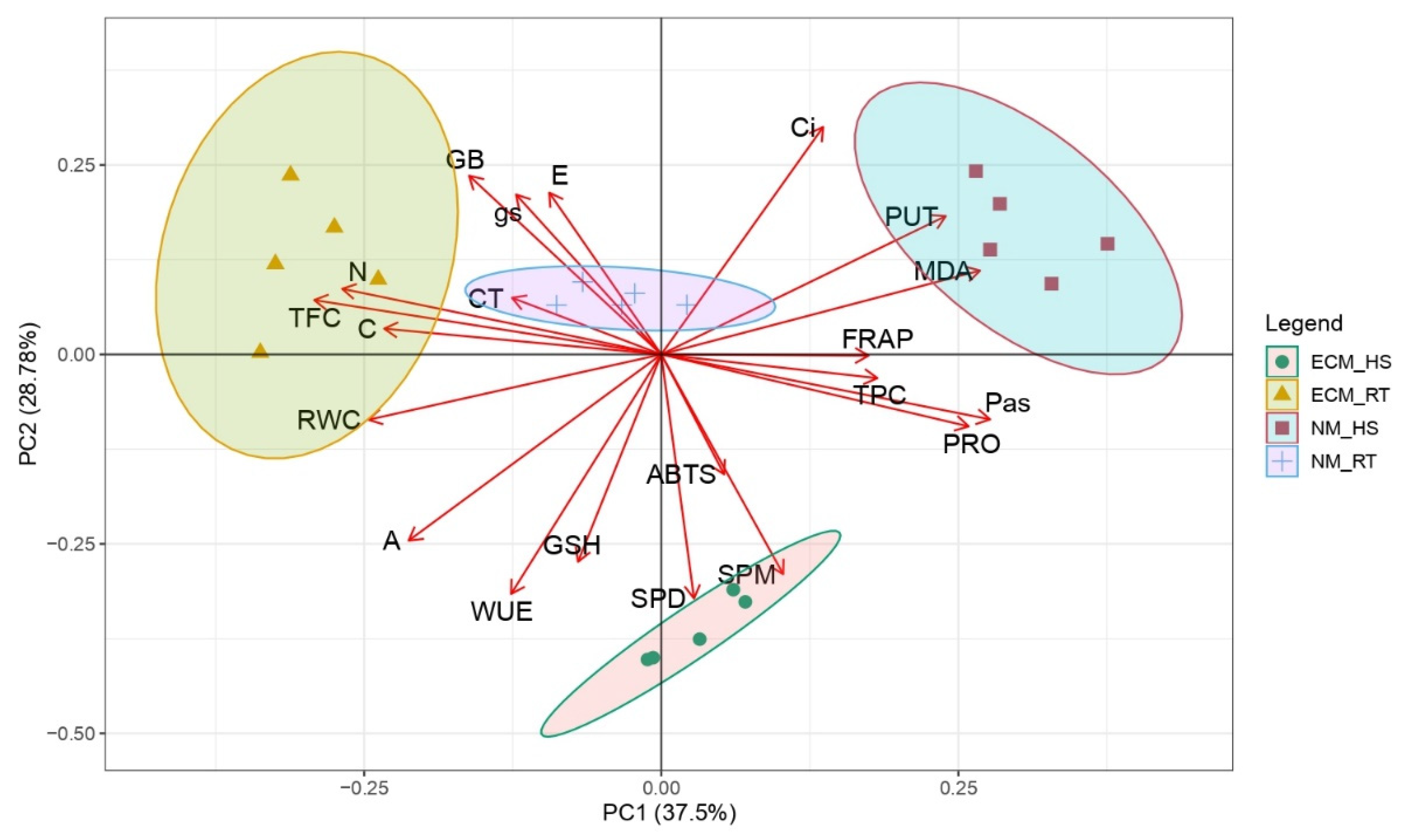
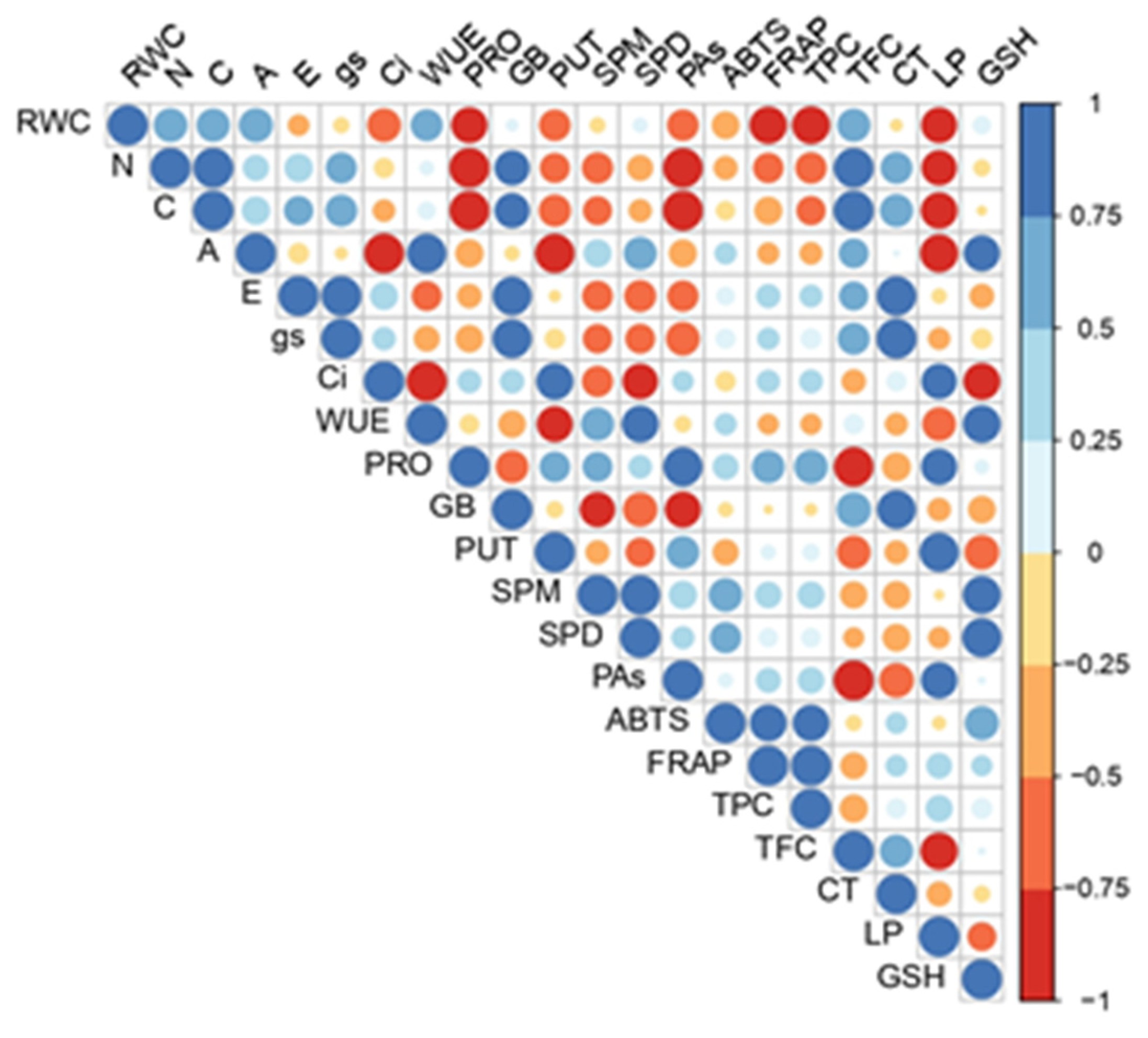
2.5. Principal Component Analysis (PCA) Analysis and Correlation Matrix
3. Discussion
3.1. Perturbations in the Gas Exchange Parameters Caused by Heat and Inoculation with ECM Fungi
3.2. Effects of High-Temperature Stress and Ectomycorrhiza upon Osmolytes
3.3. Alternation of Polyamine Metabolism Induced by Heat and Ectomycorrhiza
3.4. Phenolics and Antioxidant Capacities Modulated by Heat in the Presence and Absence of Ectomycorrhizal Fungi
4. Materials and Methods
4.1. Experimental Design
4.2. Physiological Measurement
4.3. Measurements of Osmolytes’ Accumulation
4.3.1. Polyamine Determination
4.3.2. Assessment of Free Proline
4.3.3. Determination of Glycine Betaine as a Predominant Quaternary Ammonium Compound (QAC)
4.4. Assays of Antioxidant Defense Systems
- Phosphate-buffered saline (PBS; 0.1 M KH2PO4, KOH, pH = 7) extracts were made by vigorously vortexing approximately 0.1 g of freeze-dried plant material with 2 mL of PBS buffer and centrifuging it for 30 min at 13,200 rpm at 4 °C. The glutathione, malondialdehyde (MDA), and FRAP assays were performed on PBS extract supernatants. The antioxidant system response of oak was studied using the following assays: The determination of glutathione (GSH) was carried out colorimetrically after extracts reaction with Ellman’s reagent (5,5-dithio-bis-2-nitrobenzoic acid, DTNS) according to Xue et al. [121].
- ABTS assay based on monitoring of the blue-green colored transformation of the cationic radical 2,2′-azinobis(3-ethylbenzothiozoline-6-sulfonic acid, ABTS.+) into its neutral colorless form at 734 nm was performed according to Miller et al. [122].
- The FRAP (the ferric reducing ability of plasma/extract) test based on a non-specific reaction by which any system that has a less positive redox potential than the ferric 2, 4, 6-tripyridil-S-atrazine complex (Fe3+-TPTZ2]2−/ferric leads to its reduction [123].
- The number of total phenolics was determined according to the extract’s reactivity with Folin–Ciocalteu reagent as previously described [124].
- The total flavonoid content (TFC) was also determined spectrophotometrically by using AlCl3 as flavonoid complexing reagent using the method according to Chang et al. [125].
- The intensity of lipid peroxidation (LP) was determined based on the content of malondialdehyde (MDA), as one of the end-products of lipid peroxidation due to its reactivity with thiobarbituric acid (TBA) [126].
- The condensed tannins (CT) were quantified by applying butanol-HCl-Fe(III) method previously described by Porter et al. [127].
4.5. Elemental Analysis of Inorganic Nitrogen and Carbon
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Tollefson, J. IPCC Says Limiting Global Warming to 1.5 °C Will Require Drastic Action. Nature 2018, 562, 172–173. [Google Scholar] [CrossRef]
- Bouwman, M.; Forrester, D.I.; Den Ouden, J.; Nabuurs, G.J.; Mohren, G.M.J. Species Interactions under Climate Change in Mixed Stands of Scots Pine and Pedunculate Oak. For. Ecol. Manage. 2021, 481, 118615. [Google Scholar] [CrossRef]
- Kostić, S.; Orlović, S.; Karaklić, V.; Kesić, L.; Zorić, M.; Stojanović, D.B. Allometry and Post-Drought Growth Resilience of Pedunculate Oak (Quercus robur L.) Varieties. Forests 2021, 12, 930. [Google Scholar] [CrossRef]
- Stojanović, D.B.; Orlović, S.; Zlatković, M.; Kostić, S.; Vasić, V.; Miletić, B.; Kesić, L.; Matović, B.; Božanić, D.; Pavlović, L.; et al. Climate Change within Serbian Forests: Current State and Future Perspectives. Topola 2021, 208, 39–56. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global Convergence in the Vulnerability of Forests to Drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Rađević, V.; Pap, P.; Vasić, V. Management of the Common Oak Forests in Ravni Srem: Yesterday, Today, Tomorrow. Topola 2020, 206, 41–52. [Google Scholar] [CrossRef]
- Saidi, Y.; Finka, A.; Goloubinoff, P. Heat Perception and Signalling in Plants: A Tortuous Path to Thermotolerance. New Phytol. 2011, 190, 556–565. [Google Scholar] [CrossRef]
- Húdoková, H.; Petrik, P.; Petek-Petrik, A.; Konôpková, A.; Leštianska, A.; Střelcová, K.; Kmet’, J.; Kurjak, D. Heat-Stress Response of Photosystem II in Five Ecologically Important Tree Species of European Temperate Forests. Biologia 2022, 77, 671–680. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and Biotic Stress Combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M. Heat Tolerance in Plants: An Overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic Stress, the Field Environment and Stress Combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Suzuki, N.; Mittler, R. Reactive Oxygen Species and Temperature Stresses: A Delicate Balance between Signaling and Destruction. Physiol. Plant 2006, 126, 45–51. [Google Scholar] [CrossRef]
- Santisree, P.; Bhatnagar-Mathur, P.; Sharma, K.K. NO to Drought-Multifunctional Role of Nitric Oxide in Plant Drought: Do We Have All the Answers? Plant Sci. 2015, 239, 44–55. [Google Scholar] [CrossRef]
- Ye, J.; Zhong, T.; Yu, D.; Sun, S. Physiological and Molecular Biology of High Temperature Stress in Plants. Ann. Agric. Crop Sci. 2021, 6, 1072. [Google Scholar]
- Prasad, P.V.V.; Bheemanahalli, R.; Jagadish, S.V.K. Field Crops and the Fear of Heat Stress—Opportunities, Challenges and Future Directions. Field Crops Res. 2017, 200, 114–121. [Google Scholar] [CrossRef]
- Kebert, M.; Vuksanović, V.; Stefels, J.; Bojović, M.; Horák, R.; Kostić, S.; Kovačević, B.; Orlović, S.; Neri, L.; Magli, M.; et al. Species-Level Differences in Osmoprotectants and Antioxidants Contribute to Stress Tolerance of Quercus robur L., and Q. cerris L. Seedlings under Water Deficit and High Temperatures. Plants 2022, 11, 1744. [Google Scholar] [CrossRef]
- Nadeem, H.; Khan, A.; Gupta, R.; Hashem, M.; Alamri, S.; Siddiqui, M.A.; Ahmad, F. Stress Combination—When Two Negatives May Become Antagonistic, Synergistic, or Additive for Plants: A Review. Pedosphere 2022, in press. [Google Scholar] [CrossRef]
- Alabouvette, C.; Olivain, C.; Steinberg, C. Biological Control of Plant Diseases: The European Situation. Eur. J. Plant Pathol. 2006, 114, 329–341. [Google Scholar] [CrossRef]
- Zaidi, N.W.; Dar, M.H.; Singh, S.; Singh, U.S. Chapter 38—Trichoderma Species as Abiotic Stress Relievers in Plants. In Biotechnology and Biology of Trichoderma; Gupta, V.K., Schmoll, M., Herrera-Estrella, A., Upadhyay, R.S., Druzhinina, I., Tuohy, M.G., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 515–525. [Google Scholar]
- Ahluwalia, O.; Singh, P.C.; Bhatia, R. A Review on Drought Stress in Plants: Implications, Mitigation and the Role of Plant Growth Promoting Rhizobacteria. Res. Environ. Sust. 2021, 5, 100032. [Google Scholar] [CrossRef]
- Bhat, M.A.; Kumar, V.; Bhat, M.A.; Wani, I.A.; Dar, F.L.; Farooq, I.; Bhatti, F.; Koser, R.; Rahman, S.; Jan, A.T. Mechanistic Insights of the Interaction of Plant Growth-Promoting Rhizobacteria (PGPR) With Plant Roots Toward Enhancing Plant Productivity by Alleviating Salinity Stress. Fron. Microbiol. 2020, 11, 1952. [Google Scholar] [CrossRef]
- Jung, S.C.; Martinez-Medina, A.; Lopez-Raez, J.A.; Pozo, M.J. Mycorrhiza-Induced Resistance and Priming of Plant Defenses. J. Chem. Ecol. 2012, 38, 651–664. [Google Scholar] [CrossRef]
- Kebert, M.; Kostić, S.; Zlatković, M.; Stojnić, S.; Čapelja, E.; Zorić, M.; Kiprovski, B.; Budakov, D.; Orlović, S. Ectomycorrhizal Fungi Modulate Biochemical Response against Powdery Mildew Disease in Quercus robur L. Forests 2022, 13, 1491. [Google Scholar] [CrossRef]
- Kivlin, S.N.; Emery, S.M.; Rudgers, J.A. Fungal Symbionts Alter Plant Responses to Global Change. Am. J. Bot. 2013, 100, 1445–1457. [Google Scholar] [CrossRef]
- Pischl, P.H.; Barber, N.A. Plant Responses to Arbuscular Mycorrhizae under Elevated Temperature and Drought. J. Plant Ecol. 2017, 10, 692–701. [Google Scholar] [CrossRef]
- Simard, S.; Austin, M. The Role of Mycorrhizas in Forest Soil Stability with Climate Change. In Climate Change and Variability; Simard, S., Ed.; IntechOpen: London, UK, 2010; pp. 275–302. [Google Scholar]
- Milović, M.; Kebert, M.; Orlović, S. How Mycorrhizas Can Help Forests to Cope with Ongoing Climate Change? Sumar. List 2021, 145, 279–286. [Google Scholar] [CrossRef]
- Malhi, G.S.; Kaur, M.; Kaushik, P.; Alyemeni, M.N.; Alsahli, A.A.; Ahmad, P. Arbuscular Mycorrhiza in Combating Abiotic Stresses in Vegetables: An Eco-Friendly Approach. Saudi J. Biol. Sci. 2021, 28, 1465–1476. [Google Scholar] [CrossRef] [PubMed]
- Nahuelcura, J.; Ruiz, A.; Gomez, F.; Cornejo, P. The Effect of Arbuscular Mycorrhizal Fungi on the Phenolic Compounds Profile, Antioxidant Activity and Grain Yields in Wheat Cultivars Growing under Hydric Stress. J. Sci. Food Agric. 2022, 102, 407–416. [Google Scholar] [CrossRef]
- Santander, C.; Ruiz, A.; García, S.; Aroca, R.; Cumming, J.; Cornejo, P. Efficiency of Two Arbuscular Mycorrhizal Fungal Inocula to Improve Saline Stress Tolerance in Lettuce Plants by Changes of Antioxidant Defense Mechanisms. J. Sci. Food Agric. 2020, 100, 1577–1587. [Google Scholar] [CrossRef]
- Kosonen, M.; Keski-Saari, S.; Ruuhola, T.; Constabel, C.P.; Julkunen-Tiitto, R. Effects of Overproduction of Condensed Tannins and Elevated Temperature on Chemical and Ecological Traits of Genetically Modified Hybrid Aspens (Populus tremula × P. tremuloides). J. Chem. Ecol. 2012, 38, 1235–1246. [Google Scholar] [CrossRef]
- Endo, I.; Kobatake, M.; Tanikawa, N.; Nakaji, T.; Ohashi, M.; Makita, N. Anatomical Patterns of Condensed Tannin in Fine Roots of Tree Species from a Cool-Temperate Forest. Ann. Bot. 2021, 128, 59–71. [Google Scholar] [CrossRef]
- Velikova, V.B.; Edreva, A.M.; Tsonev, T.D.; Jones, H.G. Singlet Oxygen Quenching by Phenylamides and Their Parent Compounds. Z. Nat. C 2007, 62, 833–838. [Google Scholar] [CrossRef]
- Sharma, K.; Gupta, S.; Thokchom, S.D.; Jangir, P.; Kapoor, R. Arbuscular Mycorrhiza-Mediated Regulation of Polyamines and Aquaporins During Abiotic Stress: Deep Insights on the Recondite Players. Front. Plant Sci. 2021, 12, 642101. [Google Scholar] [CrossRef]
- Zou, Y.-N.; Zhang, F.; Srivastava, A.K.; Wu, Q.-S.; Kuča, K. Arbuscular Mycorrhizal Fungi Regulate Polyamine Homeostasis in Roots of Trifoliate Orange for Improved Adaptation to Soil Moisture Deficit Stress. Front. Plant Sci. 2021, 11, 600792. [Google Scholar] [CrossRef]
- Cicatelli, A.; Lingua, G.; Todeschini, V.; Biondi, S.; Torrigiani, P.; Castiglione, S. Arbuscular Mycorrhizal Fungi Restore Normal Growth in a White Poplar Clone Grown on Heavy Metal-Contaminated Soil, and This Is Associated with Upregulation of Foliar Metallothionein and Polyamine Biosynthetic Gene Expression. Ann. Bot. 2010, 106, 791–802. [Google Scholar] [CrossRef]
- Minocha, R.; Majumdar, R.; Minocha, S.C. Polyamines and Abiotic Stress in Plants: A Complex Relationship. Front. Plant Sci. 2014, 5, 175. [Google Scholar] [CrossRef]
- Kebert, M.; Rapparini, F.; Neri, L.; Bertazza, G.; Orlović, S.; Biondi, S. Copper-Induced Responses in Poplar Clones Are Associated with Genotype- and Organ-Specific Changes in Peroxidase Activity and Proline, Polyamine, ABA, and IAA Levels. J. Plant Growth Regul. 2017, 36, 131–147. [Google Scholar] [CrossRef]
- Tun, N.N.; Santa-Catarina, C.; Begum, T.; Silveira, V.; Handro, W.; Floh, E.I.S.; Scherer, G.F.E. Polyamines Induce Rapid Biosynthesis of Nitric Oxide (NO) in Arabidopsis Thaliana Seedlings. Plant Cell Physiol. 2006, 47, 346–354. [Google Scholar] [CrossRef]
- Martínez-Medina, A.; Pescador, L.; Fernández, I.; Rodríguez-Serrano, M.; García, J.M.; Romero-Puertas, M.C.; Pozo, M.J. Nitric Oxide and Phytoglobin PHYTOGB1 Are Regulatory Elements in the Solanum Lycopersicum–Rhizophagus Irregularis Mycorrhizal Symbiosis. New Phytol. 2019, 223, 1560–1574. [Google Scholar] [CrossRef]
- Parankusam, S.; Adimulam, S.S.; Bhatnagar-Mathur, P.; Sharma, K.K. Nitric Oxide (NO) in Plant Heat Stress Tolerance: Current Knowledge and Perspectives. Front. Plant Sci. 2017, 8, 1582. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vicente, I.; Lorenzo, O. Nitric Oxide Regulation of Temperature Acclimation: A Molecular Genetic Perspective. J. Exp. Bot. 2021, 72, 5789–5794. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Sehar, Z.; Fatma, M.; Umar, S.; Sofo, A.; Khan, N.A. Nitric Oxide and Abscisic Acid Mediate Heat Stress Tolerance through Regulation of Osmolytes and Antioxidants to Protect Photosynthesis and Growth in Wheat Plants. Antioxidants 2022, 11, 372. [Google Scholar] [CrossRef]
- Shamshiri, M.H.; Fattahi, M. Evaluation of Two Biochemical Markers for Salt Stress in Three Pistachio Rootstocks Inoculated with Arbuscular Mzycorrhiza (Glomus mosseae). J. Stress Physiol. Biochem. 2014, 10, 335–346. [Google Scholar]
- Wu, H.-H.; Zou, Y.-N.; Rahman, M.M.; Ni, Q.-D.; Wu, Q.-S. Mycorrhizas Alter Sucrose and Proline Metabolism in Trifoliate Orange Exposed to Drought Stress. Sci. Rep. 2017, 7, 42389. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Park, E.-J.; Jeknic, Z.; Chen, T.H.H. Exogenous Application of Glycinebetaine Increases Chilling Tolerance in Tomato Plants. Plant Cell Physiol. 2006, 47, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A.; Steppe, K. Responses of Tree Species to Heat Waves and Extreme Heat Events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- Ruehr, N.K.; Gast, A.; Weber, C.; Daub, B.; Arneth, A. Water Availability as Dominant Control of Heat Stress Responses in Two Contrasting Tree Species. Tree Physiol. 2016, 36, 164–178. [Google Scholar] [CrossRef]
- Birami, B.; Gattmann, M.; Heyer, A.G.; Grote, R.; Arneth, A.; Ruehr, N.K. Heat Waves Alter Carbon Allocation and Increase Mortality of Aleppo Pine Under Dry Conditions. Front. For. Glob. Change 2018, 1, 8. [Google Scholar] [CrossRef]
- Dixon, R.K.; Hiol-Hiol, F. Gas Exchange and Photosynthesis of Eucalyptus Camaldulensis Seedlings Inoculated with Different Ectomycorrhizal Symbionts. Plant Soil 1992, 147, 143–149. [Google Scholar] [CrossRef]
- Hachani, C.; Lamhamedi, M.S.; Zine El Abidine, A.; Abassi, M.; Khasa, D.P.; Béjaoui, Z. Water Relations, Gas Exchange, Chlorophyll Fluorescence and Electrolyte Leakage of Ectomycorrhizal Pinus halepensis Seedlings in Response to Multi-Heavy Metal Stresses (Pb, Zn, Cd). Microorganisms 2021, 10, 57. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Zwiazek, J.; Voicu, M. Responses of Ectomycorrhizal American Elm (Ulmus americana) Seedlings to Salinity and Soil Compaction. Plant Soil 2008, 308, 189–200. [Google Scholar] [CrossRef]
- Zhu, X.-C.; Song, F.-B.; Liu, S.-Q.; Liu, T.-D. Effects of Arbuscular Mycorrhizal Fungus on Photosynthesis and Water Status of Maize under High Temperature Stress. Plant Soil 2011, 346, 189–199. [Google Scholar] [CrossRef]
- Mathur, S.; Sharma, M.P.; Jajoo, A. Improved Photosynthetic Efficacy of Maize (Zea mays) Plants with Arbuscular Mycorrhizal Fungi (AMF) under High Temperature Stress. J. Photochem. Photobiol. B 2018, 180, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Ma, T.; Guo, S.; Liu, R.; Li, M. Leaf Anatomy, Photosynthesis and Chlorophyll Fluorescence of Lettuce as Influenced by Arbuscular Mycorrhizal Fungi under High Temperature Stress. Sci. Hortic. 2021, 280, 109933. [Google Scholar] [CrossRef]
- Kipfer, T.; Wohlgemuth, T.; van der Heijden, M.G.A.; Ghazoul, J.; Egli, S. Growth Response of Drought-Stressed Pinus Sylvestris Seedlings to Single- and Multi-Species Inoculation with Ectomycorrhizal Fungi. PLoS ONE 2012, 7, e35275. [Google Scholar] [CrossRef]
- Martins, A.; Casimiro, A.; Pais, M.S. Influence of Mycorrhization on Physiological Parameters of Micropropagated Castanea Sativa Mill. Plants. Mycorrhiza 1997, 7, 161–165. [Google Scholar] [CrossRef]
- Dosskey, M.G.; Linderman, R.G.; Boersma, L. Carbon-Sink Stimulation of Photosynthesis in Douglas Fir Seedlings by Some Ectomycorrhizas. New Phytol. 1990, 115, 269–274. [Google Scholar] [CrossRef]
- Heinonsalo, J.; Juurola, E.; Linden, A.; Pumpanen, J. Ectomycorrhizal Fungi Affect Scots Pine Photosynthesis through Nitrogen and Water Economy, Not Only through Increased Carbon Demand. Environ. Exp. Bot. 2015, 109, 103–112. [Google Scholar] [CrossRef]
- Dosskey, M.G.; Boersma, L.; Linderman, R.G. Role for the Photosynthate Demand of Ectomycorrhizas in the Response of Douglas Fir Seedlings to Drying Soil. New Phytol. 1991, 117, 327–334. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Chen, H.; Yang, B.; Zhang, F.; Huang, Y. Influence of Arbuscular Mycorrhizae on Photosynthesis and Water Status of Maize Plants under Salt Stress. Mycorrhiza 2008, 18, 287–296. [Google Scholar] [CrossRef]
- Rhodes, D.; Hanson, A.D. Quaternary Ammonium and Tertiary Sulfonium Compounds in Higher Plants. Annu. Rev. Plant Phys. 1993, 44, 357–384. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Pre-Sowing Seed Treatment—A Shotgun Approach to Improve Germination, Plant Growth, and Crop Yield Under Saline and Non-Saline Conditions. Adv. Agr. 2005, 88, 223–271. [Google Scholar]
- Cvikrová, M.; Gemperlová, L.; Martincová, O.; Vanková, R. Effect of Drought and Combined Drought and Heat Stress on Polyamine Metabolism in Proline-over-Producing Tobacco Plants. Plant Physiol. Biochem. 2013, 73, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Fatma, M.; Khan, N.A.; Umar, S. Chapter 28—Regulatory Role of Proline in Heat Stress Tolerance: Modulation by Salicylic Acid. In Plant Signaling Molecules; Khan, M.I.R., Reddy, P.S., Ferrante, A., Khan, N.A., Eds.; Woodhead Publishing: Sawston, UK, 2019; pp. 437–448. [Google Scholar]
- Horak, R.; Župunski, M.; Pajević, S.; Borišev, M.; Arsenov, D.; Nikolić, N.; Orlović, S. Carbon Assimilation in Oak (Quercus spp.) Populations under Acute and Chronic High-Temperature Stress. Photosynthetica 2019, 57, 875–889. [Google Scholar] [CrossRef]
- Yue, Y.; Jiang, H.; Du, J.; Shi, L.; Bin, Q.; Yang, X.; Wang, L. Variations in Physiological Response and Expression Profiles of Proline Metabolism-Related Genes and Heat Shock Transcription Factor Genes in Petunia Subjected to Heat Stress. Sci. Hortic. 2019, 258, 108811. [Google Scholar] [CrossRef]
- Lopez-Delacalle, M.; Silva, C.J.; Mestre, T.C.; Martinez, V.; Blanco-Ulate, B.; Rivero, R.M. Synchronization of Proline, Ascorbate and Oxidative Stress Pathways under the Combination of Salinity and Heat in Tomato Plants. Environ. Exp. Bot. 2021, 183, 104351. [Google Scholar] [CrossRef]
- Alvarez, M.E.; Savouré, A.; Szabados, L. Proline Metabolism as Regulatory Hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar] [CrossRef]
- Alamri, S.A.; Siddiqui, M.H.; Al-Khaishany, M.Y.; Khan, M.N.; Ali, H.M.; Alakeel, K.A. Nitric Oxide-Mediated Cross-Talk of Proline and Heat Shock Proteins Induce Thermotolerance in Vicia faba L. Environ. Exp. Bot. 2019, 161, 290–302. [Google Scholar] [CrossRef]
- Zhang, T.; Hu, Y.; Zhang, K.; Tian, C.; Guo, J. Arbuscular Mycorrhizal Fungi Improve Plant Growth of Ricinus Communis by Altering Photosynthetic Properties and Increasing Pigments under Drought and Salt Stress. Ind. Crops Prod. 2018, 117, 13–19. [Google Scholar] [CrossRef]
- Zhang, Y.-C.; Zou, Y.-N.; Liu, L.-P.; Wu, Q.-S. Common Mycorrhizal Networks Activate Salicylic Acid Defense Responses of Trifoliate Orange (Poncirus trifoliata). J. Integr. Plant Biol. 2019, 61, 1099–1111. [Google Scholar] [CrossRef]
- Al-Arjani, A.-B.F.; Hashem, A.; Abd Allah, E.F. Arbuscular Mycorrhizal Fungi Modulates Dynamics Tolerance Expression to Mitigate Drought Stress in Ephedra Foliata Boiss. Saudi J. Biol. Sci. 2020, 27, 380–394. [Google Scholar] [CrossRef] [PubMed]
- Aslanpour, M.; Doulati Baneh, H.; Tehranifar, A.; Shoor, M. The Effect of Mycorrhizal Fungi on the Amount of Glycine Betaine, Soluble Sugar, Proline, Leaf Water Content and Leaf Chlorophyll of the White Seedless Grape under Drought Stress Conditions. Int. J. Adv. Biotechnol. Res. 2016, 7, 1119–1133. [Google Scholar]
- Karunanantham, K.; Lakshminarayanan, S.P.; Ganesamurthi, A.K.; Ramasamy, K.; Rajamony, V.R. Chapter 10—Arbuscular Mycorrhiza—A Health Engineer for Abiotic Stress Alleviation. In Rhizosphere Engineering; Dubey, R.C., Kumar, P., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 171–198. [Google Scholar]
- Hagh-Doust, N.; Färkkilä, S.M.A.; Hosseyni Moghaddam, M.S.; Tedersoo, L. Symbiotic Fungi as Biotechnological Tools: Methodological Challenges and Relative Benefits in Agriculture and Forestry. Fungal Biol. Rev. 2022, 42, 34–55. [Google Scholar] [CrossRef]
- Yang, X.; Wen, X.; Gong, H.; Lu, Q.; Yang, Z.; Tang, Y.; Liang, Z.; Lu, C. Genetic Engineering of the Biosynthesis of Glycinebetaine Enhances Thermotolerance of Photosystem II in Tobacco Plants. Planta 2007, 225, 719–733. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Teixidó, N.; Cañamás, T.P.; Usall, J.; Torres, R.; Magan, N.; Viñas, I. Accumulation of the Compatible Solutes, Glycine-Betaine and Ectoine, in Osmotic Stress Adaptation and Heat Shock Cross-Protection in the Biocontrol Agent Pantoea Agglomerans CPA-2. Lett. Appl. Microbiol. 2005, 41, 248–252. [Google Scholar] [CrossRef]
- Mäkelä, P.; Kontturi, M.; Pehu, E.; Somersalo, S. Photosynthetic Response of Drought- and Salt-Stressed Tomato and Turnip Rape Plants to Foliar-Applied Glycinebetaine. Physiol. Plant. 1999, 105, 45–50. [Google Scholar] [CrossRef]
- Papageorgiou, G.C.; Murata, N. The Unusually Strong Stabilizing Effects of Glycine Betaine on the Structure and Function of the Oxygen-Evolving Photosystem II Complex. Photosynth. Res. 1995, 44, 243–252. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Xia, R.-X.; Zou, Y.-N.; Wang, G.-Y. Osmotic Solute Responses of Mycorrhizal Citrus (Poncirus trifoliata) Seedlings to Drought Stress. Acta Physiol. Plant 2007, 29, 543–549. [Google Scholar] [CrossRef]
- Carillo, P.; Mastrolonardo, G.; Nacca, F.; Parisi, D.; Verlotta, A.; Fuggi, A. Nitrogen Metabolism in Durum Wheat under Salinity: Accumulation of Proline and Glycine Betaine. Funct. Plant Biol. 2008, 35, 412–426. [Google Scholar] [CrossRef]
- Carillo, P. GABA Shunt in Durum Wheat. Front. Plant Sci. 2018, 9, 100. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Maximova, E.; Fuggi, A.; Carillo, P. Durum Wheat Roots Adapt to Salinity Remodeling the Cellular Content of Nitrogen Metabolites and Sucrose. Front. Plant Sci. 2017, 7, 2035. [Google Scholar] [CrossRef] [PubMed]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and Temporal Profile of Glycine Betaine Accumulation in Plants Under Abiotic Stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef]
- Lingua, G.; Franchin, C.; Todeschini, V.; Castiglione, S.; Biondi, S.; Burlando, B.; Parravicini, V.; Torrigiani, P.; Berta, G. Arbuscular Mycorrhizal Fungi Differentially Affect the Response to High Zinc Concentrations of Two Registered Poplar Clones. Environ. Pollut. 2008, 153, 137–147. [Google Scholar] [CrossRef]
- Majumdar, R.; Shao, L.; Minocha, R.; Long, S.; Minocha, S.C. Ornithine: The Overlooked Molecule in the Regulation of Polyamine Metabolism. Plant Cell Physiol. 2013, 54, 990–1004. [Google Scholar] [CrossRef]
- Navakoudis, E.; Kotzabasis, K. Polyamines: A Bioenergetic Smart Switch for Plant Protection and Development. J. Plant Physiol. 2022, 270, 153618. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Pál, M.; Szalai, G.; Janda, T. Speculation: Polyamines Are Important in Abiotic Stress Signaling. Plant Sci. 2015, 237, 16–23. [Google Scholar] [CrossRef]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small Amines with Large Effects on Plant Abiotic Stress Tolerance. Cells 2020, 9, E2373. [Google Scholar] [CrossRef]
- Sakamoto, A.; Tamakoshi, M.; Moriya, T.; Oshima, T.; Takao, K.; Sugita, Y.; Furuchi, T.; Niitsu, M.; Uemura, T.; Igarashi, K.; et al. Polyamines Produced by an Extreme Thermophile Are Essential for Cell Growth at High Temperature. J. Biochem. 2022, 172, 109–115. [Google Scholar] [CrossRef]
- Garg, N.; Sharma, A. Role of Putrescine (Put) in Imparting Salt Tolerance through Modulation of Put Metabolism, Mycorrhizal and Rhizobial Symbioses in Cajanus cajan (L.) Millsp. Symbiosis 2019, 79, 59–74. [Google Scholar] [CrossRef]
- Garg, N.; Saroy, K. Interactive Effects of Polyamines and Arbuscular Mycorrhiza in Modulating Plant Biomass, N2 Fixation, Ureide, and Trehalose Metabolism in Cajanus cajan (L.) Millsp. Genotypes under Nickel Stress. Environ. Sci. Pollut. Res. Int. 2020, 27, 3043–3064. [Google Scholar] [CrossRef] [PubMed]
- Shaul-Keinan, O.; Gadkar, V.; Ginzberg, I.; Grünzweig, J.M.; Chet, I.; Elad, Y.; Wininger, S.; Belausov, E.; Eshed, Y.; Atzmon, N.; et al. Hormone Concentrations in Tobacco Roots Change during Arbuscular Mycorrhizal Colonization with Glomus Intraradices. New Phytol. 2002, 154, 501–507. [Google Scholar] [CrossRef]
- Khalloufi, M.; Martínez-Andújar, C.; Lachaâl, M.; Karray-Bouraoui, N.; Pérez-Alfocea, F.; Albacete, A. The Interaction between Foliar GA3 Application and Arbuscular Mycorrhizal Fungi Inoculation Improves Growth in Salinized Tomato (Solanum lycopersicum L.) Plants by Modifying the Hormonal Balance. J. Plant Physiol. 2017, 214, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Evelin, H.; Giri, B.; Kapoor, R. Ultrastructural Evidence for AMF Mediated Salt Stress Mitigation in Trigonella Foenum-Graecum. Mycorrhiza 2013, 23, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.-M.; Zheng, F.-L.; Wu, Q.-S. Elucidating the Dialogue between Arbuscular Mycorrhizal Fungi and Polyamines in Plants. World J. Microbiol. Biotechnol. 2022, 38, 159. [Google Scholar] [CrossRef] [PubMed]
- Talaat, N.B.; Shawky, B.T. Modulation of Nutrient Acquisition and Polyamine Pool in Salt-Stressed Wheat (Triticum aestivum L.) Plants Inoculated with Arbuscular Mycorrhizal Fungi. Acta Physiol. Plant 2013, 35, 2601–2610. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, B. Arbuscular Mycorrhiza Induced Putrescine Degradation into γ-Aminobutyric Acid, Malic Acid Accumulation, and Improvement of Nitrogen Assimilation in Roots of Water-Stressed Maize Plants. Mycorrhiza 2020, 30, 329–339. [Google Scholar] [CrossRef]
- Shelp, B.J.; Bozzo, G.G.; Trobacher, C.P.; Zarei, A.; Deyman, K.L.; Brikis, C.J. Hypothesis/Review: Contribution of Putrescine to 4-Aminobutyrate (GABA) Production in Response to Abiotic Stress. Plant Sci. 2012, 193, 130–135. [Google Scholar] [CrossRef]
- Wu, Q.-S.; Ying-Ning, Z.; Liu, C.-Y.; Xiao, J.; Li, Y. Arbuscular Mycorrhizas Alter Root System Architecture of Citrus Tangerine through Regulating Metabolism of Endogenous Polyamines. Plant Growth Regul. 2012, 68, 27–35. [Google Scholar] [CrossRef]
- Sarjala, T.; Niemi, K.; Häggman, H. Mycorrhiza Formation Is Not Needed for Early Growth Induction and Growth-Related Changes in Polyamines in Scots Pine Seedlings in Vitro. Plant Physiol. Biochem. 2010, 48, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Pintó-Marijuan, M.; Munné-Bosch, S. Photo-Oxidative Stress Markers as a Measure of Abiotic Stress-Induced Leaf Senescence: Advantages and Limitations. J. Exp. Bot. 2014, 65, 3845–3857. [Google Scholar] [CrossRef] [PubMed]
- Pospíšil, P. Production of Reactive Oxygen Species by Photosystem II as a Response to Light and Temperature Stress. Front. Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef] [PubMed]
- Petrik, P.; Petek-Petrik, A.; Konôpková, A.; Fleischer, P.; Stojnic, S.; Zavadilova, I.; Kurjak, D. Seasonality of PSII Thermostability and Water Use Efficiency of in Situ Mountainous Norway Spruce (Picea abies). J. For. Res. 2022, 1, 1–12. [Google Scholar] [CrossRef]
- Kaushal, N.; Awasthi, R.; Gupta, K.; Gaur, P.; Siddique, K.H.M.; Nayyar, H. Heat-Stress-Induced Reproductive Failures in Chickpea (Cicer arietinum) Are Associated with Impaired Sucrose Metabolism in Leaves and Anthers. Funct. Plant Biol. 2013, 40, 1334–1349. [Google Scholar] [CrossRef]
- Yadav, R.; Juneja, S.; Kumar, R.; Saini, R.; Kumar, S. Understanding Cross-Tolerance Mechanism and Effect of Drought Priming on Individual Heat Stress and Combinatorial Heat and Drought Stress in Chickpea. J. Crop Sci. Biotechnol. 2022, 25, 515–533. [Google Scholar] [CrossRef]
- Hassan, F.A.S.; Ali, E.; Gaber, A.; Fetouh, M.I.; Mazrou, R. Chitosan Nanoparticles Effectively Combat Salinity Stress by Enhancing Antioxidant Activity and Alkaloid Biosynthesis in Catharanthus roseus (L.) G. Don. Plant Physiol. Biochem. 2021, 162, 291–300. [Google Scholar] [CrossRef]
- Salvucci, M.; Crafts-Brandner, S. Inhibition of Photosynthesis by Heat Stress: The Activation State of RuBisCO as a Limiting Factor in Photosynthesis. Physiol. Plant. 2004, 120, 179–186. [Google Scholar] [CrossRef]
- Wilson, R.A.; Gupta, S.; Sangha, M.K.; Kaur, G. Effect of Heat Stress on Enzymatic and Non-Enzymatic Antioxidants in Brassica Rapa. J. Environ. Biol. 2019, 40, 119–124. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Xylia, P.; Antoniou, O.; Tzortzakis, N. Climate Change Due to Heat and Drought Stress Can Alter the Physiology of Maratheftiko Local Cyprian Grapevine Variety. J. Water Clim. Change 2018, 9, 715–727. [Google Scholar] [CrossRef]
- Tommasino, E.; López Colomba, E.; Carrizo, M.; Grunberg, K.; Quiroga, M.; Carloni, E.; Griffa, S.; Ribotta, A.; Luna, C. Individual and Combined Effects of Drought and Heat on Antioxidant Parameters and Growth Performance in Buffel Grass (Cenchrus ciliaris L.) Genotypes. S. Afr. J. Bot. 2018, 119, 104–111. [Google Scholar] [CrossRef]
- Moura, D.S.; Brito, G.G.; Campos, Â.D.; Moraes, Í.L.; Fagundes, P.R.R.; Deuner, S. Phenylalanine Ammonia-Lyase and Source-Flow-Sink Related Attributes in Rice Genotypes Subjected to High Night Temperatures. J. Agric. Sci. 2017, 9, 268–282. [Google Scholar] [CrossRef][Green Version]
- Zhou, R.; Yu, X.; Kjær, K.H.; Rosenqvist, E.; Ottosen, C.-O.; Wu, Z. Screening and Validation of Tomato Genotypes under Heat Stress Using Fv/Fm to Reveal the Physiological Mechanism of Heat Tolerance. Environ. Exp. Bot. 2015, 118, 1–11. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Scaramagli, S.; Biondi, S.; Capitani, F.; Gerola, P.; Altamura, M.M.; Torrigiani, P. Polyamine Conjugate Levels and Ethylene Biosynthesis: Inverse Relationship with Vegetative Bud Formation in Tobacco Thin Layers. Physiol. Plant. 1999, 105, 366–375. [Google Scholar] [CrossRef]
- Grieve, C.M.; Grattan, S.R. Rapid Assay for Determination of Water Soluble Quaternary Ammonium Compounds. Plant Soil 1983, 70, 303–307. [Google Scholar] [CrossRef]
- Xue, T.; Hartikainen, H.; Piironen, V. Antioxidative and Growth-Promoting Effect of Selenium on Senescing Lettuce. Plant Soil 2001, 237, 55–61. [Google Scholar] [CrossRef]
- Miller, N.J.; Rice-Evans, C.A. Factors Influencing the Antioxidant Activity Determined by the ABTS. + Radical Cation Assay. Free. Radic. Res. 1997, 26, 195–199. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Kim, D.-O.; Jeong, S.W.; Lee, C.Y. Antioxidant Capacity of Phenolic Phytochemicals from Various Cultivars of Plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Chang, C.-C.; Yang, M.-H.; Wen, H.-M.; Chern, J.-C. Estimation of Total Flavonoid Content in Propolis by Two Complementary Colometric Methods. J. Food Drug Anal. 2002, 10, 3. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the Thiobarbituric Acid-Reactive-Substances Assay for Estimating Lipid Peroxidation in Plant Tissues Containing Anthocyanin and Other Interfering Compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Porter, L.J.; Hrstich, L.N.; Chan, B.G. The Conversion of Procyanidins and Prodelphinidins to Cyanidin and Delphinidin. Phytochemistry 1985, 25, 223–230. [Google Scholar] [CrossRef]
- Karthikeyan, R.; Kumaravel, S. Study on Phenolic Content, Antioxidant Activity and CHNS Elemental Analysis of Amorphophallus Sylvaticus. Int. J. Agric. Life Sci. 2016, 2, 12–17. [Google Scholar]
- Kassambara, A. Pipe-Friendly Framework for Basic Statistical Tests [R Package Rstatix, Version 0.7.0]; Free Software Foundation Inc.: Boston, MA, USA, 2021. [Google Scholar]
- Wickham, H. Ggplot2. WIREs Comput. Stat. 2011, 3, 180–185. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kebert, M.; Kostić, S.; Čapelja, E.; Vuksanović, V.; Stojnić, S.; Markić, A.G.; Zlatković, M.; Milović, M.; Galović, V.; Orlović, S. Ectomycorrhizal Fungi Modulate Pedunculate Oak’s Heat Stress Responses through the Alternation of Polyamines, Phenolics, and Osmotica Content. Plants 2022, 11, 3360. https://doi.org/10.3390/plants11233360
Kebert M, Kostić S, Čapelja E, Vuksanović V, Stojnić S, Markić AG, Zlatković M, Milović M, Galović V, Orlović S. Ectomycorrhizal Fungi Modulate Pedunculate Oak’s Heat Stress Responses through the Alternation of Polyamines, Phenolics, and Osmotica Content. Plants. 2022; 11(23):3360. https://doi.org/10.3390/plants11233360
Chicago/Turabian StyleKebert, Marko, Saša Kostić, Eleonora Čapelja, Vanja Vuksanović, Srđan Stojnić, Anđelina Gavranović Markić, Milica Zlatković, Marina Milović, Vladislava Galović, and Saša Orlović. 2022. "Ectomycorrhizal Fungi Modulate Pedunculate Oak’s Heat Stress Responses through the Alternation of Polyamines, Phenolics, and Osmotica Content" Plants 11, no. 23: 3360. https://doi.org/10.3390/plants11233360
APA StyleKebert, M., Kostić, S., Čapelja, E., Vuksanović, V., Stojnić, S., Markić, A. G., Zlatković, M., Milović, M., Galović, V., & Orlović, S. (2022). Ectomycorrhizal Fungi Modulate Pedunculate Oak’s Heat Stress Responses through the Alternation of Polyamines, Phenolics, and Osmotica Content. Plants, 11(23), 3360. https://doi.org/10.3390/plants11233360