
GC, GC/MS Analysis, and Biological Effects of Essential Oils from Thymus mastchina and Elettaria cardamomum

 , , , and
, , , and 
Abstract
1. Introduction
2. Results
2.1. Volatile Composition of T. mastichina Essential Oil
2.2. Volatile Composition of E. cardamomum Essential Oil
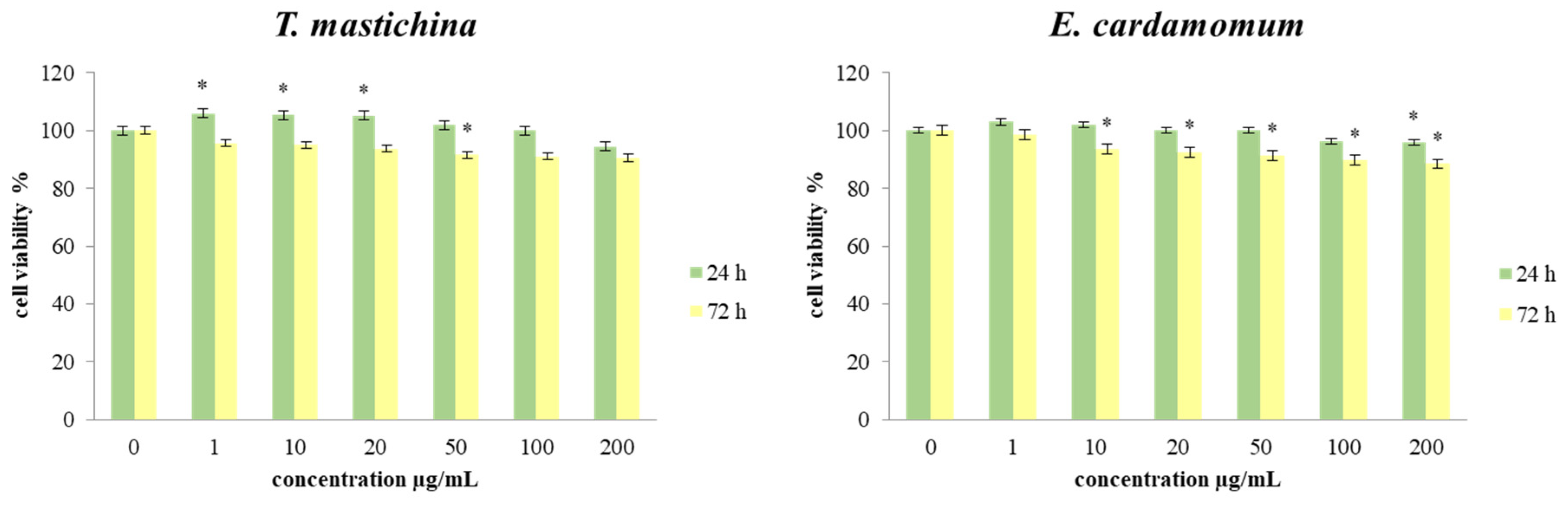
2.3. Cell Viability Assays
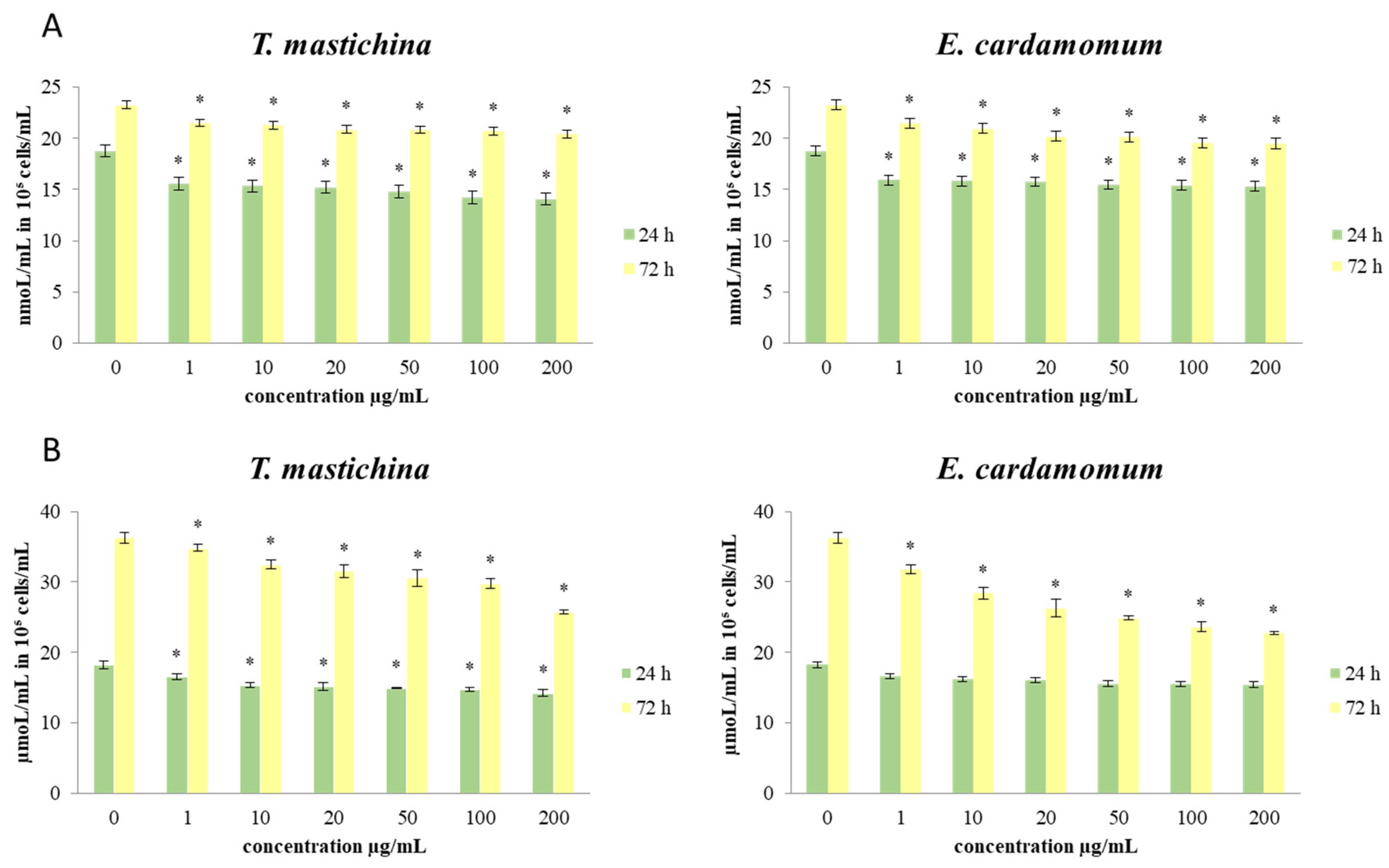
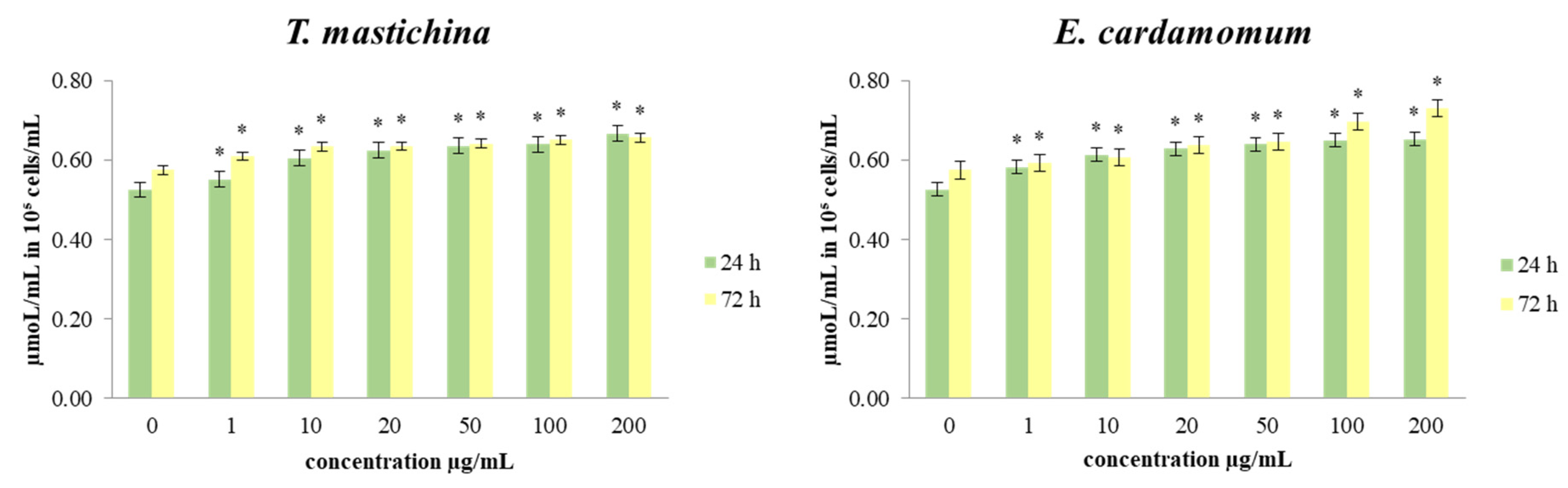
2.4. The Effects of Essential Oils on Redox Status Parameters
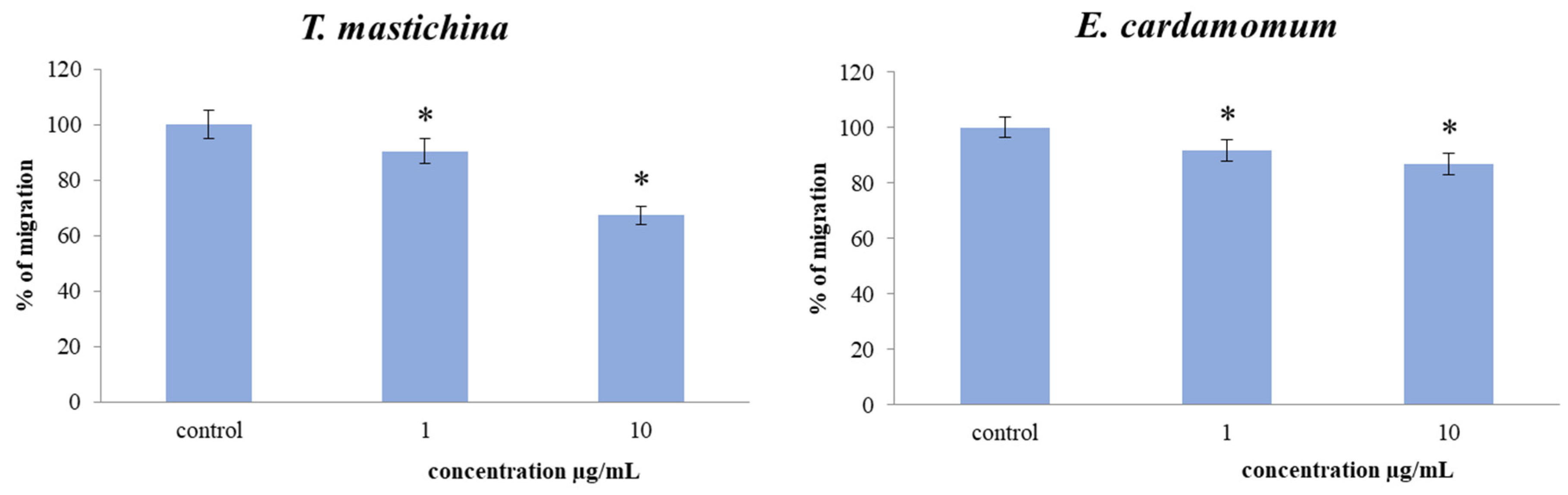
2.5. Transwell Assay for Cell Migration
2.6. Antimicrobial Testing
2.7. Antibiofilm Testing
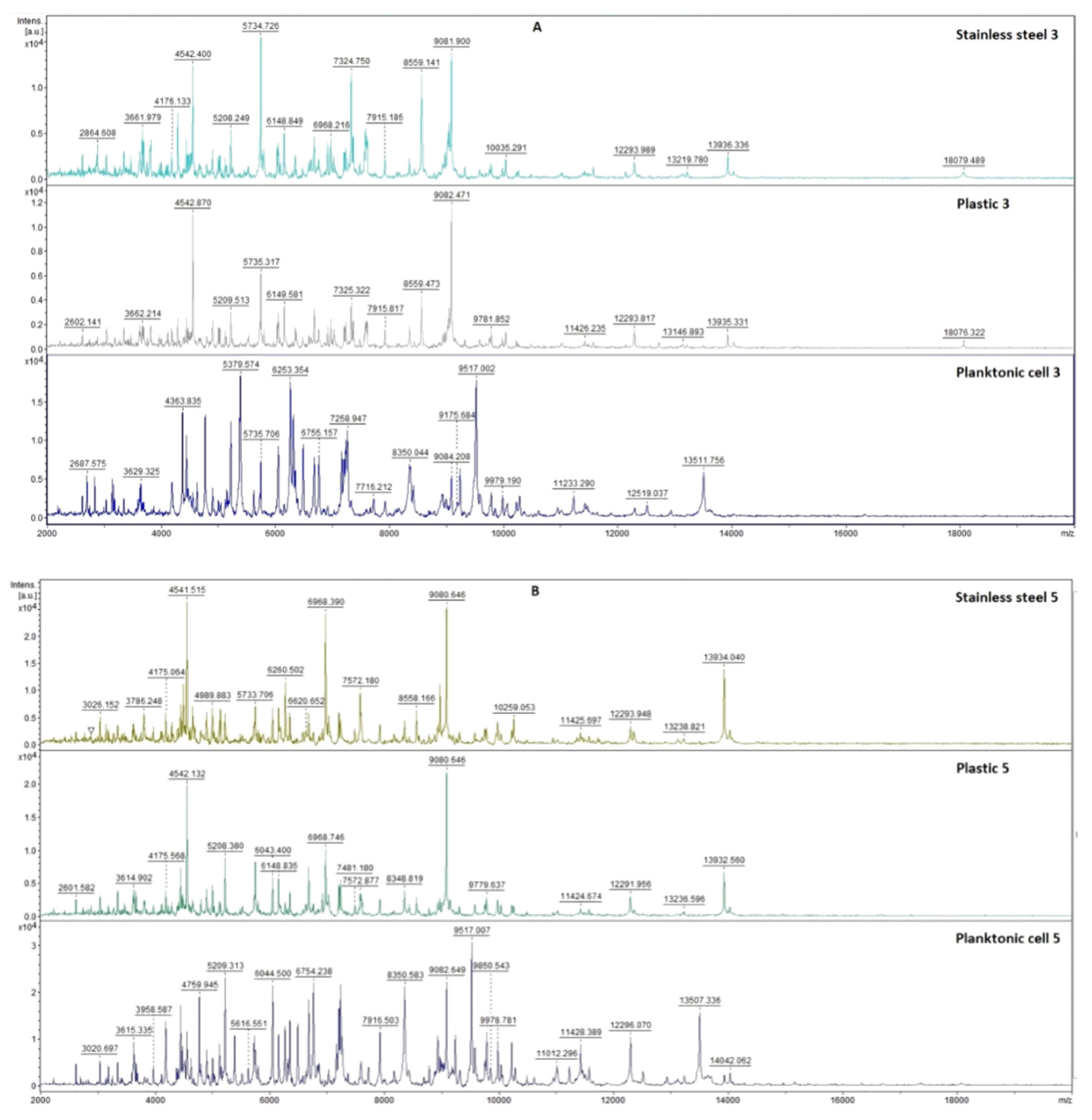
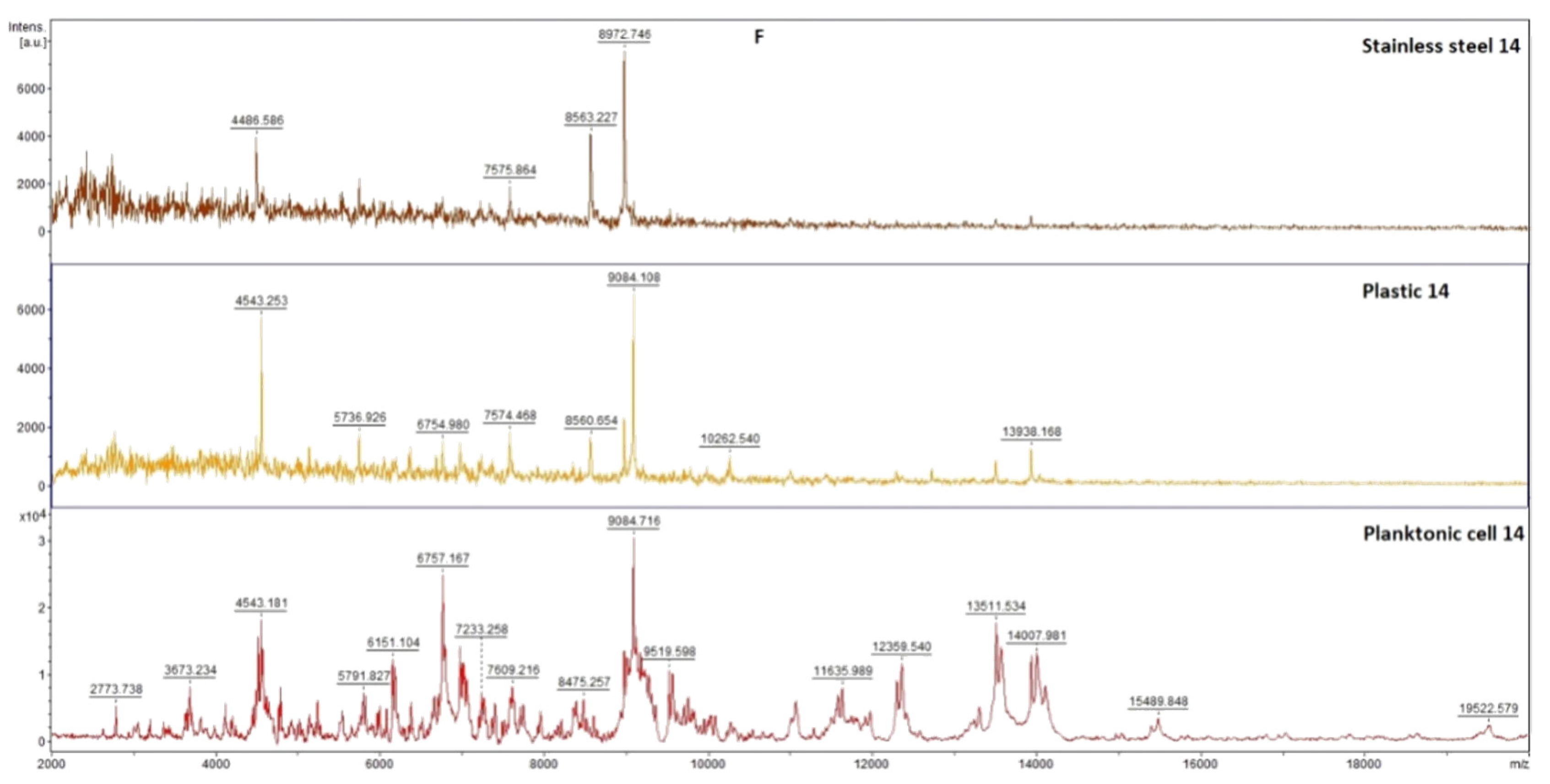
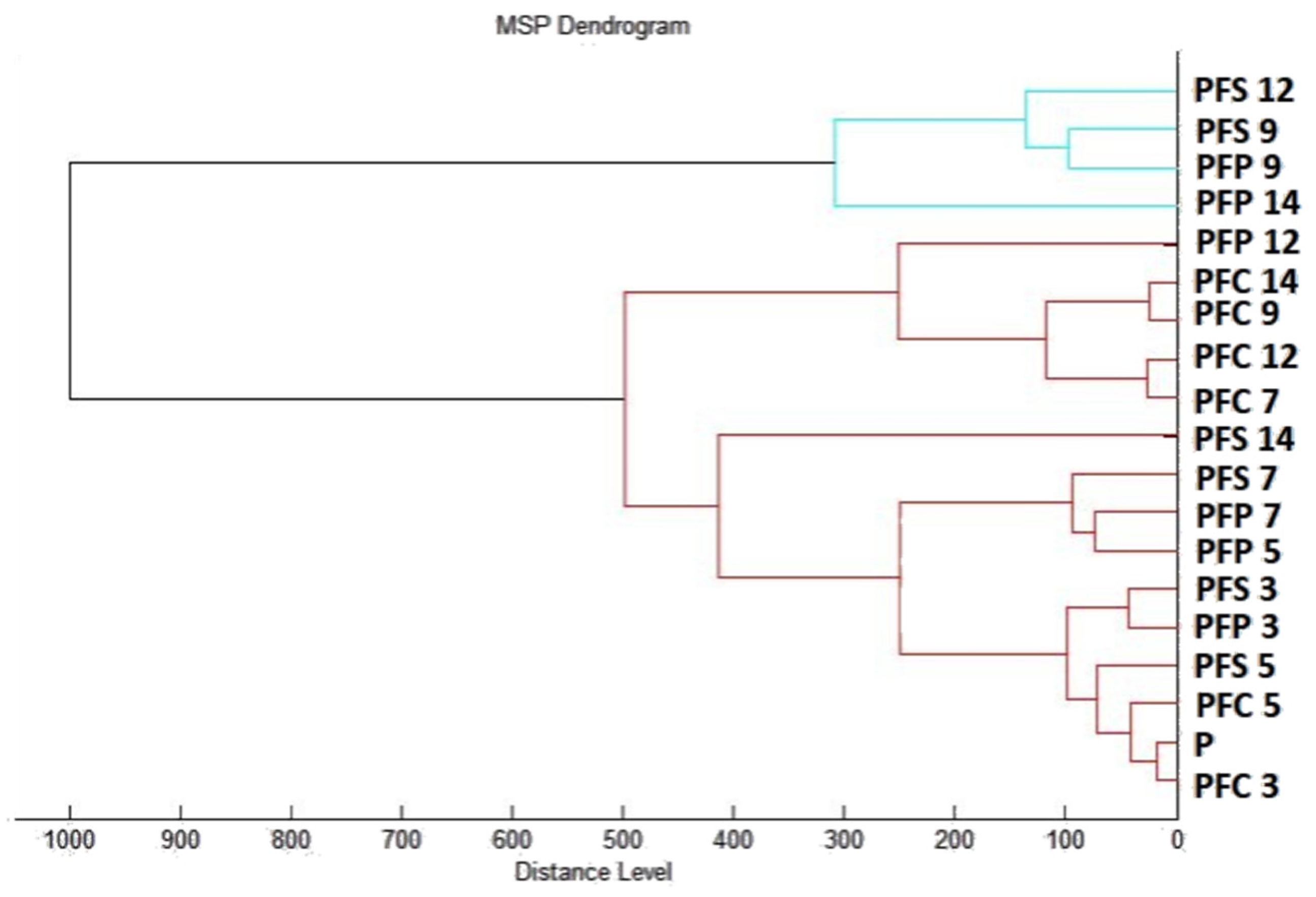
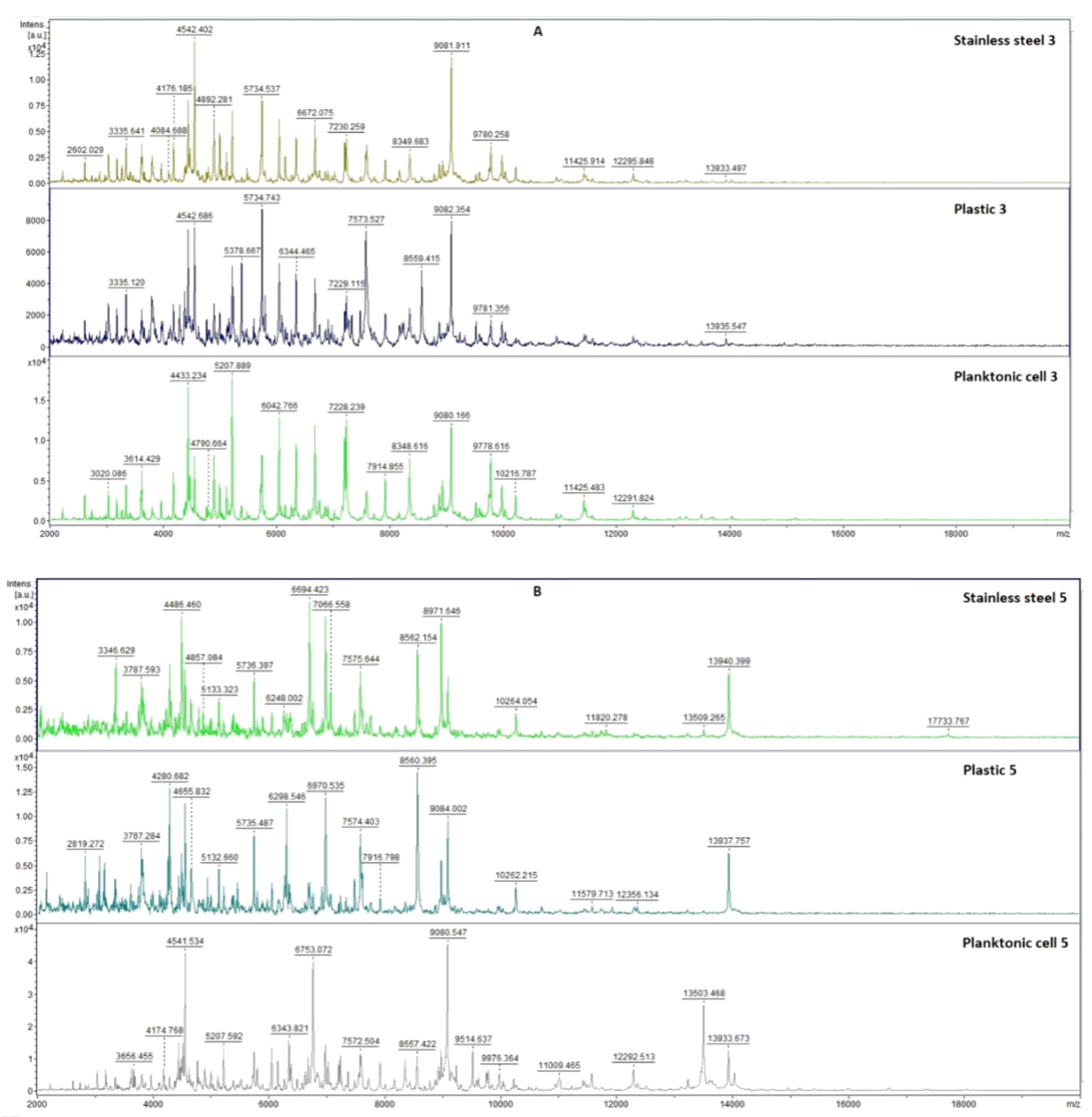
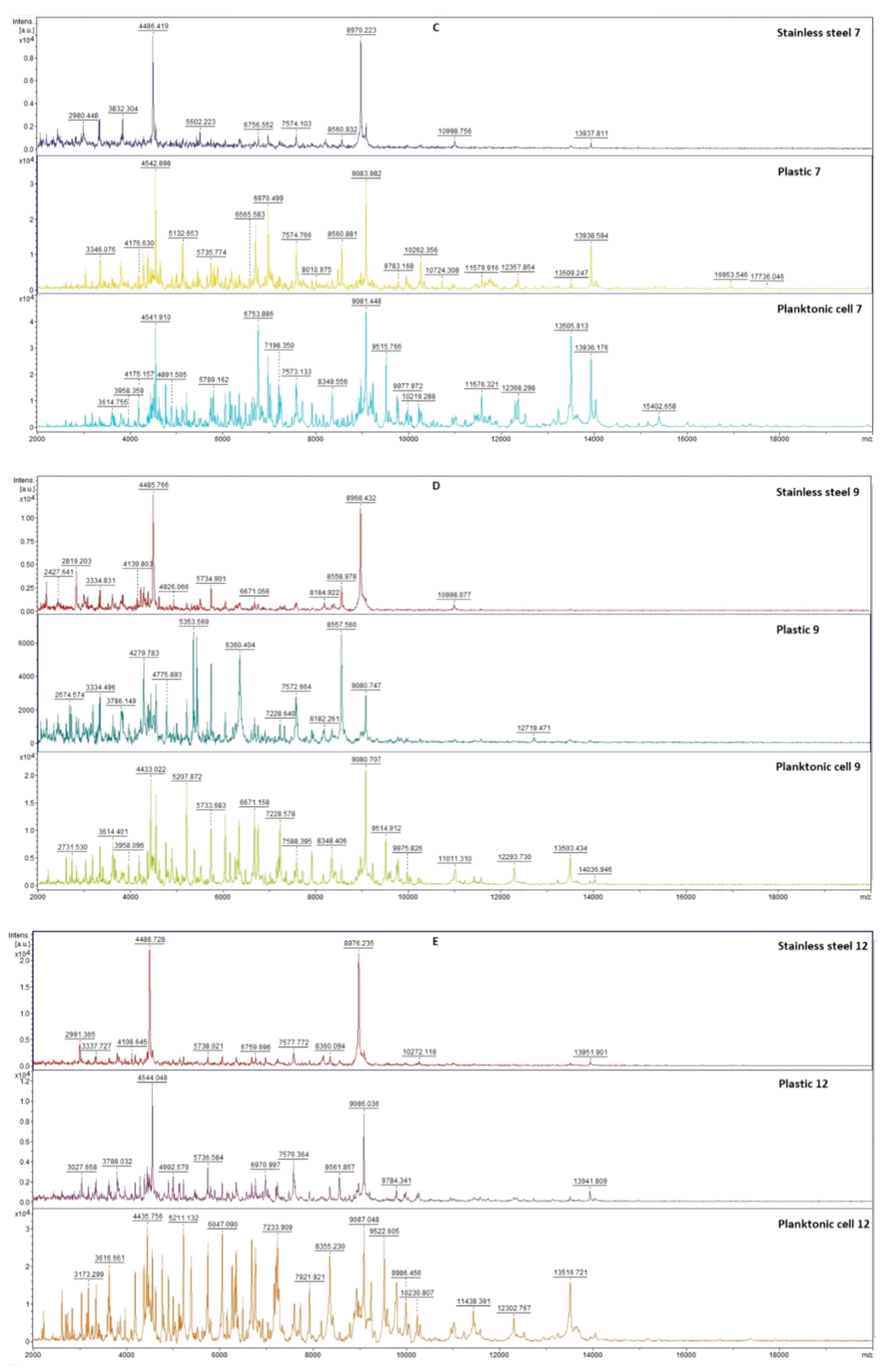
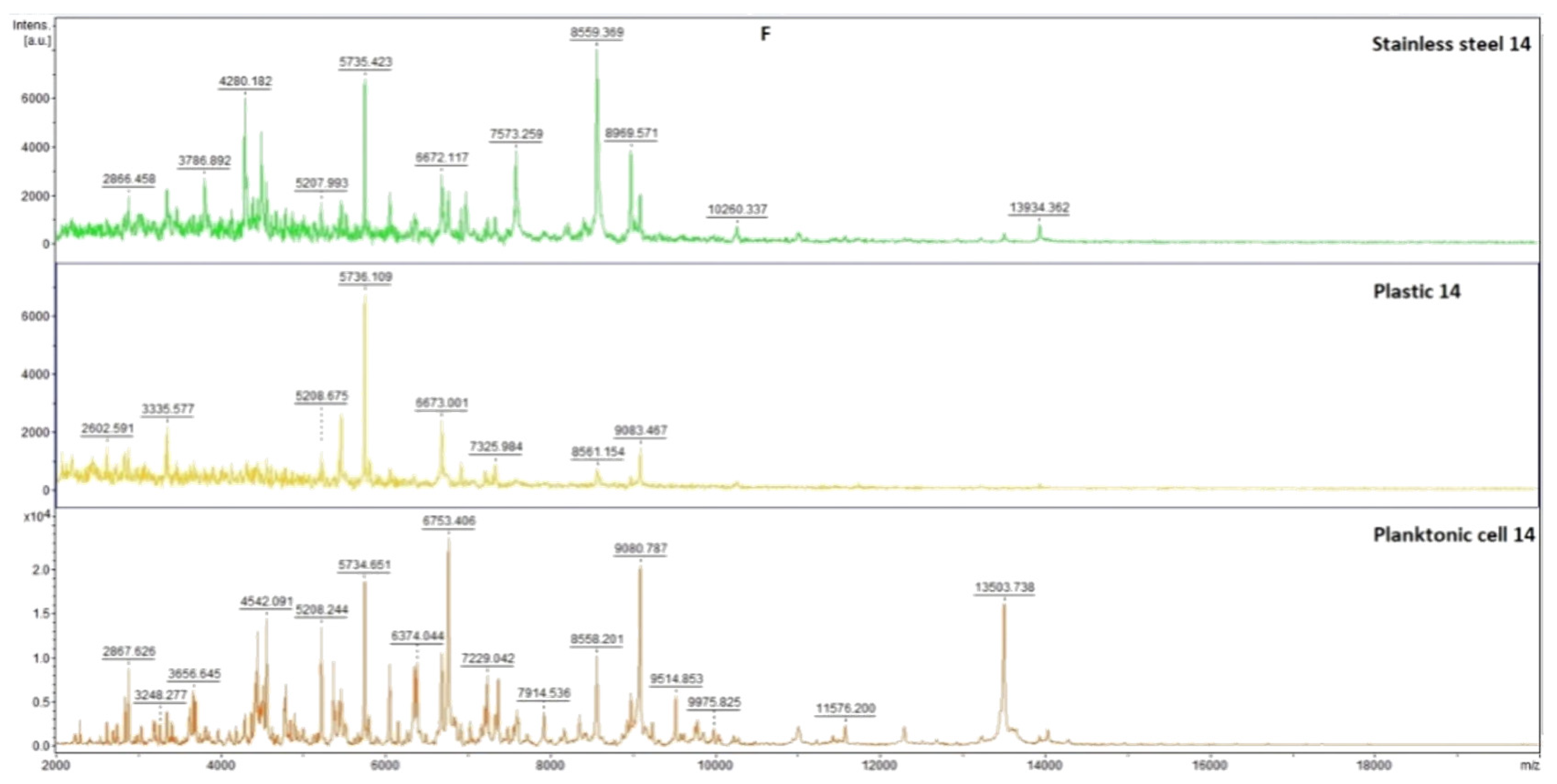
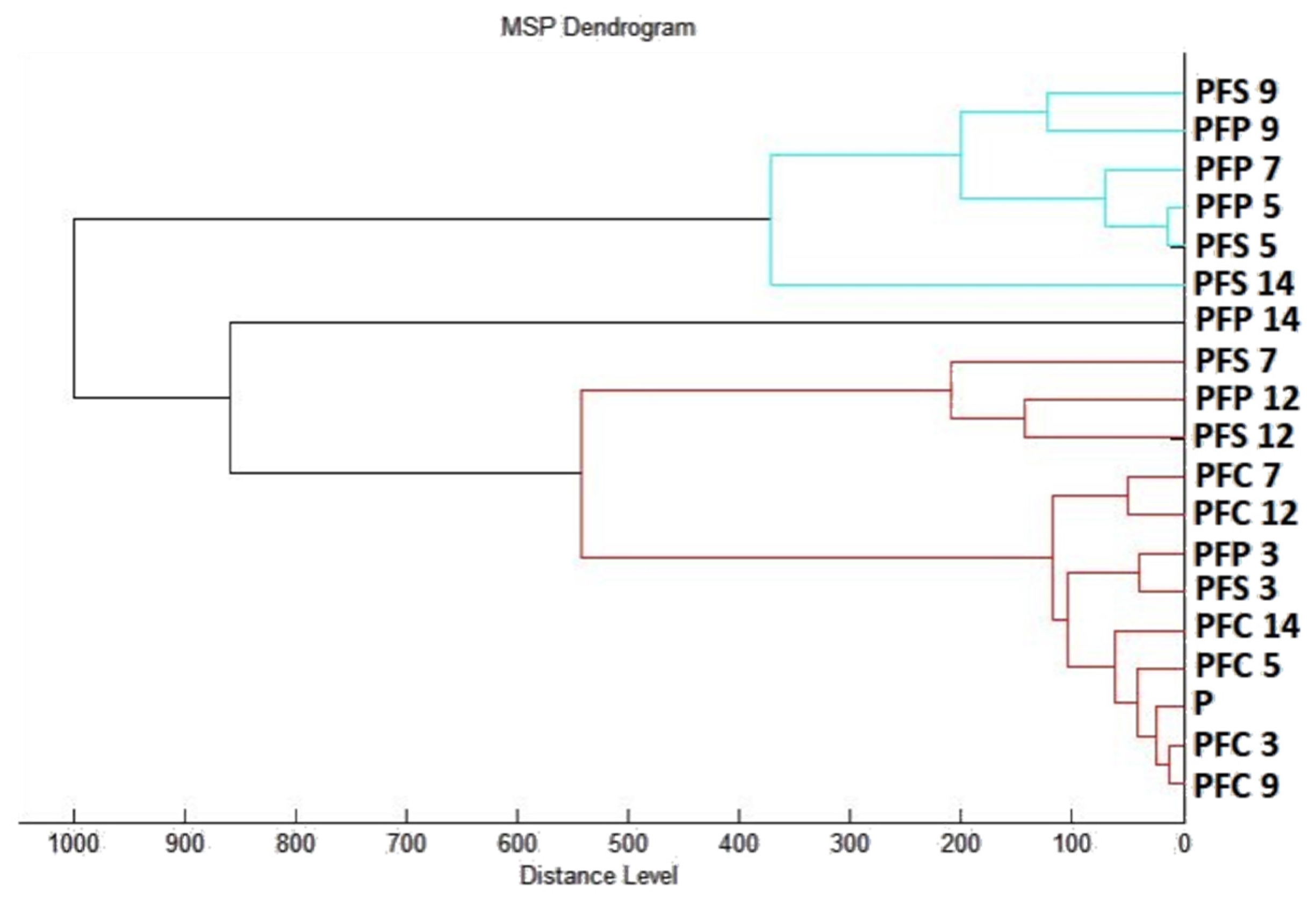
2.7.1. The Effect of T. mastichina EO on P. fluorescens Biofilms
2.7.2. The Effect of E. cardamomum EO on P. fluorescens Biofilms
3. Discussion
4. Materials and Methods
4.1. Essential Oils
4.2. Gas Chromatography–Mass Spectrometry and Gas Chromatography Analyses of T. mastichina and E. cardamomum
4.3. Cell Culture and Treatment
4.4. Determination of Cell Viability (MTT Assay)
4.5. Measurement of Superoxide Anion Radical (NBT Test)
4.6. Measurement of NO Concentration (Griess Method)
4.7. Total Glutathione Concentration
4.8. Transwell Assay for Cell Migration
4.9. Tested Microorganisms
4.10. Antimicrobial Activity—Disc Diffusion Method
4.11. Minimum Inhibitory Concentration (MIC)
4.12. Analysis of Differences in Biofilm Development with MALDI-TOF MS Biotyper
4.13. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bondi, M.; Lauková, A.; de Niederhausern, S.; Messi, P.; Papadopoulou, C. Natural Preservatives to Improve Food Quality and Safety. J. Food Qual. 2017, 2017, 1090932. [Google Scholar] [CrossRef]
- Yap, P.S.X.; Yiap, B.C.; Ping, H.C.; Lim, S.H.E. Essential Oils, A New Horizon in Combating Bacterial Antibiotic Resistance. Open Microbiol. J. 2014, 8, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Key, T.J.; Verkasalo, P.K.; Banks, E. Epidemiology of Breast Cancer. Lancet Oncol. 2001, 2, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Britt, K.L.; Cuzick, J.; Phillips, K.-A. Key Steps for Effective Breast Cancer Prevention. Nat. Rev. Cancer 2020, 20, 417–436. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Benelli, G.; Pavoni, L.; Bonacucina, G.; Cespi, M.; Cianfaglione, K.; Bajalan, I.; Morshedloo, M.R.; Lupidi, G.; Romano, D.; et al. Microemulsions for Delivery of Apiaceae Essential Oils—Towards Highly Effective and Eco-Friendly Mosquito Larvicides? Ind. Crops Prod. 2019, 129, 631–640. [Google Scholar] [CrossRef]
- Baptista-Silva, S.; Borges, S.; Ramos, O.L.; Pintado, M.; Sarmento, B. The Progress of Essential Oils as Potential Therapeutic Agents: A Review. J. Essent. Oil Res. 2020, 32, 279–295. [Google Scholar] [CrossRef]
- Falleh, H.; ben Jemaa, M.; Saada, M.; Ksouri, R. Essential Oils: A Promising Eco-Friendly Food Preservative. Food Chem. 2020, 330, 127268. [Google Scholar] [CrossRef]
- Méndez-Tovar, I.; Martín, H.; Santiago, Y.; Ibeas, A.; Herrero, B.; Asensio-S.-Manzanera, M.C. Variation in Morphological Traits among Thymus mastichina (L.) L. Populations. Genet. Resour. Crop. Evol. 2015, 62, 1257–1267. [Google Scholar] [CrossRef]
- Cutillas, A.-B.; Carrasco, A.; Martinez-Gutierrez, R.; Tomas, V.; Tudela, J. Thymus Mastichina L. Essential Oils from Murcia (Spain): Composition and Antioxidant, Antienzymatic and Antimicrobial Bioactivities. PLoS ONE 2018, 13, e0190790. [Google Scholar] [CrossRef]
- Taghouti, M.; Martins-Gomes, C.; Schäfer, J.; Santos, J.A.; Bunzel, M.; Nunes, F.M.; Silva, A.M. Chemical Characterization and Bioactivity of Extracts from Thymus Mastichina: A Thymus with a Distinct Salvianolic Acid Composition. Antioxidants 2019, 9, 34. [Google Scholar] [CrossRef]
- Gordo, J.; Máximo, P.; Cabrita, E.; Lourenço, A.; Oliva, A.; Almeida, J.; Filipe, M.; Cruz, P.; Barcia, R.; Santos, M.; et al. Thymus mastichina : Chemical Constituents and Their Anti-Cancer Activity. Nat. Prod. Commun 2012, 7, 1934578X1200701. [Google Scholar] [CrossRef]
- Rodrigues, M.; Lopes, A.C.; Vaz, F.; Filipe, M.; Alves, G.; Ribeiro, M.P.; Coutinho, P.; Araujo, A.R.T.S. Thymus Mastichina: Composition and Biological Properties with a Focus on Antimicrobial Activity. Pharmaceuticals 2020, 13, 479. [Google Scholar] [CrossRef] [PubMed]
- Delgado, T.; Marinero, P.; Asensio-S.-Manzanera, M.C.; Asensio, C.; Herrero, B.; Pereira, J.A.; Ramalhosa, E. Antioxidant Activity of Twenty Wild Spanish Thymus Mastichina L. Populations and Its Relation with Their Chemical Composition. LWT Food Sci. Technol. 2014, 57, 412–418. [Google Scholar] [CrossRef]
- Aazza, S.; El-Guendouz, S.; Miguel, M.G.; Antunes, M.D.; Faleiro, M.L.; Correia, A.I.; Figueiredo, A.C. Antioxidant, Anti-Inflammatory and Anti-Hyperglycaemic Activities of Essential Oils from Thymbra capitata, Thymus albicans, Thymus caespititius, Thymus carnosus, Thymus lotocephalus and Thymus mastichina from Portugal. Nat. Prod. Commun. 2016, 11, 1934578X1601100. [Google Scholar] [CrossRef]
- Ashokkumar, K.; Murugan, M.; Dhanya, M.K.; Warkentin, T.D. Botany, Traditional Uses, Phytochemistry and Biological Activities of Cardamom [Elettaria cardamomum (L.) Maton]—A Critical Review. J. Ethnopharmacol. 2020, 246, 112244. [Google Scholar] [CrossRef] [PubMed]
- Anwar, F.; Abbas, A.; Alkharfy, K.M.; Gilani, A.-H. Cardamom (Elettaria cardamomum Maton) Oils. In Essential Oils in Food Preservation, Flavor and Safety; Elsevier: Amsterdam, The Netherlands, 2016; pp. 295–301. [Google Scholar]
- Alam, A.; Rehman, N.U.; Ansari, M.N.; Palla, A.H. Effects of Essential Oils of Elettaria cardamomum Grown in India and Guatemala on Gram-Negative Bacteria and Gastrointestinal Disorders. Molecules 2021, 26, 2546. [Google Scholar] [CrossRef]
- Abdullah; Asghar, A.; Butt, M.S.; Shahid, M.; Huang, Q. Evaluating the Antimicrobial Potential of Green Cardamom Essential Oil Focusing on Quorum Sensing Inhibition of Chromobacterium violaceum. J. Food Sci. Technol. 2017, 54, 2306–2315. [Google Scholar] [CrossRef]
- Elmati, J.; Mountassif, D.; Amarouch, H. Antimicrobial Activity of Elettaria Cardamomum: Toxicity, Biochemical and Histological Studies. Food Chem. 2007, 104, 1560–1568. [Google Scholar] [CrossRef]
- Al-Yousef, H.M.; Alqahtani, A.S.; Hassan, W.H.B.; Alzoubi, A.; Abdelaziz, S. Chemical Profile, In Vitro Antioxidant, Pancreatic Lipase, and Alpha-Amylase Inhibition Assays of the Aqueous Extract of Elettaria cardamomum L. Fruits. J. Chem. 2021, 2021, 5583001. [Google Scholar] [CrossRef]
- Morsy, N.F.S. A Short Extraction Time of High Quality Hydrodistilled Cardamom (Elettaria cardamomum L. Maton) Essential Oil Using Ultrasound as a Pretreatment. Ind. Crops Prod. 2015, 65, 287–292. [Google Scholar] [CrossRef]
- Jamal, A.; Javed, K.; Aslam, M.; Jafri, M.A. Gastroprotective Effect of Cardamom, Elettaria cardamomum Maton. Fruits in Rats. J. Ethnopharmacol. 2006, 103, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Tarfaoui, K.; Brhadda, N.; Ziri, R.; Oubihi, A.; Imtara, H.; Haida, S.; al kamaly, O.M.; Saleh, A.; Parvez, M.K.; Fettach, S.; et al. Chemical Profile, Antibacterial and Antioxidant Potential of Zingiber Officinale Roscoe and Elettaria cardamomum (L.) Maton Essential Oils and Extracts. Plants 2022, 11, 1487. [Google Scholar] [CrossRef] [PubMed]
- Pourkhosravani, E.; Dehghan Nayeri, F.; Mohammadi Bazargani, M. Decoding Antibacterial and Antibiofilm Properties of Cinnamon and Cardamom Essential Oils: A Combined Molecular Docking and Experimental Study. AMB Express 2021, 11, 143. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Majumdar, R.S.; Alam, P. Systematics Evaluations of Morphological Traits, Chemical Composition, and Antimicrobial Properties of Selected Varieties of Elettaria cardamomum (L.) Maton. Nat. Prod. Commun. 2019, 14, 1934578X1989268. [Google Scholar] [CrossRef]
- Savan, E.K.; Küçükbay, F.Z. Essential Oil Composition of Elettaria cardamomum Maton. J. Appl. Biol. Sci. 2013, 7, 42–45. [Google Scholar]
- Masoumi-Ardakani, Y.; Mandegary, A.; Esmaeilpour, K.; Najafipour, H.; Sharififar, F.; Pakravanan, M.; Ghazvini, H. Chemical Composition, Anticonvulsant Activity, and Toxicity of Essential Oil and Methanolic Extract of Elettaria cardamomum. Planta Med. 2016, 82, 1482–1486. [Google Scholar] [CrossRef]
- Souissi, M.; Azelmat, J.; Chaieb, K.; Grenier, D. Antibacterial and Anti-Inflammatory Activities of Cardamom (Elettaria cardamomum) Extracts: Potential Therapeutic Benefits for Periodontal Infections. Anaerobe 2020, 61, 102089. [Google Scholar] [CrossRef]
- Blowman, K.; Magalhães, M.; Lemos, M.F.L.; Cabral, C.; Pires, I.M. Anticancer Properties of Essential Oils and Other Natural Products. Evid. -Based Complement. Altern. Med. 2018, 2018, 3149362. [Google Scholar] [CrossRef]
- Huang, J.; Chan, P.S.; Lok, V.; Chen, X.; Ding, H.; Jin, Y.; Yuan, J.; Lao, X.; Zheng, Z.-J.; Wong, M.C. Global Incidence and Mortality of Breast Cancer: A Trend Analysis. Aging 2021, 13, 5748–5803. [Google Scholar] [CrossRef]
- de Fátima Braga Magalhães, I.; Junqueira Moraga Tellis, C.; da Silva Calabrese, K.; Lucia Abreu-Silva, A.; Almeida-Souza, F. Essential Oils’ Potential in Breast Cancer Treatment: An Overview. In Essential Oils—Bioactive Compounds, New Perspectives and Applications; IntechOpen: London, UK, 2020. [Google Scholar]
- Bagheri, S.M.; Asl, A.A.; Shams, A.; Mirghanizadeh-Bafghi, S.A.; Hafizibarjin, Z. Evaluation of Cytotoxicity Effects of Oleo-Gum-Resin and Its Essential Oil of Ferula Assa-Foetida and Ferulic Acid on 4T1 Breast Cancer Cells. Indian J. Med. Paediatr. Oncol. 2017, 38, 116–120. [Google Scholar] [CrossRef]
- Yeo, S.K.; Ali, A.Y.; Hayward, O.A.; Turnham, D.; Jackson, T.; Bowen, I.D.; Clarkson, R. β-Bisabolene, a Sesquiterpene from the Essential Oil Extract of Opoponax (Commiphora guidottii), Exhibits Cytotoxicity in Breast Cancer Cell Lines. Phytother. Res. 2016, 30, 418–425. [Google Scholar] [CrossRef]
- Torgovnick, A.; Schumacher, B. DNA Repair Mechanisms in Cancer Development and Therapy. Front. Genet. 2015, 6, 157. [Google Scholar] [CrossRef]
- Fitsiou, E.; Pappa, A. Anticancer Activity of Essential Oils and Other Extracts from Aromatic Plants Grown in Greece. Antioxidants 2019, 8, 290. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, Y.; Sarkar, F. Signaling Mechanism(S) of Reactive Oxygen Species in Epithelial-Mesenchymal Transition Reminiscent of Cancer Stem Cells in Tumor Progression. Curr. Stem Cell Res. Ther. 2010, 5, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Xiang, J.; Su, L.; Tang, X. The Regulation of Nitric Oxide in Tumor Progression and Therapy. J. Int. Med. Res. 2020, 48, 030006052090598. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Wu, Z.; Li, Z.; Liu, L. Celastrol Inhibits Migration and Invasion of Triple-Negative Breast Cancer Cells by Suppressing Interleukin-6 via Downregulating Nuclear Factor-ΚB (NF-ΚB). Med. Sci. Monit. 2020, 26, e922814-1–e922814-9. [Google Scholar] [CrossRef]
- Xing, X.; Ma, J.-H.; Fu, Y.; Zhao, H.; Ye, X.-X.; Han, Z.; Jia, F.-J.; Li, X. Essential Oil Extracted from Erythrina corallodendron L. Leaves Inhibits the Proliferation, Migration, and Invasion of Breast Cancer Cells. Medicine 2019, 98, e17009. [Google Scholar] [CrossRef]
- Faleiro, M.L.; Miguel, M.G.; Ladeiro, F.; Venancio, F.; Tavares, R.; Brito, J.C.; Figueiredo, A.C.; Barroso, J.G.; Pedro, L.G. Antimicrobial Activity of Essential Oils Isolated from Portuguese Endemic Species of Thymus. Lett. Appl. Microbiol. 2003, 36, 35–40. [Google Scholar] [CrossRef]
- Ballester-Costa, C.; Sendra, E.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Chemical Composition and in Vitro Antibacterial Properties of Essential Oils of Four Thymus Species from Organic Growth. Ind. Crops Prod. 2013, 50, 304–311. [Google Scholar] [CrossRef]
- Al-Zereini, W.A.; Al-Trawneh, I.N.; Al-Qudah, M.A.; TumAllah, H.M.; al Rawashdeh, H.A.; Abudayeh, Z.H. Essential Oils from Elettaria cardamomum (L.) Maton Grains and Cinnamomum Verum J. Presl Barks: Chemical Examination and Bioactivity Studies. J. Pharm. Pharmacogn. Res. 2022, 10, 173–185. [Google Scholar] [CrossRef]
- Dias-Souza, M.V.; dos Santos, R.M.; de Siqueira, E.P.; Ferreira-Marçal, P.H. Antibiofilm Activity of Cashew Juice Pulp against Staphylococcus Aureus, High Performance Liquid Chromatography/Diode Array Detection and Gas Chromatography-Mass Spectrometry Analyses, and Interference on Antimicrobial Drugs. J. Food Drug Anal. 2017, 25, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Budzyńska, A.; Różalska, S.; Sadowska, B.; Różalska, B. Candida albicans/Staphylococcus aureus Dual-Species Biofilm as a Target for the Combination of Essential Oils and Fluconazole or Mupirocin. Mycopathologia 2017, 182, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Jafri, H.; Ahmad, I. Thymus Vulgaris Essential Oil and Thymol Inhibit Biofilms and Interact Synergistically with Antifungal Drugs against Drug Resistant Strains of Candida albicans and Candida tropicalis. J. Mycol. Med. 2020, 30, 100911. [Google Scholar] [CrossRef]
- Xie, Y.-Y.; Zhang, Y.-W.; Liu, X.-Z.; Ma, X.-F.; Qin, X.-T.; Jia, S.-R.; Zhong, C. Aggregation-Induced Emission-Active Amino Acid/Berberine Hydrogels with Enhanced Photodynamic Antibacterial and Anti-Biofilm Activity. Chem. Eng. J. 2021, 413, 127542. [Google Scholar] [CrossRef]
- Kačániová, M.; Galovičová, L.; Borotová, P.; Valková, V.; Ďúranová, H.; Kowalczewski, P.Ł.; Said-Al Ahl, H.A.H.; Hikal, W.M.; Vukic, M.; Savitskaya, T.; et al. Chemical Composition, In Vitro and In Situ Antimicrobial and Antibiofilm Activities of Syzygium aromaticum (Clove) Essential Oil. Plants 2021, 10, 2185. [Google Scholar] [CrossRef]
- Vieira, M.; Bessa, L.J.; Martins, M.R.; Arantes, S.; Teixeira, A.P.S.; Mendes, Â.; Martins da Costa, P.; Belo, A.D.F. Chemical Composition, Antibacterial, Antibiofilm and Synergistic Properties of Essential Oils from Eucalyptus globulus Labill. and Seven Mediterranean Aromatic Plants. Chem. Biodivers. 2017, 14, e1700006. [Google Scholar] [CrossRef] [PubMed]
- Abdullah; Asghar, A.; Algburi, A.; Huang, Q.; Ahmad, T.; Zhong, H.; Javed, H.U.; Ermakov, A.M.; Chikindas, M.L. Anti-Biofilm Potential of Elletaria cardamomum Essential Oil against Escherichia coli O157:H7 and Salmonella Typhimurium JSG 1748. Front. Microbiol. 2021, 12, 620227. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy; Allured Pub. Corp.: Carol Stream, IL, USA, 2007; ISBN 9781932633214. [Google Scholar]
- van den Dool, H.; Dec. Kratz, P. A Generalization of the Retention Index System Including Linear Temperature Programmed Gas—Liquid Partition Chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Auclair, C.; Voisin, E. Nitroblue Tetrazolium Reduction. In Handbook Methods for Oxygen Radical Research; CRC Press: Boca Raton, FL, USA, 1985. [Google Scholar]
- Griess, P. Bemerkungen Zu Der Abhandlung Der HH. Weselsky Und Benedikt ”Ueber Einige Azoverbindungen”. Ber. Der Dtsch. Chem. Ges. 1879, 12, 426–428. [Google Scholar] [CrossRef]
- Beutler, E. Reduced Glutathione (GSH). In Red Blood Cell Metabolism: A Manual of Biochemical Methods, 2nd ed.; Bergmeyen, H.V., Ed.; Grune and Stratton: New York, NY, USA, 1975; pp. 112–114. [Google Scholar]
- Chen, H.-C. Boyden Chamber Assay. In Cell Migration; Humana Press: Totowa, NJ, USA, 2005; pp. 15–22. [Google Scholar]
- Kačániová, M.; Galovičová, L.; Valková, V.; Tvrdá, E.; Terentjeva, M.; Žiarovská, J.; Kunová, S.; Savitskaya, T.; Grinshpan, D.; Štefániková, J.; et al. Antimicrobial and Antioxidant Activities of Cinnamomum cassia Essential Oil and Its Application in Food Preservation. Open Chem. 2021, 19, 214–227. [Google Scholar] [CrossRef]
- Kačániová, M.; Terentjeva, M.; Galovičová, L.; Ivanišová, E.; Štefániková, J.; Valková, V.; Borotová, P.; Kowalczewski, P.Ł.; Kunová, S.; Felšöciová, S.; et al. Biological Activity and Antibiofilm Molecular Profile of Citrus aurantium Essential Oil and Its Application in a Food Model. Molecules 2020, 25, 3956. [Google Scholar] [CrossRef] [PubMed]
- Galovičová, L.; Borotová, P.; Valková, V.; Vukovic, N.L.; Vukic, M.; Štefániková, J.; Ďúranová, H.; Kowalczewski, P.Ł.; Čmiková, N.; Kačániová, M. Thymus vulgaris Essential Oil and Its Biological Activity. Plants 2021, 10, 1959. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Compound b | % | AI a | KI a | ||
|---|---|---|---|---|---|---|
| (lit.) | (calc.) | (lit.) | (calc.) | |||
| 1 | tricyclene | tr c | 912 | 922 | 926 | 925 |
| 2 | α-thujene | 0.1 | 924 | 924 | 930 | 928 |
| 3 | α-pinene | 3.7 | 932 | 932 | 939 | 937 |
| 4 | camphene | 0.6 | 946 | 949 | 954 | 953 |
| 5 | verbenene | tr | 961 | 952 | 967 | 957 |
| 6 | sabinene | 2.1 | 969 | 971 | 975 | 975 |
| 7 | β-pinene | 4 | 974 | 976 | 979 | 980 |
| 8 | 3-octanone | tr | 979 | 983 | 983 | 986 |
| 9 | β-myrcene | 1.5 | 988 | 987 | 990 | 990 |
| 10 | dehydro-1,8-cineole | tr | 988 | 990 | 991 | 991 |
| 11 | 3-octanol | tr | 988 | 998 | 991 | 997 |
| 12 | δ-2-carene | tr | 1001 | 1000 | 1002 | 999 |
| 13 | α-phellandrene | 0.1 | 1002 | 1007 | 1002 | 1007 |
| 14 | δ-3-carene | tr | 1008 | 1009 | 1011 | 1009 |
| 15 | α-terpinene | 0.2 | 1014 | 1016 | 1017 | 1019 |
| 16 | p-cymene | 0.8 | 1020 | 1024 | 1024 | 1027 |
| 17 | limonene | 2.1 | 1024 | 1028 | 1029 | 1031 |
| 18 | 1,8-cineole | 47.1 | 1026 | 1037 | 1031 | 1038 |
| 19 | (E)-β-ocimene | 0.7 | 1044 | 1044 | 1050 | 1049 |
| 20 | γ-terpinene | 0.4 | 1054 | 1056 | 1059 | 1060 |
| 21 | cis-linalool oxide | 0.3 | 1067 | 1069 | 1072 | 1072 |
| 22 | α-terpinolene | 0.1 | 1086 | 1084 | 1088 | 1085 |
| 23 | trans-linalool oxide | 0.2 | 1084 | 1086 | 1086 | 1087 |
| 24 | p-cymenene | tr | 1089 | 1089 | 1091 | 1090 |
| 25 | linalool | 19.4 | 1095 | 1104 | 1096 | 1104 |
| 26 | hotrienol | 0.2 | e | 1105 | 1109 d | 1106 |
| 27 | cis-p-menth-2-en-1-ol | tr | 1118 | 1124 | 1137 | 1141 |
| 28 | nopinone | tr | 1135 | 1138 | 1140 | 1142 |
| 29 | trans-pinocarveol | 0.1 | 1135 | 1140 | 1139 | 1145 |
| 30 | camphor | 0.7 | 1141 | 1147 | 1146 | 1151 |
| 31 | pinocarvone | tr | 1160 | 1162 | 1164 | 1166 |
| 32 | δ-terpineol | 1.2 | 1162 | 1170 | 1169 | 1174 |
| 33 | borneol | 0.7 | 1165 | 1172 | 1169 | 1175 |
| 34 | 4-terpineol | 1.2 | 1174 | 1180 | 1177 | 1182 |
| 35 | p-cymen-8-ol | tr | 1179 | 1187 | 1182 | 1188 |
| 36 | α-terpineol | 4.6 | 1186 | 1196 | 1188 | 1196 |
| 37 | verbenone | tr | 1204 | 1207 | 1205 | 1205 |
| 38 | trans-carveol | tr | 1215 | 1218 | 1216 | 1219 |
| 39 | nerol | tr | 1227 | 1223 | 1229 | 1225 |
| 40 | citronellol | tr | 1223 | 1225 | 1225 | 1229 |
| 41 | isobornyl formate | 0.1 | 1235 | 1227 | 1239 | 1229 |
| 42 | carvone | tr | 1239 | 1242 | 1243 | 1245 |
| 43 | linalool acetate | 1.8 | 1254 | 1249 | 1257 | 1253 |
| 44 | geranial | tr | 1264 | 1266 | 1267 | 1270 |
| 45 | bornyl acetate | 0.3 | 1284 | 1283 | 1285 | 1284 |
| 46 | thymol | 0.1 | 1289 | 1289 | 1290 | 1290 |
| 47 | δ-terpinyl acetate | 0.3 | 1316 | 1311 | 1317 | 1313 |
| 48 | α-terpinyl acetate | 1.8 | 1346 | 1345 | 1349 | 1348 |
| 49 | neryl acetate | 0.1 | 1359 | 1357 | 1361 | 1360 |
| 50 | α-ylangene (α-copaene) | tr | 1373 | 1373 | 1375 | 1376 |
| 51 | geranyl acetate | 0.2 | 1379 | 1377 | 1381 | 1379 |
| 52 | β-bourbonene | tr | 1387 | 1381 | 1388 | 1383 |
| 53 | β-elemene | tr | 1389 | 1387 | 1390 | 1389 |
| 54 | α-gurjunene | tr | 1409 | 1404 | 1409 | 1405 |
| 55 | trans-caryophyllene | 1.1 | 1417 | 1417 | 1419 | 1419 |
| 56 | α-trans-bergamotene | tr | 1432 | 1430 | 1434 | 1433 |
| 57 | aromadendrene | 0.1 | 1439 | 1435 | 1441 | 1439 |
| 58 | α-humulene | tr | 1452 | 1452 | 1454 | 1456 |
| 59 | germacrene D | 0.1 | 1480 | 1478 | 1481 | 1481 |
| 60 | viridiflorene | 0.1 | 1496 | 1489 | 1496 | 1492 |
| 61 | bicyclogermacrene | 0.2 | 1500 | 1492 | 1500 | 1494 |
| 62 | α-muurolene | tr | 1500 | 1495 | 1500 | 1497 |
| 63 | δ-amorphene | 0.1 | 1511 | 1509 | 1512 | 1512 |
| 64 | γ-cadinene | 0.1 | 1513 | 1515 | 1513 | 1518 |
| 65 | α-cadinene | tr | 1537 | 1533 | 1538 | 1537 |
| 66 | elemol | 0.2 | 1548 | 1545 | 1549 | 1549 |
| 67 | spathulenol | 0.1 | 1577 | 1573 | 1578 | 1577 |
| 68 | caryophyllene oxide | 0.1 | 1581 | 1578 | 1582 | 1583 |
| 69 | veridiflorol | 0.2 | 1592 | 1590 | 1592 | 1594 |
| 70 | γ-eudesmol | 0.1 | 1630 | 1628 | 1632 | 1628 |
| 71 | α-eudesmol | 0.2 | 1651 | 1651 | 1653 | 1648 |
| total | 99.5 | |||||
| Class of Compounds | % |
|---|---|
| monoterpenes | 96.6 |
| monoterpene hydrocarbons | 16.4 |
| oxygenated monoterpenes | 80.2 |
| monoterpene epoxide | 47.1 |
| monoterpene alcohols | 27.8 |
| monoterpene ketones | 0.7 |
| monoterpene esters | 4.6 |
| sesquiterpenes | 2.7 |
| sesquiterpene hydrocarbons | 1.8 |
| oxygenated sesquiterpenes | 0.9 |
| sesquiterpene alcohols | 0.8 |
| sesquiterpene epoxides | 0.1 |
| nonterpenic compounds | 0.2 |
| ketones | tr |
| alcohols | 0.2 |
| total | 99.5 |
| No | Compound b | % | AI a | KI a | ||
|---|---|---|---|---|---|---|
| (lit.) | (calc.) | (lit.) | (calc.) | |||
| 1 | α-thujene | 0.3 | 924 | 923 | 930 | 926 |
| 2 | α-pinene | 1.9 | 932 | 931 | 939 | 933 |
| 3 | α-fenchene | tr c | 945 | 946 | 952 | 947 |
| 4 | camphene | tr | 946 | 948 | 954 | 949 |
| 5 | sabinene | 5.1 | 969 | 971 | 975 | 973 |
| 6 | β-pinene | 0.5 | 974 | 977 | 979 | 977 |
| 7 | 6-methyl-5-hepten-2-one | tr | 981 | 981 | 985 | 985 |
| 8 | β-myrcene | 2.1 | 988 | 987 | 990 | 990 |
| 9 | octanal | tr | 998 | 1003 | 998 | 1004 |
| 10 | α-phellandrene | tr | 1002 | 1006 | 1002 | 1007 |
| 11 | α-terpinene | 0.1 | 1014 | 1016 | 1017 | 1020 |
| 12 | p-cymene | 0.3 | 1020 | 1023 | 1024 | 1029 |
| 13 | Limonene | 2.3 | 1024 | 1028 | 1029 | 1034 |
| 14 | 1,8-cineole | 28.6 | 1026 | 1034 | 1031 | 1041 |
| 15 | (E)-β-ocimene | tr | 1044 | 1043 | 1050 | 1052 |
| 16 | γ-terpinene | 0.8 | 1054 | 1056 | 1059 | 1063 |
| 17 | cis-sabinene hydrate | 0.7 | 1065 | 1069 | 1070 | 1074 |
| 18 | α-terpinolene | 0.1 | 1086 | 1084 | 1088 | 1087 |
| 19 | linalool | 4.5 | 1095 | 1100 | 1096 | 1101 |
| 20 | (E)-4,8-dimethyl-1,3,7-nonatriene | tr | e | 1112 | 1110 d | 1114 |
| 21 | cis-p-menth-2-en-1-ol | tr | 1118 | 1123 | 1121 | 1127 |
| 22 | cis-limonene oxide | tr | 1132 | 1131 | 1136 | 1135 |
| 23 | trans-limonene oxide | tr | 1137 | 1135 | 1142 | 1140 |
| 24 | δ-terpineol | tr | 1162 | 1169 | 1166 | 1173 |
| 25 | borneol | tr | 1165 | 1171 | 1169 | 1174 |
| 26 | 4-terpineol | 1.1 | 1174 | 1180 | 1177 | 1182 |
| 27 | p-cymen-8-ol | tr | 1179 | 1186 | 1182 | 1188 |
| 28 | α-terpineol | 2.4 | 1186 | 1195 | 1188 | 1196 |
| 29 | decanal | tr | 1201 | 1206 | 1201 | 1207 |
| 30 | octanol acetate | 0.1 | 1211 | 1209 | 1213 | 1211 |
| 31 | neral | 0.2 | 1235 | 1237 | 1238 | 1240 |
| 32 | linalool acetate | 7.9 | 1254 | 1250 | 1257 | 1254 |
| 33 | geranial | 0.3 | 1267 | 1266 | 1264 | 1270 |
| 34 | bornyl acetate | tr | 1284 | 1283 | 1285 | 1285 |
| 35 | δ-terpinyl acetate | tr | 1316 | 1313 | 1317 | 1313 |
| 36 | methyl geranate | 0.1 | 1322 | 1320 | 1324 | 1322 |
| 37 | α-terpinyl acetate | 37.3 | 1346 | 1352 | 1349 | 1355 |
| 38 | α-ylangene | tr | 1373 | 1368 | 1375 | 1372 |
| 39 | geranyl acetate | 0.9 | 1379 | 1377 | 1381 | 1379 |
| 40 | β-elemene | tr | 1389 | 1387 | 1390 | 1390 |
| 41 | aromadendrene | tr | 1439 | 1446 | 1441 | 1451 |
| 42 | germacrene D | tr | 1480 | 1478 | 1481 | 1482 |
| 43 | β-selinene | 0.3 | 1489 | 1486 | 1490 | 1490 |
| 44 | α-selinene | 0.1 | 1498 | 1492 | 1498 | 1496 |
| 45 | γ-cadinene | 0.2 | 1513 | 1510 | 1513 | 1514 |
| 46 | (E)-nerolidol | 1.3 | 1561 | 1559 | 1563 | 1562 |
| Total | 99.5 | |||||
| Class of Compounds | % |
|---|---|
| monoterpenes | 97.5 |
| monoterpene hydrocarbons | 13.5 |
| oxygenated monoterpenes | 84.0 |
| monoterpene epoxide | 28.6 |
| monoterpene alcohols | 9.0 |
| monoterpene aldehydes | 0.2 |
| monoterpene esters | 46.2 |
| sesquiterpenes | 1.9 |
| sesquiterpene hydrocarbons | 0.6 |
| oxygenated sesquiterpenes | 1.3 |
| sesquiterpene alcohols | 1.3 |
| nonterpenic compounds | 0.1 |
| ketones | tr |
| aldehydes | tr |
| esters | 0.1 |
| total | 99.5 |
| Microorganism | Zone of Inhibition (mm) | Activity of EO | Zone of Inhibition (mm) | Activity of EO |
|---|---|---|---|---|
| T. mastichina EO | E. cardamomum EO | |||
| Escherichia coli CCM 3954 | 5.33 ± 0.58 a,b | * | 1.33 ± 0.58 b,a | - |
| Haemophilus influenzae CCM 4454 | 8.00 ± 1.00 a,b | * | 10.33 ± 0.58 b,a | ** |
| Streptococcus pneumoniae CCM 4501 | 6.33 ± 0.58 a,b | * | 1.00 ± 0.00 b,a | - |
| Listeria monocytogenes CCM 4699 | 9.00 ± 1.00 a,b | * | 1.00 ± 0.00 b,a | - |
| Staphylococcus aureus CCM 2461 | 6.33 ± 1.53 a,b | * | 1.67 ± 0.58 b,a | - |
| Yersinia enterocolitica CCM 7204 | 5.67 ± 0.58 a,b | * | 10.00 ± 0.00 b,a | * |
| Pseudomonas fluorescens-biofilm | 5.67 ± 0.58 a,b | * | 13.33 ± 1.15 b,a | ** |
| Candida glabrata CCM 8270 | 9.33 ± 1.15 a,b | * | 11.33 ± 1.53 b,a | ** |
| Candida tropicalis CCM 8264 | 8.33 ± 1.53 a,b | * | 11.00 ± 1.00 b,a | ** |
| Candida albicans CCM 8261 | 6.33 ± 0.58 a,b | * | 8.00 ± 1.00 b,a | * |
| Microorganism | MIC 50 (µL/mL) | MIC 90 (µL/mL) | MIC 50 (µL/mL) | MIC 90 (µL/mL) |
|---|---|---|---|---|
| T. mastichina EO | E. cardamomum EO | |||
| Escherichia coli CCM 3954 | 3.56 | 5.75 | 49.86 | 56.21 |
| Haemophilus influenzae CCM 4454 | 0.86 | 2.11 | 1.65 | 3.48 |
| Streptococcus pneumoniae CCM 4501 | 12.35 | 15.46 | 49.86 | 56.21 |
| Listeria monocytogenes CCM 4699 | 1.65 | 3.48 | 49.86 | 56.21 |
| Staphylococcus aureus CCM 2461 | 12.35 | 15.46 | 49.86 | 56.21 |
| Yersinia enterocolitica CCM 7204 | 0.86 | 2.11 | 1.65 | 3.48 |
| Pseudomonas fluorescens-biofilm | 6.36 | 9.54 | 0.86 | 2.11 |
| Candida glabrata CCM 8270 | 0.86 | 2.11 | 0.86 | 2.11 |
| Candida tropicalis CCM 8264 | 6.36 | 9.54 | 1.65 | 3.48 |
| Candida albicans CCM 8261 | 12.35 | 15.46 | 12.35 | 15.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vukovic, N.L.; Vukic, M.D.; Obradovic, A.D.; Matic, M.M.; Galovičová, L.; Kačániová, M. GC, GC/MS Analysis, and Biological Effects of Essential Oils from Thymus mastchina and Elettaria cardamomum. Plants 2022, 11, 3213. https://doi.org/10.3390/plants11233213
Vukovic NL, Vukic MD, Obradovic AD, Matic MM, Galovičová L, Kačániová M. GC, GC/MS Analysis, and Biological Effects of Essential Oils from Thymus mastchina and Elettaria cardamomum. Plants. 2022; 11(23):3213. https://doi.org/10.3390/plants11233213
Chicago/Turabian StyleVukovic, Nenad L., Milena D. Vukic, Ana D. Obradovic, Milos M. Matic, Lucia Galovičová, and Miroslava Kačániová. 2022. "GC, GC/MS Analysis, and Biological Effects of Essential Oils from Thymus mastchina and Elettaria cardamomum" Plants 11, no. 23: 3213. https://doi.org/10.3390/plants11233213
APA StyleVukovic, N. L., Vukic, M. D., Obradovic, A. D., Matic, M. M., Galovičová, L., & Kačániová, M. (2022). GC, GC/MS Analysis, and Biological Effects of Essential Oils from Thymus mastchina and Elettaria cardamomum. Plants, 11(23), 3213. https://doi.org/10.3390/plants11233213