
Nitric Oxide and Salicylic Acid Regulate Glutathione and Ethylene Production to Enhance Heat Stress Acclimation in Wheat Involving Sulfur Assimilation


 ,
,  ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
2.1. Screening of T. aestivum Cultivars for HS Tolerance
2.2. Effect of NO, S and SA on Growth Parameters and Photosynthetic Characteristics under HS
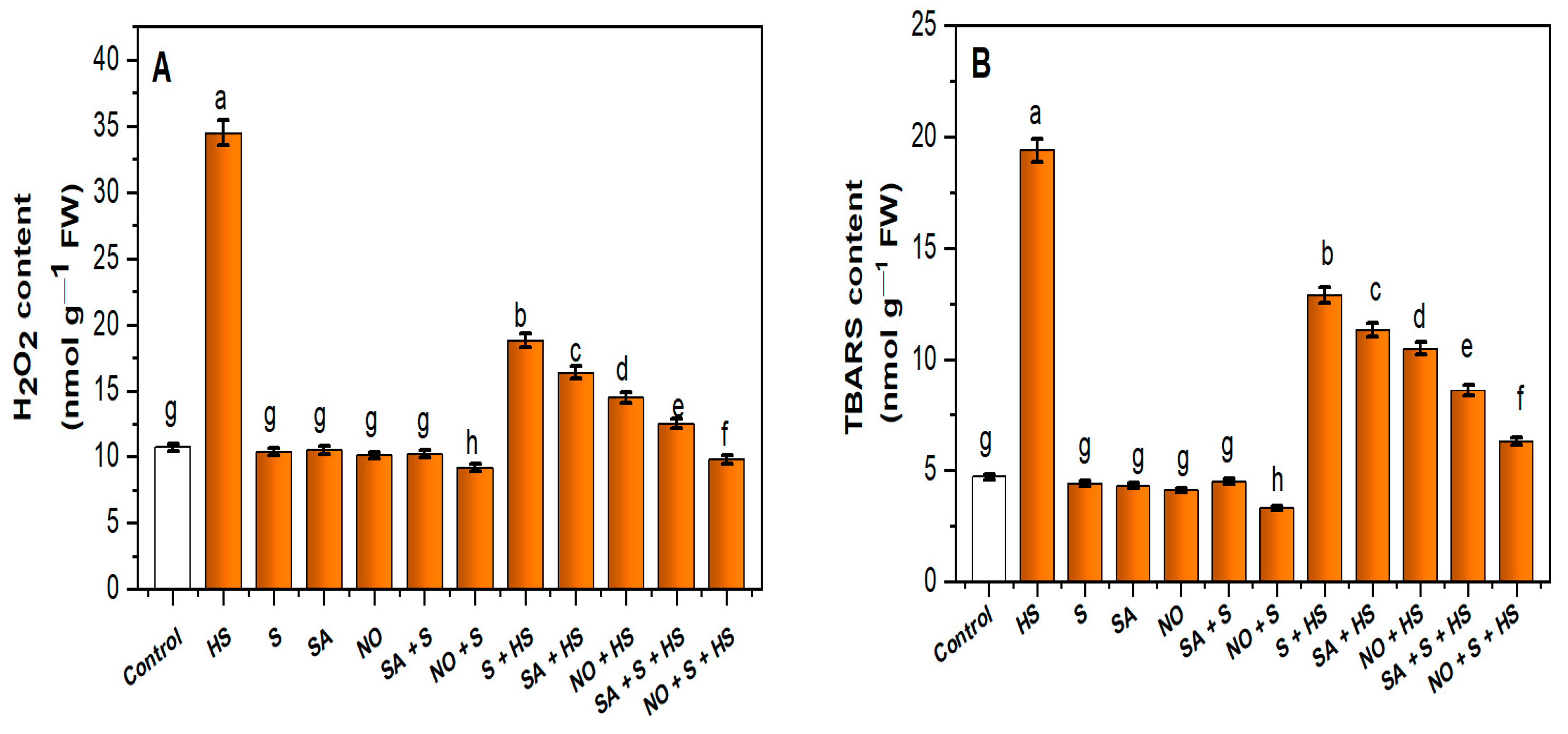
2.3. Application of NO, S and SA Reduced H2O2 and Thiobarbituric Acid Reactive Substances (TBARS) Contents under HS
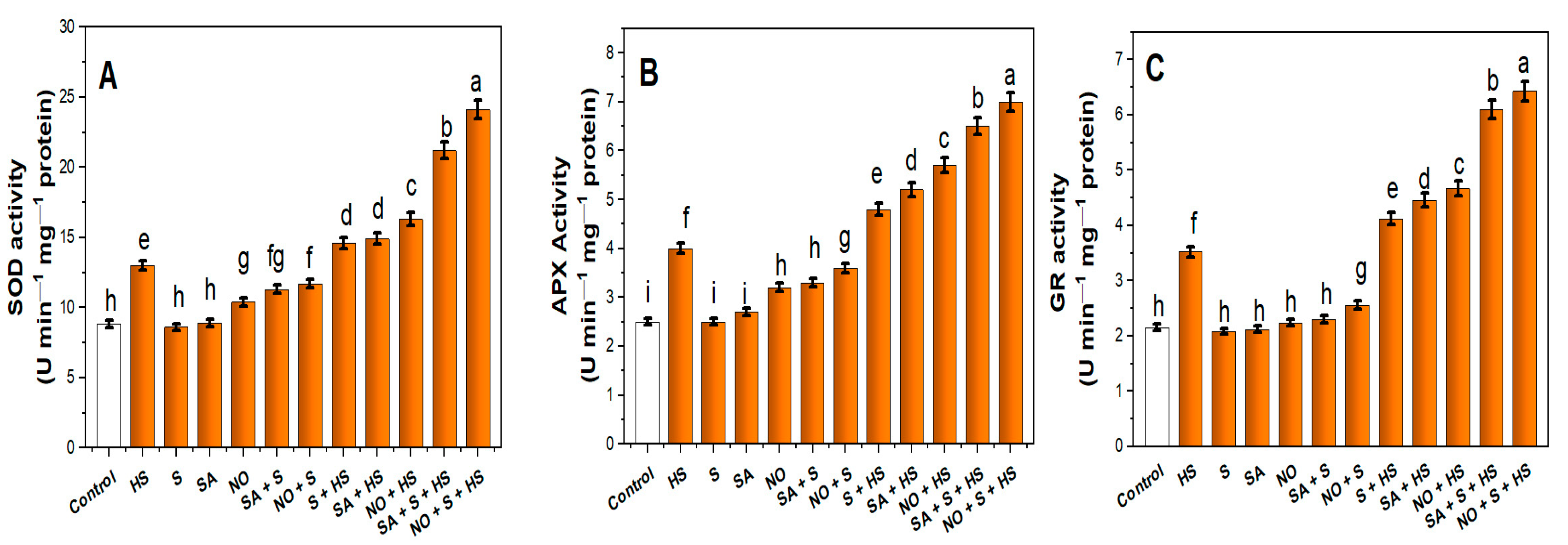
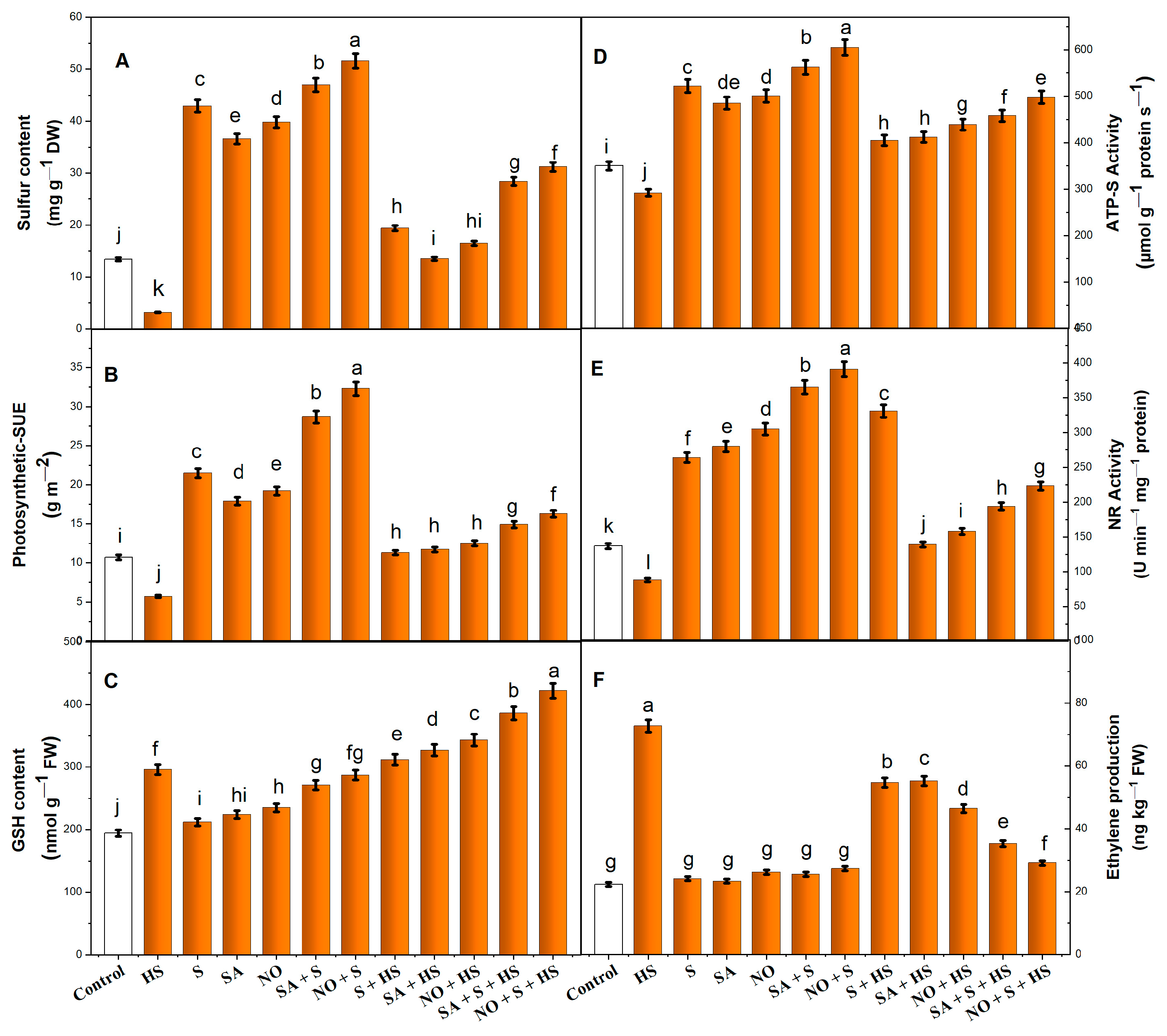
2.4. Application of NO, S and SA Stimulated Antioxidant Enzyme Activity and S Assimilation under HS
2.5. Application of NO, S and SA Maintains Nitrate Reductase (NR) Activity under HS
2.6. Impact of NO, S and SA on Ethylene Production under HS
2.7. Influence of SA Biosynthesis Inhibitor (AIP) and NO Biosynthesis Inhibitor (cPTIO) on Growth and Photosynthetic Parameters under HS
2.8. Influence of SA Biosynthesis Inhibitor (AIP) and NO Biosynthesis Inhibitor (cPTIO) on Oxidative Stress and Antioxidant Enzymes under HS
2.9. Influence of SA Biosynthesis Inhibitor (AIP) and NO Scavenger (cPTIO) on the Assimilation of S and N and the Evolution of Ethylene and Production of GSH under HS
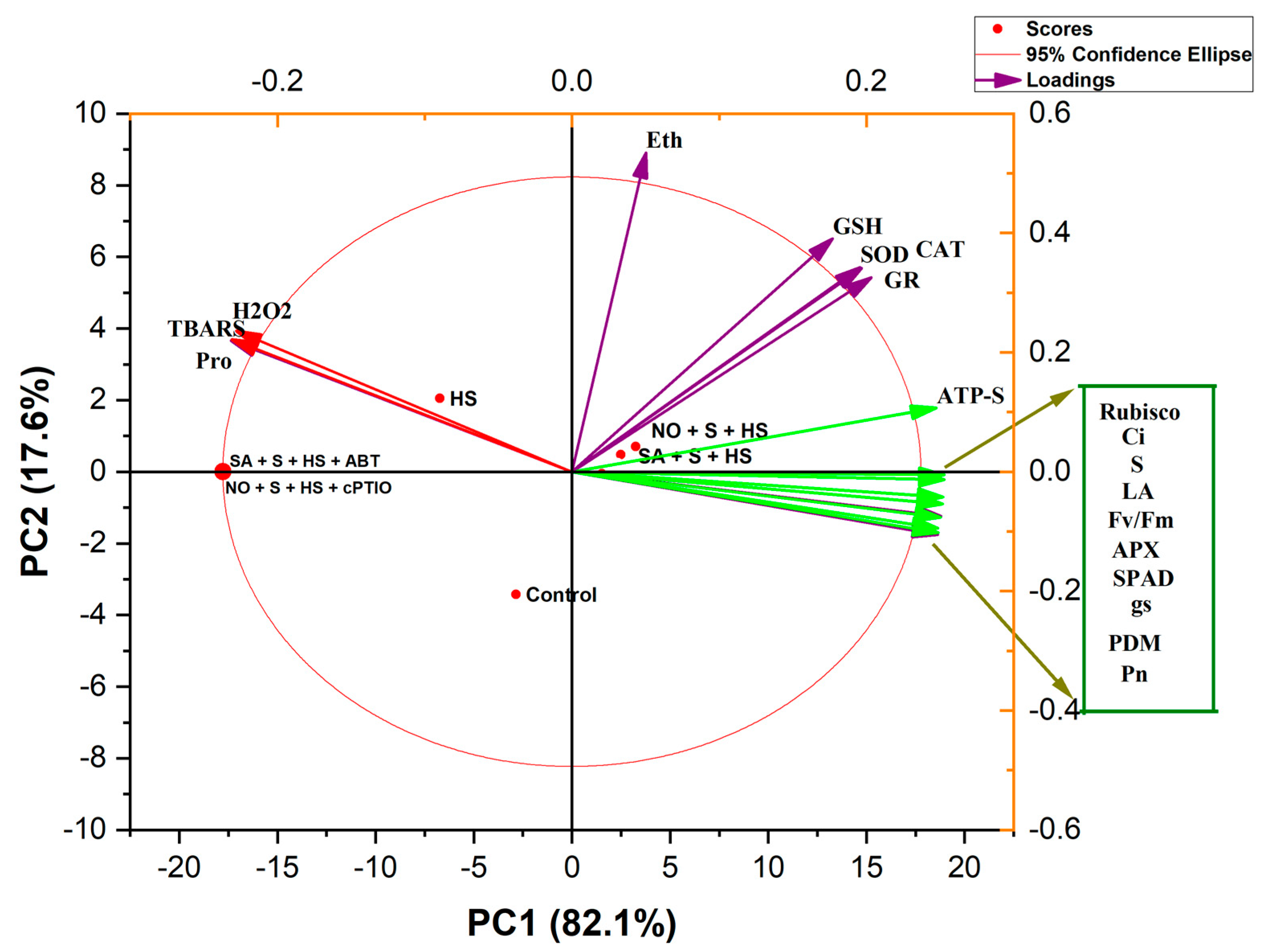
2.10. Principal Component Analysis (PCA)
3. Discussion
3.1. Screening of T. aestivum Cultivars for Their p-SUE under HS
3.2. Individual and Combined Roles of SA, NO and S in Alleviating HS
3.3. Confirmatory Experimental Results for the Involvement of SA (Using AIP) and NO (Using cPTIO) in Mitigating HS
4. Materials and Methods
4.1. Plant Culture and Treatments
4.2. Measurement of Growth Parameters, Photosynthetic Gas Exchange Parameters and Chlorophyll Content
4.3. Determination of H2O2 Content and Lipid Peroxidation
4.4. Determination of Leaf S Content, Assays of Antioxidant Enzyme, NR and ATP-S Activity
4.5. Determination of Photosynthetic SUE, GSH Content and Ethylene Production
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pawlak, K.; Kołodziejczak, M. The role of agriculture in ensuring food security in developing countries: Considerations in the context of the problem of sustainable food production. Sustainability 2020, 12, 5488. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 29, 9326–9331. [Google Scholar] [CrossRef]
- IPCC. Climate Change—The Physical Science Basis—Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2021. [Google Scholar]
- Gautam, H.; Fatma, M.; Sehar, Z.; Iqbal, N.; Albaqami, M.; Khan, N.A. Exogenously-sourced ethylene positively modulates photosynthesis, carbohydrate metabolism, and antioxidant defense to enhance heat tolerance in rice. Int. J. Mol. Sci. 2022, 23, 1031. [Google Scholar] [CrossRef]
- Gautam, H.; Sehar, Z.; Rehman, M.T.; Hussain, A.; Al Ajmi, M.F.; Khan, N.A. Nitric oxide enhances photosynthetic nitrogen and sulfur-use efficiency and activity of ascorbate-glutathione cycle to reduce high temperature stress-induced oxidative stress in rice (Oryza sativa L.) plants. Biomolecules 2021, 11, 305. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and HS: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Nisa, M.U.; Huang, Y.; Benhamed, M.; Raynaud, C. The plant DNA damage response: Signaling pathways leading to growth inhibition and putative role in response to stress conditions. Front. Plant Sci. 2019, 10, 653. [Google Scholar] [CrossRef]
- Tack, J.; Barkley, A.; Nalley, L.L. Effect of warming temperatures on US wheat yields. Proc. Natl. Acad. Sci. USA 2015, 2, 6931–9936. [Google Scholar] [CrossRef]
- Abhinandan, K.; Skori, L.; Stanic, M.; Hickerson, N.; Jamshed, M.; Samuel, M.A. Abiotic stress signaling in wheat–an inclusive overview of hormonal interactions during abiotic stress responses in wheat. Front. Plant Sci. 2018, 9, 734. [Google Scholar] [CrossRef]
- Challinor, A.J.; Watson, J.; Lobell, D.B.; Howden, S.M.; Smith, D.R.; Chhetri, N. A meta-analysis of crop yield under climate change and adaptation. Nat. Clim. Change 2014, 4, 287–291. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Change 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.J.; Khan, N.A.; Anjum, N.A.; Masood, A.; Khan, M.I. Mechanistic elucidation of salicylic acid and sulphur-induced defence systems, nitrogen metabolism, photosynthetic, and growth potential of mungbean (Vigna radiata) under salt stress. J. Plant Growth Regul. 2021, 40, 1000–1016. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Khan, N.A.; Corpas, F.J. Crosstalk between abscisic acid and nitric oxide under heat stress: Exploring new vantage points. Plant Cell Rep. 2021, 40, 1429–1450. [Google Scholar] [CrossRef] [PubMed]
- Jahan, B.; Rasheed, F.; Sehar, Z.; Fatma, M.; Iqbal, N.; Masood, A.; Anjum, N.A.; Khan, N.A. Coordinated role of nitric oxide, ethylene, nitrogen, and sulfur in plant salt stress tolerance. Stresses 2021, 1, 181–199. [Google Scholar] [CrossRef]
- Rasheed, F.; Anjum, N.A.; Masood, A.; Sofo, A.; Khan, N.A. The key roles of salicylic acid and sulfur in plant salinity stress tolerance. J. Plant Growth Regul. 2020, 30, 1891–1904. [Google Scholar] [CrossRef]
- Rasheed, F.; Sehar, Z.; Fatma, M.; Iqbal, N.; Masood, A.; Anjum, N.A.; Khan, N.A. Involvement of ethylene in reversal of salt stress by salicylic acid in the presence of sulfur in mustard (brassica juncea L.). J. Plant Growth Regul. 2021, 3, 3449–3466. [Google Scholar] [CrossRef]
- Ali, M.M.; Shafique, M.W.; Gull, S.; Naveed, W.A.; Javed, T.; Yousef, A.F.; Mauro, R.P. Alleviation of HS in tomato by exogenous application of sulfur. Horticulturae 2021, 7, 21. [Google Scholar] [CrossRef]
- Rajput, V.D.; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; Mandzhieva, S. Recent developments in enzymatic antioxidant defence mechanism in plants with special reference to abiotic stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef]
- Parankusam, S.; Adimulam, S.S.; Bhatnagar-Mathur, P.; Sharma, K.K. Nitric oxide (NO) in plant heat stress tolerance: Current knowledge and perspectives. Front. Plant Sci. 2017, 8, 1582. [Google Scholar] [CrossRef]
- Popova, L.; Tuan, T. Nitric oxide in plants: Properties, biosynthesis and physiological functions. Iran. J. Sci. Technol. A 2010, 34, 173–183. [Google Scholar]
- Cséplő, Á.; Zsigmond, L.; Andrási, N.; Baba, A.I.; Labhane, N.M.; Pető, A.; Rigó, G. The AtCRK5 protein kinase is required to maintain the ROS no balance affecting the PIN2-mediated root gravitropic response in arabidopsis. Int. J. Mol. Sci. 2021, 22, 5979. [Google Scholar] [CrossRef] [PubMed]
- Mishina, T.E.; Lamb, C.; Zeier, J. Expression of a nitric oxide degrading enzyme induces a senescence programme in Arabidopsis. Plant Cell Environ. 2007, 30, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Lamattina, L.; Spoel, S.H.; Loake, G.J. Nitric oxide function in plant biology: A redox cue in deconvolution. N. Phytol. 2014, 202, 1142–1156. [Google Scholar] [CrossRef]
- Iqbal, N.; Sehar, Z.; Fatma, M.; Umar, S.; Sofo, A.; Khan, N.A. Nitric oxide and abscisic acid mediate heat stress tolerance through regulation of osmolytes and antioxidants to protect photosynthesis and growth in wheat plants. Antioxidants 2022, 11, 372. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Ding, W.; Zhao, M.; Sun, B.; Zhang, L. Nitric oxide protects against oxidative stress under heat stress in the calluses from two ecotypes of reed. Plant Sci. 2006, 171, 449–458. [Google Scholar] [CrossRef]
- Yang, S.; Ulhassan, Z.; Shah, A.M.; Khan, A.R.; Azhar, W.; Hamid, Y.; Hussain, S.; Sheteiwy, M.S.; Salam, A.; Zhou, W. Salicylic acid underpins silicon in ameliorating chromium toxicity in rice by modulating antioxidant defense, ion homeostasis and cellular ultrastructure. Plant Physiol. Biochem. 2021, 166, 1001–1013. [Google Scholar] [CrossRef]
- Fatma, M.; Masood, A.; Per, T.S.; Rasheed, F.; Khan, N.A. Interplay between nitric oxide and sulfur assimilation in salt tolerance in plants. Crop J. 2016, 4, 153–161. [Google Scholar] [CrossRef]
- Hütsch, B.W.; Jahn, D.; Schubert, S. Grain yield of wheat (Triticum aestivum L.) under long-term heat stress is sink-limited with stronger inhibition of kernel setting than grain filling. J. Agron. Crop Sci. 2019, 205, 22–32. [Google Scholar] [CrossRef]
- Akasha, A.; Ashraf, M.; Shereen, A.; Mahboob, W.; Faisal, S. Heat tolerance screening studies and evaluating salicylic acid efficacy against high temperature in rice (Oryza sativa L.) genotypes. J. Plant Biochem Physiol. 2019, 7, 2. [Google Scholar]
- Mathur, S.; Agrawal, D.; Jajoo, A. Photosynthesis: Response to high temperature stress. J. Photochem. Photobiol. B Biol. 2014, 137, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Umar, S.; Khan, N.A.; Corpas, F.J. Nitric oxide and hydrogen sulfide coordinately reduce glucose sensitivity and decrease oxidative stress via ascorbate-glutathione cycle in heat-stressed wheat (Triticum aestivum L.) plants. Antioxidants 2021, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.L.; Liu, T.; Zhang, S.B.; Huang, W. Balancing light use efficiency and photoprotection in tobacco leaves grown at different light regimes. Environ Exp Bot. 2020, 175, 104–146. [Google Scholar] [CrossRef]
- Wang, Q.L.; Chen, J.H.; He, N.Y.; Guo, F.Q. Metabolic reprogramming in chloroplasts under heat stress in plants. Int. J. Mol. Sci. 2018, 19, 849. [Google Scholar] [CrossRef] [PubMed]
- Sita, K.; Sehgal, A.; Bhardwaj, A.; Bhandari, K.; Kumar, S.; Prasad, P.V.; Jha, U.; Siddique, K.H.; Nayyar, H. Nitric oxide secures reproductive efficiency in heat-stressed lentil (Lens culinaris Medik.) plants by enhancing the photosynthetic ability to improve yield traits. Physiol. Mol. Biol. Plants. 2021, 27, 2549–2566. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H. Nitric oxide alleviates the inhibition of salinity stress on seed germination and seedling growth of Cynanchum bungei Decne (Asclepiadaceae). Hort. Sci. 2015, 50, 119–122. [Google Scholar] [CrossRef]
- Chen, Y.E.; Mao, H.T.; Wu, N.; Ud Din, M.A.; Khan, M.A.; Zhang, H.Y.; Yuan, S. Salicylic acid protects photosystem II by alleviating photoinhibition in Arabidopsis thaliana under high light. Int. J. Mol. Sci. 2020, 21, 1229. [Google Scholar] [CrossRef] [PubMed]
- Jahan, M.S.; Wang, Y.; Shu, S.; Zhong, M.; Chen, Z.; Wu, J.; Guo, S. Exogenous salicylic acid increases the heat tolerance in Tomato (Solanum lycopersicum L.) by enhancing photosynthesis efficiency and improving antioxidant defense system through scavenging of reactive oxygen species. Sci. Hortic. 2019, 247, 421–429. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence-a practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Lu, C.M.; Zhang, J.H. Heat-induced multiple effects on PSII in wheat plants. J. Plant Physiol. 2000, 156, 259–265. [Google Scholar] [CrossRef]
- Camejo, D.; Rodríguez, P.; Morales, M.A.; Dell’Amico, J.M.; Torrecillas, A.; Alarcón, J.J. High temperature effects on photosynthetic activity of two tomato cultivars with different heat susceptibility. J. Plant Physiol. 2005, 162, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Resurreccion, A.P.; Makino, A.; Bennett, J.; Mae, T. Effects of sulfur nutrition on the growth and photosynthesis of rice. Soil Sci. Plant Nutr. 2001, 47, 611–620. [Google Scholar] [CrossRef]
- Lunde, C.; Zygadlo, A.; Simonsen, H.T.; Nielsen, P.L.; Blennow, A.; Haldrup, A. Sulfur starvation in rice: The effect on photosynthesis, carbohydrate metabolism, and oxidative stress protective pathways. Physiol. Plant. 2008, 134, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Kopriva, S. Regulation of sulfate assimilation in Arabidopsis and beyond. Ann. Bot. 2006, 97, 479–495. [Google Scholar] [CrossRef]
- Wangeline, A.L.; Burkhead, J.L.; Hale, K.L.; Lindblom, S.D.; Terry, N.; Pilon, M.; Pilon-Smits, E.A. Overexpression of ATP sulfurylase in Indian mustard: Effects on tolerance and accumulation of twelve metals. J. Environ. Qual. 2004, 33, 54–60. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Mahmud, J.A.; Nahar, K.; Mohsin, S.M.; Parvin, K.; Fujita, M. Interaction of sulfur with phytohormones and signaling molecules in conferring abiotic stress tolerance to plants. Plant Signal. Behav. 2018, 13, e1477905. [Google Scholar] [CrossRef]
- Nazar, R.; Umar, S.; Khan, N.A.; Sareer, O. Salicylic acid supplementation improves photosynthesis and growth in mustard through changes in proline accumulation and ethylene formation under drought stress. S. Afr. J. Bot. 2015, 98, 84–94. [Google Scholar] [CrossRef]
- Pal, U.R.; Gossett, D.R.; Sims, J.L.; Leggett, J.E. Molybdenum and sulfur nutrition effects on nitrate reduction in burley tobacco. Canad. J. Bot. 1976, 54, 2014–2022. [Google Scholar] [CrossRef]
- Sehar, Z.; Iqbal, N.; Fatma, M.; Rather, B.A.; Albaqami, M.; Khan, N.A. Ethylene suppresses abscisic acid, modulates antioxidant system to counteract arsenic-inhibited photosynthetic performance in the presence of selenium in mustard. Front. Plant Sci. 2022, 13, 852704. [Google Scholar] [CrossRef]
- Sehar, Z.; Gautam, H.; Iqbal, N.; Alvi, A.F.; Jahan, B.; Fatma, M.; Albaqami, M.; Khan, N.A. The functional interplay between ethylene, hydrogen sulfide and sulfur in plant heat stress tolerance. Biomolecules 2022, 12, 678. [Google Scholar] [CrossRef]
- Masood, A.; Khan, M.I.R.; Fatma, M.; Asgher, M.; Per, T.S.; Khan, N.A. Involvement of ethylene in gibberellic acid-induced sulfur assimilation, photosynthetic responses, and alleviation of cadmium stress in mustard. Plant Physiol. Biochem. 2016, 104, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wen, T.; Huang, Y.; Guan, Y.; Hu, J. Salicylic acid biosynthesis inhibitors increase chilling injury to maize (Zea mays L.) seedlings. Plant Growth Regul. 2018, 86, 11–21. [Google Scholar] [CrossRef]
- Song, L.; Zhao, H.; Hou, M. Involvement of nitric oxide in acquired thermotolerance of rice seedlings. Russ. J. Plant Physiol. 2013, 60, 785–790. [Google Scholar] [CrossRef]
- Kaya, C.; Okant, M.; Ugurlar, F.; Alyemeni, M.N.; Ashraf, M.; Ahmad, P. Melatonin-mediated nitric oxide improves tolerance to cadmium toxicity by reducing oxidative stress in wheat plants. Chemosphere 2019, 225, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Sehar, Z.; Masood, A.; Khan, N.A. Nitric oxide reverses glucose-mediated photosynthetic repression in wheat (Triticum aestivum L.) under salt stress. Environ. Exp. Bot. 2019, 151, 277–289. [Google Scholar] [CrossRef]
- Okuda, T.; Matsuda, Y.; Yamanaka, A.; Sagisaka, S. Abrupt increase in the level of hydrogen peroxide in leaves of winter wheat is caused by cold treatment. Plant Physiol. 1991, 97, 1265–1267. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–110. [Google Scholar] [CrossRef]
- Chesnin, L.; Yien, C.H. Turbidimetric determination of available sulfates. Soil Sci. Soc. Am. J. 1951, 15, 149–151. [Google Scholar] [CrossRef]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef]
- Kuo, T.M.; Warner, R.L.; Kleinhofs, A. In vitro stability of nitrate reductase from barley leaves. Phytochemistry 1982, 21, 531–533. [Google Scholar] [CrossRef]
- Nakagawa, H.; Poulle, M.; Oaks, A. Characterization of nitrate reductase from corn leaves (Zea mays cv W64A× W182E): Two molecular forms of the enzyme. Plant Physiol. 1984, 75, 285–289. [Google Scholar] [CrossRef] [PubMed]
- Lappartient, A.G.; Touraine, B. Demand-driven control of root ATP sulfurylase activity and SO42- uptake in intact canola (the role of phloem-translocated glutathione). Plant Physiol. 1996, 111, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.E. Determination of glutathione and glutathione disulfide in biological samples. In Methods in Enzymolology; Academic Press: Cambridge, MA, USA, 1985; Volume 113, pp. 548–555. [Google Scholar]
- Fatma, M.; Iqbal, N.; Gautam, H.; Sehar, Z.; Sofo, A.; D’Ippolito, I.; Khan, N.A. Ethylene and sulfur coordinately modulate the antioxidant system and ABA accumulation in mustard plants under salt stress. Plants 2021, 10, 180. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Treatments | p-SUE (g m−2) | Plant Dry Weight (g plant−1) | Net Photosynthesis (μmol CO2 m−2 s−1) |
|---|---|---|---|---|
| HD-3226 | Control | 13.5 ± 0.6 a | 1.19 ± 0.05 a | 16.8 ± 0.8 a |
| HS | 07.1 ± 0.3 d | 0.88 ± 0.04 g | 13.0 ± 0.6 d | |
| HD-3237 | Control | 12.9 ± 0.6 b | 1.13 ± 0.05 b | 14.5 ± 0.7 b |
| HS | 06.4 ± 0.3 e | 0.82 ± 0.04 h | 10.8 ± 0.5 g | |
| HD-3271 | Control | 12.6 ± 0.6 bc | 1.11 ± 0.05 c | 13.6 ± 0.6 c |
| HS | 06.0 ± 0.3 ef | 0.78 ± 0.03 hi | 09.9 ± 0.4 h | |
| HD-3059 | Control | 11.2 ± 0.5 cd | 1.05 ± 0.05 e | 11.7 ± 0.5 f |
| HS | 05.1 ± 0.2 fg | 0.72 ± 0.03 ij | 08.2 ± 0.4 i | |
| HD-3090 | Control | 10.9 ± 0.5 cd | 1.01 ± 0.05 f | 11.1 ± 0.5 g |
| HS | 04.8 ± 0.2 g | 0.68 ± 0.03 j | 07.1 ± 0.3 j | |
| HI-1620 | Control | 11.7 ± 0.5 c | 1.07 ± 0.05 d | 12.4 ± 0.6 e |
| HS | 05.5 ± 0.2 f | 0.75 ± 0.03 i | 08.7 ± 0.4 i |
| Treatments | Chlorophyll Content (SPAD Value) | Net Photosynthesis (µmol CO2 m−2 s−1) | Stomatal Conductance (mmol CO2 m−2 s−1) | Intercellular CO2 Concentration (µmol CO2 mol−1) | Leaf Area (cm2 Plant−1) | Plant Dry Weight (g Plant−1) | Maximum Quantum Efficiency of PS II |
|---|---|---|---|---|---|---|---|
| Control | 33.7 ± 1.2 j | 10.7 ± 0.3 i | 403 ± 9.3 g | 237 ± 5.5 f | 24.7 ± 0.3 i | 0.87 ± 0.05 f | 0.80 ± 0.04 e |
| HS | 23.3 ± 0.9 k | 5.7 ± 0.2 j | 292 ± 7.2 h | 147 ± 3.7 h | 16.7 ± 0.2 j | 0.47± 0.03 g | 0.62 ± 0.05 f |
| S | 38.8 ± 1.2 g | 13.0 ± 0.5 f | 465 ± 11.2 de | 280 ± 6.2 de | 30.6 ± 0.3 f | 1.01 ± 0.06 e | 0.85 ± 0.03 cd |
| SA | 39.7 ± 1.2 f | 13.2 ± 0.6 ef | 470 ± 11.7 d | 289 ± 6.5 d | 32.3 ± 0.4 e | 1.03 ± 0.06 de | 0.88 ± 0.06 c |
| NO | 40.1 ± 1.3 e | 13.5 ± 0.6 e | 476 ± 12.2 cd | 315 ± 7.1 cd | 33.7 ± 0.4 de | 1.06 ± 0.07 d | 0.89 ± 0.30 c |
| SA + S | 41.9 ± 1.3 b | 14.7 ± 0.6 b | 487 ± 12.5 b | 330 ± 7.2 bc | 36.2 ± 0.4 b | 1.15 ± 0.13 b | 0.95 ± 0.05 b |
| NO + S | 42.7 ± 1.3 a | 15.3 ± 0.7 a | 493 ± 12.8 a | 355 ± 7.7 a | 38.1 ± 0.5 a | 1.18 ± 0.13 a | 0.97 ± 0.09 a |
| S + HS | 35.2 ± 1.2 i | 11.4 ± 0.4 g | 425 ± 9.7 f | 208 ± 6.3 g | 26.3 ± 0.2 h | 0.95 ± 0.06 e | 0.82 ± 0.07 d |
| SA + HS | 35.5 ± 1.2 i | 11.6 ± 0.4 g | 432 ± 10.2 ef | 225 ± 5.3 fg | 26.8 ± 0.2 h | 0.97 ± 0.06 e | 0.82 ± 0.05 d |
| NO + HS | 36.2 ± 1.2 h | 12.8 ± 0.4 h | 443 ± 11.3 e | 260 ± 5.7 e | 27.5 ± 0.2 g | 1.03 ± 0.06 de | 0.85 ± 0.02 cd |
| SA + S + HS | 40.7 ± 1.2 d | 13.9 ± 0.5 d | 480 ± 12.1 c | 281 ± 5.8 c | 34.3 ± 0.4 d | 1.10 ± 0.11 de | 0.92 ± 0.03 bc |
| NO+ S + HS | 41.2 ± 1.3 c | 14.3 ± 0.6 c | 483 ± 12.2 bc | 312 ± 6.8 b | 35.7 ± 0.4 c | 1.12 ± 0.11 c | 0.94 ± 0.07 b |
| Treatments | Chlorophyll Content (SPAD value) | Net Photosynthesis (µmol CO2 m−2 s−1) | Stomatal Conductance (mmol CO2 m−2 s−1) | Intercellular CO2 Concentration (µmol CO2 mol−1) | Leaf Area (cm2 Plant−1) | Plant Dry Weight (g Plant−1) | Maximum Quantum Efficiency of PS II |
|---|---|---|---|---|---|---|---|
| Control | 30.4 ± 1.0 d | 13.2 ± 0.6 d | 385 ± 19.2 e | 212 ± 10.6 d | 27.3 ± 1.3 e | 0.80 ± 0.04 d | 0.82 ± 0.04 e |
| HS | 18.9 ± 0.9 e | 6.5 ± 0.3 e | 267 ± 13.3 f | 132 ± 06.6 e | 17.3 ± 0.8 f | 0.37 ± 0.01 e | 0.58 ± 0.02 f |
| SA + S + HS | 37.3 ± 1.8 b | 17.7 ± 0.8 b | 476 ± 23.8 b | 296 ± 14.8 b | 36.5 ± 1.8 c | 1.06 ± 0.05 b | 0.99 ± 0.05 c |
| NO + S + HS | 38.7 ± 1.9 a | 18.5 ± 0.9 a | 489 ± 24.4 a | 308 ± 15.4 a | 38.7 ± 1.9 a | 1.10 ± 0.05 a | 1.03 ± 0.05 a |
| SA + S + HS + AIP | 35.7 ± 1.7 c | 16.3 ± 0.8 cd | 458 ± 22.9 d | 284 ± 14.2 c | 34.6 ± 1.7 d | 1.00 ± 0.05 c | 0.96 ± 0.05 d |
| NO + S + HS + cPTIO | 37.5 ± 1.8 b | 16.8 ± 0.8 c | 472 ± 23.6 c | 291 ± 14.5 bc | 37.4 ± 1.8 b | 1.05 ± 0.05 b | 1.01 ± 0.05 b |
| Treatments | H2O2 Content | TBARS Content | SOD Activity | APX Activity | GR Activity |
|---|---|---|---|---|---|
| (nmol g−1 FW) | (U min−1 mg−1 Protein) | ||||
| Control | 13.5 ± 0.6 b | 5.3 ± 0.2 b | 7.22 ± 0.3 f | 2.7 ± 0.1 f | 2.42 ± 0.1 e |
| HS | 30.5 ± 1.5 a | 17.1 ± 0.8 a | 12.90 ± 0.6 e | 5.3 ± 0.2 e | 4.22 ± 0.2 d |
| SA + S + HS | 10.4 ± 0.5 e | 3.5 ± 0.1 d | 19.70 ± 0.9 b | 8.5 ± 0.4 c | 6.70 ± 0.3 b |
| NO + S + HS | 9.2 ± 0.4 f | 3.2 ± 0.1 de | 21.40 ± 1.0 a | 9.5 ± 0.4 a | 7.00 ± 0.3 a |
| SA + S + HS + AIP | 11.4 ± 0.5 c | 3.9 ± 0.1 c | 16.80 ± 0.8 d | 7.8 ± 0.3 d | 6.10 ± 0.3 c |
| NO + S + HS + cPTIO | 10.7 ± 0.5 d | 3.7 ± 0.1 cd | 18.50 ± 0.9 c | 8.8 ± 0.4 b | 6.50 ± 0.3 bc |
| Treatments | ATP-S Activity (µmol g−1 protein s−1) | Sulfur Content (mg g−1 DW) | p-SUE | GSH Content (nmol g−1 FW) | Ethylene Production (ng kg−1 FW s−1) |
|---|---|---|---|---|---|
| Control | 2.6 ± 0.3 d | 4.9 ± 0.2 d | 10.9 ± 0.5 e | 284 ± 14.2 f | 24.7 ± 1.2 e |
| HS | 1.9 ± 0.1 e | 2.8 ± 0.1 e | 05.6 ± 0.2 f | 303 ± 15.1 e | 70.8 ± 3.5 a |
| SA + S + HS | 6.0 ± 0.3 b | 7.7 ± 0.3 bc | 13.7 ± 0.6 a | 418 ± 20.9 b | 63.7 ± 3.1 cd |
| NO + S + HS | 6.3 ± 0.3 a | 8.0 ± 0.4 a | 14.9 ± 0.7 a | 427 ± 21.3 a | 65.8 ± 3.2 b |
| SA + S + HS + AIP | 5.7 ± 0.2 c | 7.3 ± 0.3 c | 12.6 ± 0.6 d | 371 ± 18.5 d | 60.3 ± 3.0 d |
| NO + S + HS + cPTIO | 6.1 ± 0.3 bc | 7.8 ± 0.3 c | 13.4 ± 0.6 c | 387 ± 19.3 c | 63.9 ± 3.1 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasheed, F.; Mir, I.R.; Sehar, Z.; Fatma, M.; Gautam, H.; Khan, S.; Anjum, N.A.; Masood, A.; Sofo, A.; Khan, N.A. Nitric Oxide and Salicylic Acid Regulate Glutathione and Ethylene Production to Enhance Heat Stress Acclimation in Wheat Involving Sulfur Assimilation. Plants 2022, 11, 3131. https://doi.org/10.3390/plants11223131
Rasheed F, Mir IR, Sehar Z, Fatma M, Gautam H, Khan S, Anjum NA, Masood A, Sofo A, Khan NA. Nitric Oxide and Salicylic Acid Regulate Glutathione and Ethylene Production to Enhance Heat Stress Acclimation in Wheat Involving Sulfur Assimilation. Plants. 2022; 11(22):3131. https://doi.org/10.3390/plants11223131
Chicago/Turabian StyleRasheed, Faisal, Iqbal R. Mir, Zebus Sehar, Mehar Fatma, Harsha Gautam, Sheen Khan, Naser A. Anjum, Asim Masood, Adriano Sofo, and Nafees A. Khan. 2022. "Nitric Oxide and Salicylic Acid Regulate Glutathione and Ethylene Production to Enhance Heat Stress Acclimation in Wheat Involving Sulfur Assimilation" Plants 11, no. 22: 3131. https://doi.org/10.3390/plants11223131
APA StyleRasheed, F., Mir, I. R., Sehar, Z., Fatma, M., Gautam, H., Khan, S., Anjum, N. A., Masood, A., Sofo, A., & Khan, N. A. (2022). Nitric Oxide and Salicylic Acid Regulate Glutathione and Ethylene Production to Enhance Heat Stress Acclimation in Wheat Involving Sulfur Assimilation. Plants, 11(22), 3131. https://doi.org/10.3390/plants11223131