

Sideritis elica, a New Species of Lamiaceae from Bulgaria, Revealed by Morphology and Molecular Phylogeny


Abstract
1. Introduction
2. Results
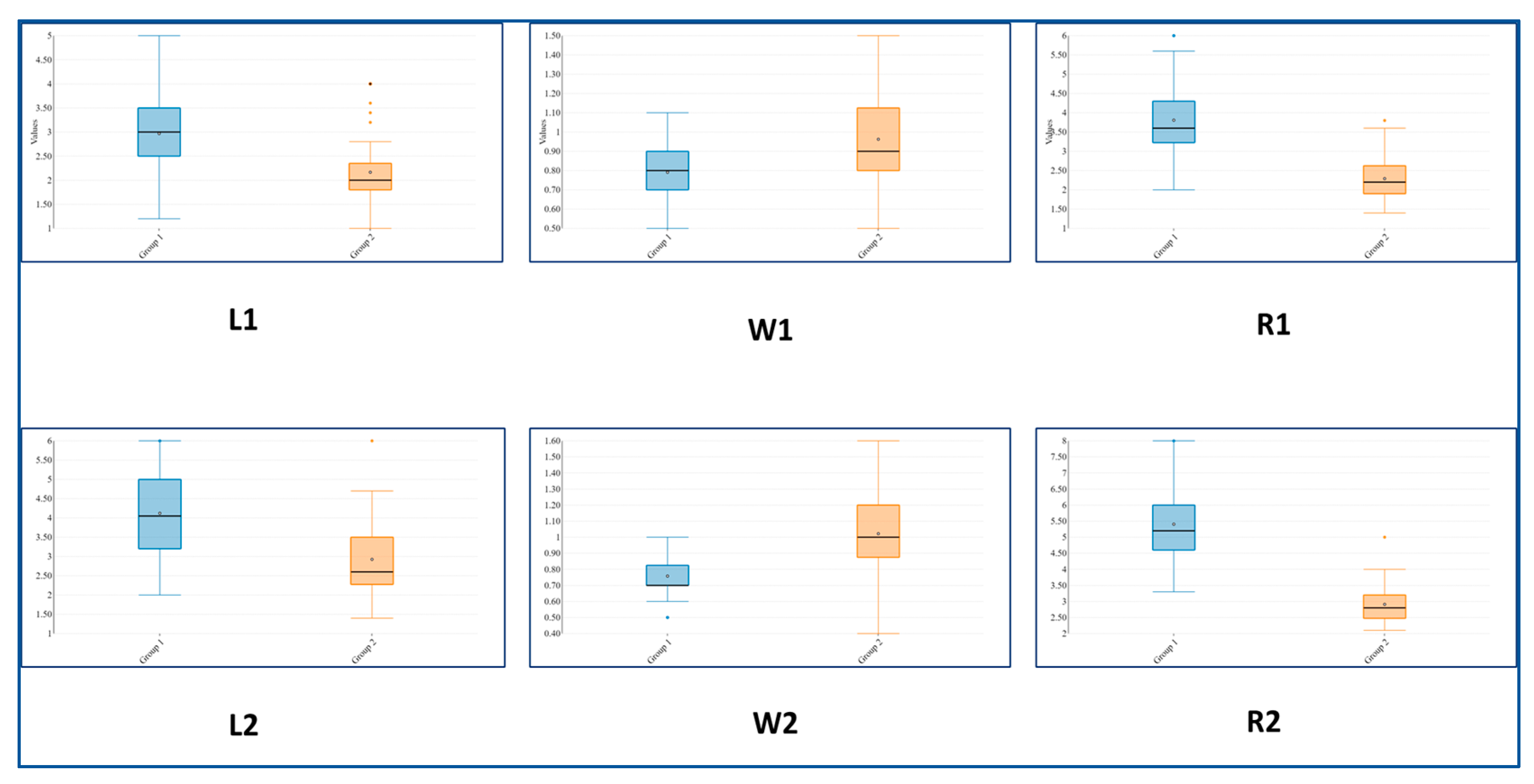
2.1. Morphological and Morphometric Data
2.2. DNA Barcoding
2.3. Taxonomic Implications
3. Sideritis elica Aneva, Zhelev and Bonchev sp. nova
4. Materials and Methods
DNA Extraction, PCR Amplification, and Sequencing
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barber, J.C.; Francisco-Ortega, J.; Santos-Guerra, A.; Marrero, A.; Jansen, R.K. Evolution of endemic Sideritis (Lamiaceae) in Macaronesia: Insight from chloroplast DNA restriction site analysis. Syst. Bot. 2000, 25, 633–647. [Google Scholar] [CrossRef]
- Barber, J.C.; Francisco-Ortega, J.; Santos-Guerra, A.; Turner, K.G.; Jansen, R.K. Origin of Macaronesian Sideritis L. Mol. Phylogenet. Evol. 2002, 23, 293–306. [Google Scholar] [CrossRef]
- Huber-Morath, A. Sideritis L. In Flora of Turkey and the East Aegean Islands; Davis, P.H., Ed.; Edinburgh University Press: Edinburgh, UK, 1982; Volume 7, pp. 178–199. [Google Scholar]
- Heywood, V. Sideritis L. In Flora Europaea; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1972; Volume 3, pp. 134–138. [Google Scholar]
- Baden, C. Sideritis L. In The Mountain Flora of Greece; Strid, A., Tan, K., Eds.; Edinburgh University Press: Edinburgh, UK, 1991; Volume 2, pp. 84–91. [Google Scholar]
- Koutsos, T.V.; Chatzopoulou, P.S. Sideritis species in Greece: The current situation. In Report of a Working Group on Medicinal and Aromatic Plants. Second and Third Meeting; Lipman, E., Ed.; Bioversity International: Rome, Italy, 2009; pp. 112–114. [Google Scholar]
- Papanikolaou, K.; Kokkini, S. A taxonomic revision of Sideritis L. section Empedoclia (Rafin.) Benntham (Labiatae) in Greece. In Aromatic Plants: Basic and Applied Aspects; Margaris, N., Koedam, A., Voukou, D., Eds.; Martinus Nijhof Publublishing: The Hague, The Netherlands, 1982; pp. 101–128. [Google Scholar]
- Assenov, I. Sideritis L. In Flora of PR Bulgaria; Velchev, V., Ed.; Bulgarian Academy of Sciences Publishing House: Sofia, Bulgaria, 1989; Volume 9, pp. 369–374. [Google Scholar]
- Evstatieva, L. Sideritis scardica Griseb. In Red Data Book of Bulgaria. Vol. 1. Plants and Fungi; Peev, D., Petrova, A.S., Anchev, M., Temniskova, D., Denchev, C.M., Ganeva, A., Gussev, C., Vladimirov, V., Eds.; IBER-BAS and MoEW: Sofia, Bulgaria, 2015; p. 607. [Google Scholar]
- Evstatieva, L. Sideritis syriaca L. In Red Data Book of Bulgaria. Vol. 1. Plants and Fungi; Peev, D., Petrova, A.S., Anchev, M., Temniskova, D., Denchev, C.M., Ganeva, A., Gussev, C., Vladimirov, V., Eds.; IBER-BAS and MoEW: Sofia, Bulgaria, 2015; p. 319. [Google Scholar]
- Stoyanov, S. Sideritis lanata L. In Red Data Book of Bulgaria. Vol. 1. Plants and Fungi; Peev, D., Petrova, A.S., Anchev, M., Temniskova, D., Denchev, C.M., Ganeva, A., Gussev, C., Vladimirov, V., Eds.; IBER-BAS and MoEW: Sofia, Bulgaria, 2015; p. 606. [Google Scholar]
- Davis, P.H.; Mill, R.R.; Tan, K. Flora of Turkey and the East Aegean Islands (Supplement); Edinburgh University Press: Edinburgh, UK, 1988; Volume 10, p. 590. [Google Scholar]
- Aytaç, Z.; Aksoy, A. A new Sideritis species (Labiatae) from Turkey. Flora Mediterr. 2000, 10, 181–184. [Google Scholar]
- Evstatieva, L. A review of the cultivation of endangered medicinal plants in Bulgaria. Ann. Sofia Univ. Fac. Biol. Book 2–Bot. 2005, 97, 45–52. [Google Scholar]
- Aneva, I.; Zhelev, P. Morphometric studies of Sideritis scardica Grsb. and S. syriaca L. in their natural populations in Bulgaria. Boletín Latinoam. Caribe De Plantas Med. Aromáticas 2019, 18, 71–80. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Kress, W.J.; Wurdack, K.J.; Zimmer, E.A.; Weigt, L.A.; Janzen, D.H. Use of DNA barcodes to identify flowering plants. Proc. Nat. Acad. Sci. USA 2005, 102, 8369–8374. [Google Scholar] [CrossRef]
- Hollingsworth, P.M.; Graham, S.W.; Little, D.P. Choosing and using a plant DNA barcode. PLoS ONE 2011, 6, e19254. [Google Scholar] [CrossRef]
- Hu, H.H.; Liu, B.; Liang, Y.S.; Ye, J.F.; Saqib, S.; Meng, Z.; Lu, L.M.; Chen, Z.D. An updated Chinese vascular plant tree of life: Phylogenetic diversity hotspots revisited. J. Syst. Evol. 2020, 58, 663–672. [Google Scholar] [CrossRef]
- Ullah, F.; Gao, Y.; Sari, İ.; Jiao, R.F.; Saqib, S.; Gao, X.F. Macro-Morphological and Ecological Variation in Rosa sericea Complex. Agronomy 2022, 12, 1078. [Google Scholar] [CrossRef]
- Struck, T.H.; Feder, J.L.; Bendiksby, M.; Birkeland, S.; Cerca, J.; Gusarov, V.I.; Kistenich, S.; Larsson, K.-H.; Liow, L.H.; Nowak, M.D.; et al. Finding evolutionary processes hidden in cryptic species. Trends Ecol. Evol. 2018, 33, 153–163. [Google Scholar] [CrossRef]
- Carstens, B.C.; Satler, J.D. The carnivorous plant described as Sarracenia alata contains two cryptic species. Biol. J. Linn. Soc. 2013, 109, 737–746. [Google Scholar] [CrossRef]
- Heylen, O.C.G.; Debortoli, N.; Marescaux, J.; Olofsson, J.K. A revised phylogeny of the Mentha spicata clade reveals cryptic species. Plants 2021, 10, 819. [Google Scholar] [CrossRef]
- Vigalondo, B.; Fernández-Mazuecos, M.; Vargas, P.; Sáez, L. Unmasking cryptic species: Morphometric and phylogenetic analyses of the Ibero-North African Linaria incarnata complex. Bot. J. Linn. Soc. 2015, 177, 395–417. [Google Scholar] [CrossRef]
- De Mattia, F.; Bruni, I.; Galimberti, A.; Cattaneo, F.; Casiraghi, M.; Labra, M. A comparative study of different DNA barcoding markers for the identification of some members of Lamiacaeae, Food Res. Int. 2011, 44, 693–702. [Google Scholar]
- Theodoridis, S.; Stefanaki, A.; Tezcan, M.; Aki, C.; Kokkini, S.; Vlachonasios, K.E. DNA barcoding in native plants of the Labiatae (Lamiaceae) family from Chios Island (Greece) and the adjacent Çeşme-Karaburun Peninsula (Turkey). Mol. Ecol. Resour. 2012, 12, 620–633. [Google Scholar] [CrossRef]
- Nazar, N.; Howard, C.; Slater, A.; Sgamma, T. Challenges in Medicinal and Aromatic Plants DNA Barcoding—Lessons from the Lamiaceae. Plants 2022, 11, 137. [Google Scholar] [CrossRef]
- Kalivas, A.; Ganopoulos, I.; Xanthopoulou, A.; Chatzopoulou, P.; Tsaftaris, A.; Madesis, P. DNA barcode ITS2 coupled with high resolution melting (HRM) analysis for taxonomic identification of Sideritis species growing in Greece. Mol. Biol. Rep. 2014, 41, 5147–5155. [Google Scholar] [CrossRef]
- Grdiša, M.; Radosavljević, I.; Liber, Z.; Stefkov, G.; Ralli, P.; Chatzopoulou, P.S.; Carović-Stanko, K.; Šatović, Z. Divergent selection and genetic structure of Sideritis scardica populations from southern Balkan Peninsula as revealed by AFLP fingerprinting. Sci. Rep. 2019, 9, 12767. [Google Scholar] [CrossRef]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | Sideritis scardica | Sideritis elica |
|---|---|---|
| Stem height | 20–30 (40) cm | 40–50 (60) cm |
| Stem pubescence | Densely pubescent | Densely white lanate |
| Length of the upper leaves (cm) | 3–3.5 | 2–2.5 |
| Width of the upper leaves (cm) | ||
| Ratio L:W of the upper 1–3 pairs of leaves | 3–6 | 2–3 |
| Leaf indumentum | Pubescent; gray and white trichomes | Densely lanate; white trichomes |
| Leaf color (mature leaves) | Grayish green | White; lanate |
| Barcoding Markers | ||||
|---|---|---|---|---|
| rbcL | trnH-psbA | |||
| WP | ||||
| D | SE | D | SE | |
| Chervenata Stena | 0.00 | 0.00 | 0.00 | 0.00 |
| Slavyanka Mountain | 0.00 | 0.00 | 0.001 | 0.001 |
| BP | ||||
| 0.002 | 0.002 | 0.018 | 0.006 | |
| Barcode Region | Primers | Primer Sequences 5′-3′ | PCR Conditions |
|---|---|---|---|
| matK | matK-1RKIM-f | ACCCAGTCCATCTGGAAATCTTGGTTC | 95 °C 5 min (95 °C 30 s, 51 °C 50 s, 72 °C 1 min)—35 cycles 72 °C 7 min |
| matK-3FKIM-r | CGTACAGTACTTTTGTGTTTACGAG | ||
| rbcL | rbcLa-F | ATGTCACCACAAACAGAGACTAAAGC | 94 °C 4 min (94 °C 30 s, 55 °C 30 s, 72 °C 1 min)—35 cycles 72 °C 10 min |
| rbcLajf634R | GAAACGGTCTCTCCAACGCAT | ||
| trnH-psbA | trnH-F | CGCGCATGGTGGATTCACAATCC | 94 °C 4 min (94 °C 30 s, 55 °C 30 s, 72 °C 1 min)—35 cycles 72 °C 7 min |
| psbA3_r | GTTATGCATGAACGTAATGCTC | ||
| ITS | ITS F1 | CCTTATCATTTAGAGGAAGGAG | 94 °C 5 min (94 °C 30 s, 50 °C 30 s, 72 °C 1 min)—35 cycles 72 °C 5 min |
| ITS 4 | TCCTCCGCTTATTGATATGC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aneva, I.; Zhelev, P.; Bonchev, G. Sideritis elica, a New Species of Lamiaceae from Bulgaria, Revealed by Morphology and Molecular Phylogeny. Plants 2022, 11, 2900. https://doi.org/10.3390/plants11212900
Aneva I, Zhelev P, Bonchev G. Sideritis elica, a New Species of Lamiaceae from Bulgaria, Revealed by Morphology and Molecular Phylogeny. Plants. 2022; 11(21):2900. https://doi.org/10.3390/plants11212900
Chicago/Turabian StyleAneva, Ina, Petar Zhelev, and Georgi Bonchev. 2022. "Sideritis elica, a New Species of Lamiaceae from Bulgaria, Revealed by Morphology and Molecular Phylogeny" Plants 11, no. 21: 2900. https://doi.org/10.3390/plants11212900
APA StyleAneva, I., Zhelev, P., & Bonchev, G. (2022). Sideritis elica, a New Species of Lamiaceae from Bulgaria, Revealed by Morphology and Molecular Phylogeny. Plants, 11(21), 2900. https://doi.org/10.3390/plants11212900