
Variability and Nativeness in the Mediterranean Taxa: Divergence and Phylogeography of Genista etnensis (Fabaceae) Inferred from Nuclear and Plastid Data

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction
2.2. Molecular Markers Selection
2.3. DNA Sequencing
2.4. Data Analysis

{kind=link}
{kind=link}
{kind=link}
| Code | Locus | (Name) Primer Sequence 5′-3′ | Ta (°C) | Ref. |
|---|---|---|---|---|
| cpDNA | ||||
| Pr1 | trnQ(UUG)-psbK IGS | (F: trnQ-IGSR)§ ACC CGT TGC CTT ACC GCT TGG (R: psbK-IGSR)§ ATC GAA AAC TTG CAG CAG CTT G | 58 | 1 |
| Pr8 | trnS(UGA)-psbZ IGS | (F: psbZ-IGS) AAT AGC CAA TTG AAA AGC (R: trnS_UGA-IGSR)§ ATC AAC CAC TCG GCC ATC | 55 | 1 |
| Pr11 | ycf3 intron-1 | (F: ycf3-E1F)§ CAT TTA CCT ATT ACA GAG ATG G (R: ycf3-E2R) TTC CGC GTA ATT TCC TTC | 50 | 1 |
| Pr18 | trnV(UAC)-atpE IGS | (F: atpE-IGSF)§ AGT GAC ATT GAT CCR CAA GAA GC (R: trnV_UAC-E1R) GTG TAA ACG AGG TGC TCT AC | 57 | 1 |
| Pr38 | psbA-trnH(GUG) IGS | (F: psbA3f) GTT ATG CAT GAA CGT AAT GCT C (R: trnHf)§ CGC GCA TGG TGG ATT CAC AATCC | 55 | 2, 3 |
| nrDNA | ||||
| N43 | ITS1+5.8S+ITS2 | (F: JK14)§ GGA GAA GTC GTA ACA AGG TTT CCG (R: SN3)§ TTC GCT CGC CGT TAC TAA GGG | 55 | 4, 5 |
3. Results
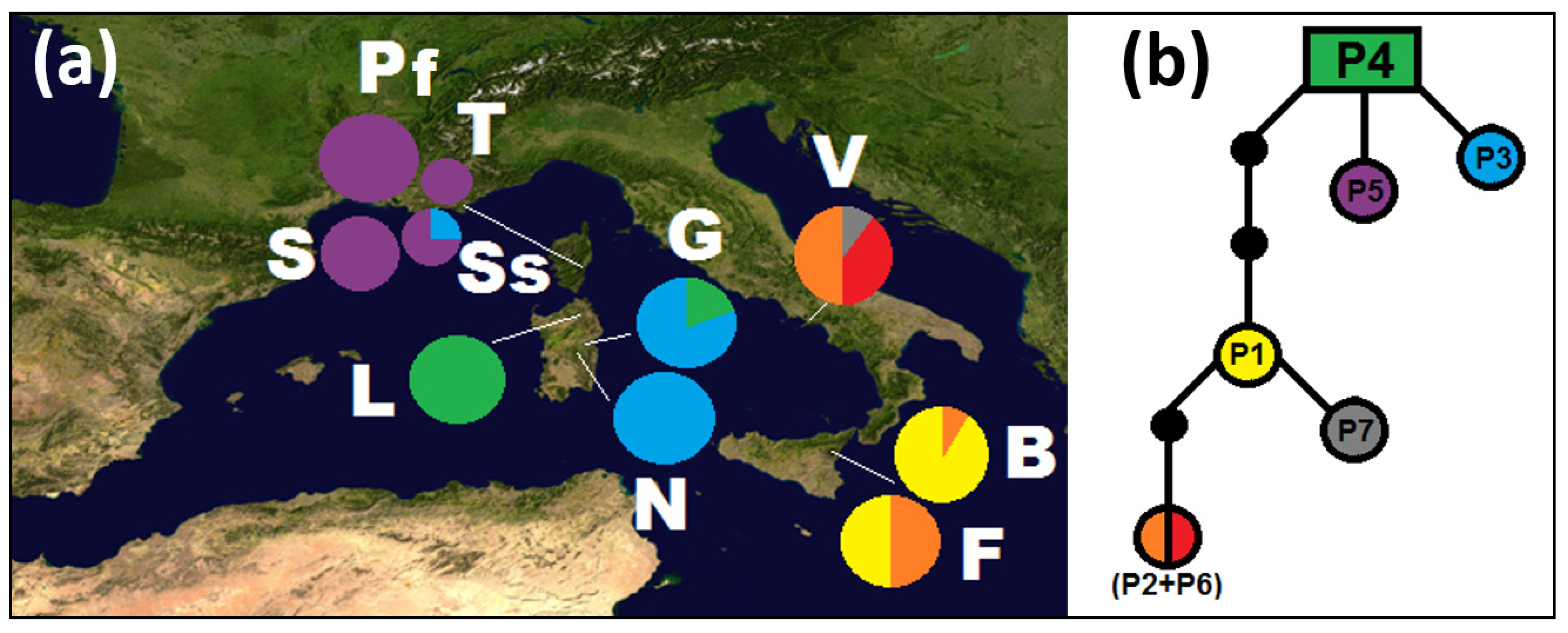
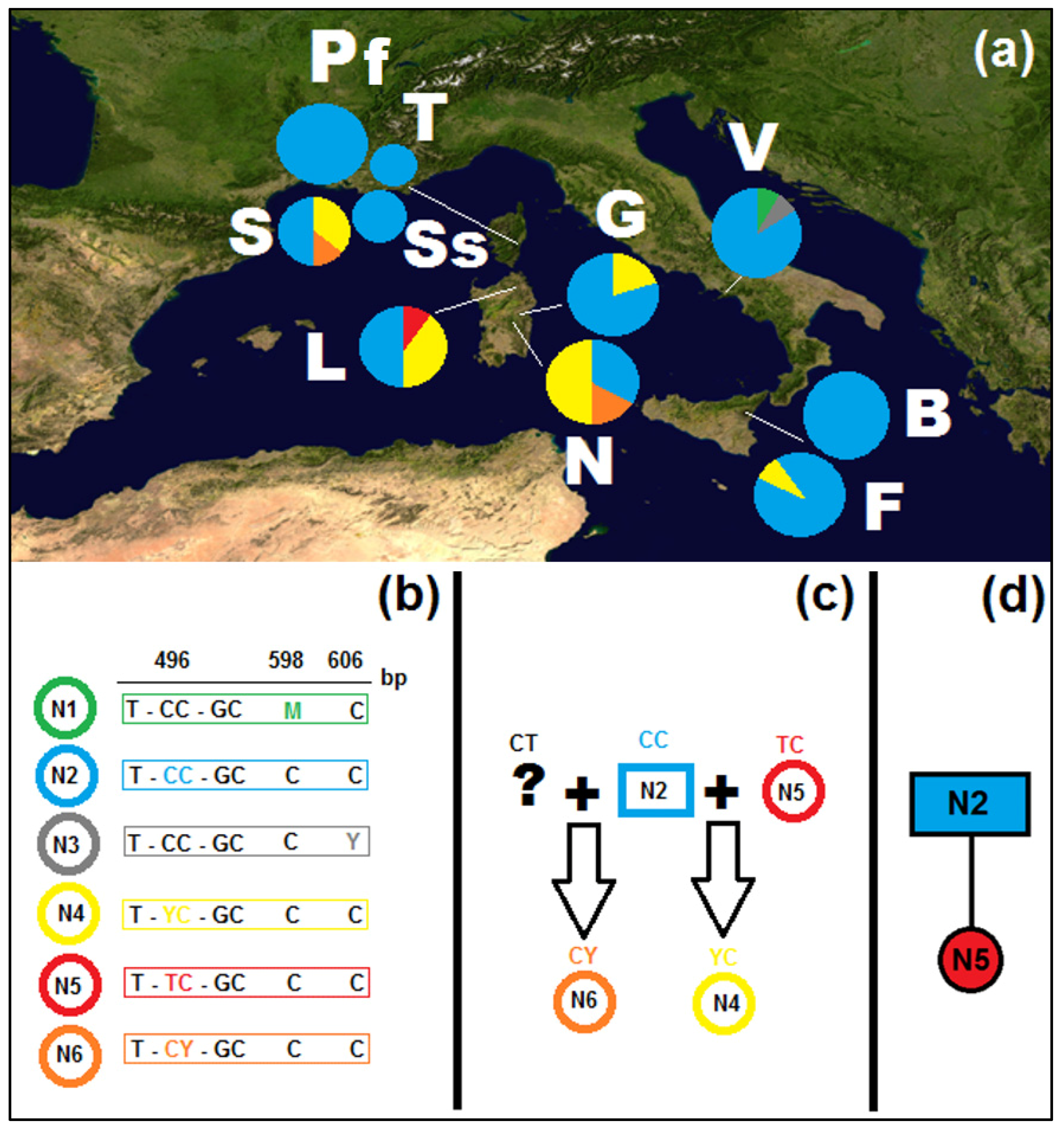
3.1. Plastid Sequences
3.2. Nuclear Sequences
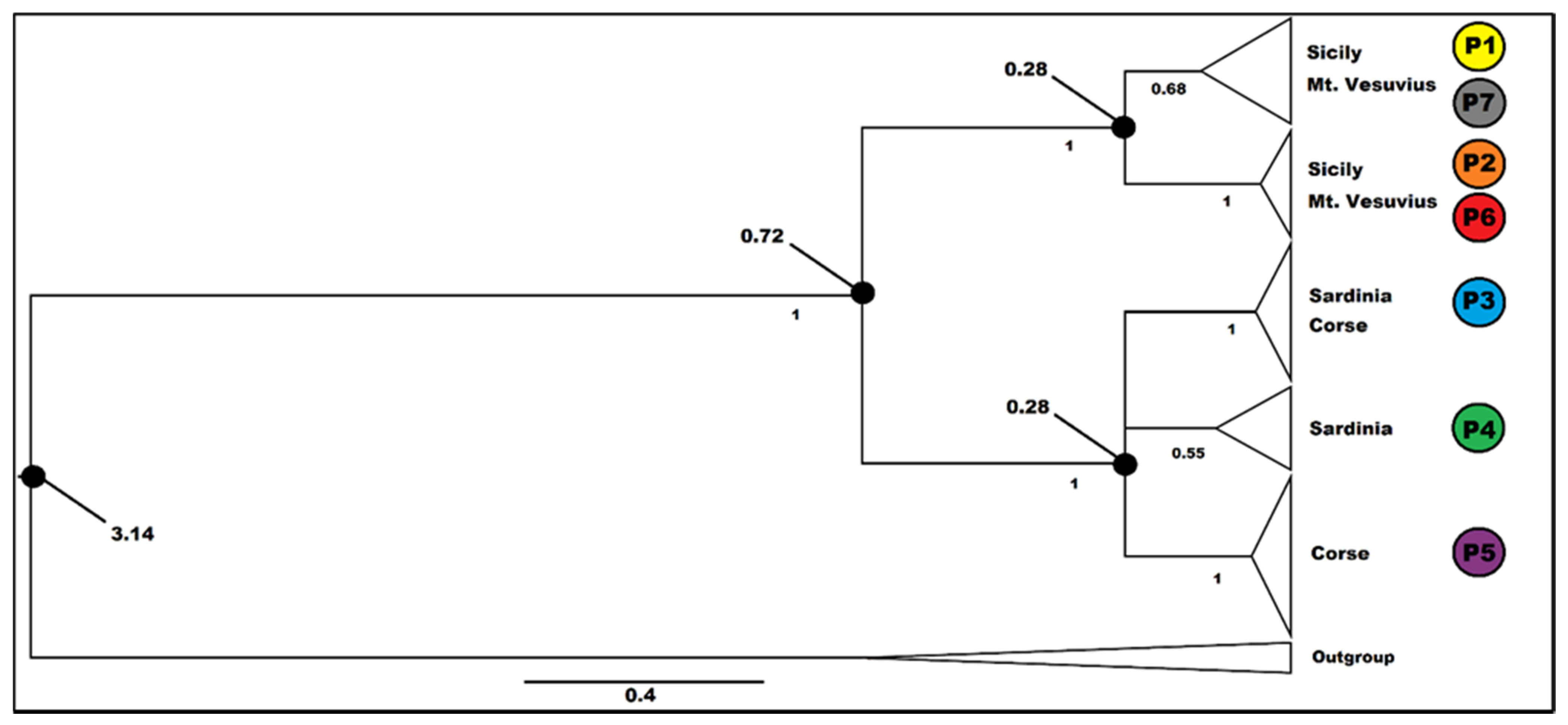
3.3. Molecular Dating
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO. “Plants of the World Online”. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 7 March 2021).
- Gibbs, P.E. A revision of the genus Genista L. Notes Roy. Bot. Gard. Edinb. 1966, 27, 11–99. [Google Scholar]
- ILDIS. International Legume Database & Information Service. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 7 March 2021).
- Duran, A.; Dural, H. Genista vuralii (Fabaceae), a new species from Turkey. Ann. Bot. Fenn. 2003, 40, 113–116. [Google Scholar]
- Gibbs, P.E. Genista L. In Flora Europaea 2; Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M., Valentine, D.H., Walters, S.M., Webb, D.A., Eds.; Cambridge University Press: Cambridge, UK, 1968; pp. 94–100. [Google Scholar]
- Greuter, W.; Burdet, R.M.; Long, G. Med-Checklist 4; Conservatoire et Jardin botaniques de la Ville de Genève: Berlin, Germany; Gèneve, Switzerland, 1989. [Google Scholar]
- Talavera, S. Genista L. In Flora Iberica; Talavera, S., Aedo, C., Castroviejo, S., Romero Zarco, C., Sáez, L., Salgueiro, F.J., Velayos, M., Eds.; CSIC: Madrid, Spain, 1999; Volume 7, pp. 45–119. [Google Scholar]
- Bacchetta, G.; Brullo, S.; Cusma Velari, T.; Feoli Chiapella, L.; Kosovel, V. Taxonomic notes on the Genista ephedroides Group (Fabaceae) from the Mediterranean area. Novon J. Bot. Nomencl. 2011, 21, 4–19. [Google Scholar] [CrossRef]
- Greuter, W. Botanical diversity, endemism, rarity, and extinction in the Mediterranean area: An analysis based on the published volumes of Med-Checklist. Bot. Chron. 1991, 10, 63–79. [Google Scholar]
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: New York, NY, USA, 2005. [Google Scholar]
- Mittermeier, R.A.; Robles Gil, P.; Hoffman, M.; Pilgrim, J.; Brooks, T.; Mittermeier, C.G.; Lamoreux, J.; da Fonseca, G.A.B. Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; University of Chicago Press: Chicago, IL, USA, 2005. [Google Scholar]
- Cañadas, E.M.; Fenu, G.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within hotspots: Endemic plant richness, environmental drivers, and implications for conservation. Biol. Conserv. 2014, 170, 282–291. [Google Scholar] [CrossRef]
- Greuter, W. Diversity of Mediterranean Island Floras; Bocconea: Milano, Italy, 2001; Volume 13, pp. 55–64. [Google Scholar]
- Junikka, L.; Uotila, P.; Lahti, T. A phytogeographical comparison of the major Mediterranean islands on the basis of Atlas Florae Europaeae. Willdenowia 2006, 36, 379–388. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Biodiversity Hotspots in the Mediterranean Basin: Setting Global Conservation Priorities. Conserv. Biol. 2001, 13, 1510–1513. [Google Scholar] [CrossRef]
- Mansion, G.; Rosenbaum, G.; Schoenenberger, N.; Bacchetta, G.; Rosselló, J.A.; Conti, E. Phylogenetic Analysis Informed by Geological History Supports Multiple, Sequential Invasions of the Mediterranean Basin by the Angiosperm Family Araceae. Syst. Biol. 2008, 57, 269–285. [Google Scholar] [CrossRef]
- Gianguzzi, L.; Cusimano, D.; Bonventre, V.; Romano, S.; Ilardi, V. Bio-ecological, phytosociological and conservation aspects of relictual and disjointed populations of Simethis mattiazzi (Vandelli) Sacc. (Xanthorrhoeaceae) in the Channel of Sicily. Acta Bot. Gall. 2012, 159, 303–318. [Google Scholar] [CrossRef]
- Chen, C.; Qi, Z.; Xu, X.; Comes, H.P.; Koch, M.A.; Jin, X.; Fu, C.; Qiu, Y. Understanding the formation of Mediterranean–African–Asian disjunctions: Evidence for Miocene climate-driven vicariance and recent long-distance dispersal in the Tertiary relict Smilax aspera (Smilacaceae). New Phytol. 2014, 204, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Hardion, L.; Dumas, P.-J.; Abdel-Samad, F.; Kharrat, M.B.D.; Surina, B.; Affre, L.; Médail, F.; Bacchetta, G.; Baumel, A. Geographical isolation caused the diversification of the Mediterranean thorny cushion-like Astragalus L. sect. Tragacantha DC. (Fabaceae). Mol. Phylogenetics Evol. 2016, 97, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Affenzeller, M.; Kadereit, J.W.; Comes, H.P. Parallel bursts of recent and rapid radiation in the Mediterranean and Eritreo-Arabian biodiversity hotspots as revealed by Globularia and Campylanthus (Plantaginaceae). J. Biogeogr. 2018, 45, 552–566. [Google Scholar] [CrossRef]
- Bacchetta, G.; Brullo, S.; Chiapella, L.F.; Velari, T.C.; Fenu, G.; Del Galdo, G.G. Taxonomic remarks on Genista salzmannii group (Fabaceae) in Sardinia and Corsica. Phytotaxa 2020, 449, 31–51. [Google Scholar] [CrossRef]
- Fernández Prieto, J.A.; Sanna, M.; Bueno, Á.; Pérez, M. Genista anglica (Fabaceae): One very diverse species or one species complex? J. Plant Res. 2016, 129, 411–422. [Google Scholar] [CrossRef]
- De Castro, O.; Vallariello, R.; Del Guacchio, E. Integration of morphology, genetics, historical and ethnobotanical data: A case of an enigmatic Genista (Fabaceae) from Ischia Island (southern Italy). Phytotaxa 2013, 82, 64–68. [Google Scholar] [CrossRef]
- Bacchetta, G.; Brullo, S.; Caputo, P.; De Castro, O.; Del Guacchio, E.; Dettori, C.A.; Giusso del Galdo, G.; Grillo, O.; Piazza, C. Morphological and micro-morphological comparative study of Genista etnensis populations. Not. Soc. Bot. Ital. 2016, 0, 27–28. [Google Scholar]
- Ortolani, E.O.; Rafinesque-Schmaltz, C.S. Statistica Generale di Sicilia; Dalla Reale stamperia: Palermo, Italy, 1810. [Google Scholar]
- Inferrera, G. La Genista aetnensis nel messinese. Boll. Soc. Bot. Ital. 1904, 6, 270–272. [Google Scholar]
- La Mantia, T.; Rühl, J.; Massa, B.; Pipitone, S.; Verde, G.L.; Bueno, R.S. Vertebrate-mediated seed rain and artificial perches contribute to overcome seed dispersal limitation in a Mediterranean old field. Restor. Ecol. 2019, 27, 1393–1400. [Google Scholar] [CrossRef]
- Moris, J.H. Stirpium Sardoarum Elenchus; Ex Typis Regiis: Carali, Belgium, 1827; Volume 1. [Google Scholar]
- Arrigoni, P.V. Flora dellIsola di Sardegna, 3rd ed.; Carlo Delfino Editore: Sassari, Italy, 2010. [Google Scholar]
- Vivant, J. Sur quelques plantes de Corse. Monde des Plantes 1966, 351, 12–14. [Google Scholar]
- Paradis, G. Notes et Contributions à la Flore de Corse, IX. Candollea 1993, 48, 551–555. [Google Scholar]
- Piazza, C.; Paradis, G. Précisions sur les stations d’une espèce très rare en Corse: Genista aetnensis. «Etat des lieux» en 1995. Monde des Plantes 1996, 456, 9–12. [Google Scholar]
- Fridlender, A. Observations sur le Genêt de l’Etna en Corse: Genista etnensis (Biv.) DC. subsp. fraisseorum subsp. nova. Bull. Mens. Soc. Linn. Lyon. 2018, 87, 73–95. [Google Scholar]
- Agostini, R. Alcuni reperti interessanti la flora della Campania. Delpinoa 1959, 1, 42–68. [Google Scholar]
- Del Guacchio, E.; La Valva, V. The non-native vascular flora of Campania (southern Italy) [Appendices]. Plant Biosyst. 2017, 52, 767–779. [Google Scholar] [CrossRef]
- Stinca, A.; Conti, P.; Menegazzi, G.; Chirico, G.B.; Bonanomi, G. Invasion Impact of the Nitrogen-fixing Shrub Genista aetnensis on Vesuvius Grand Cone. Procedia Environ. Sci. 2013, 19, 865–874. [Google Scholar] [CrossRef][Green Version]
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Arrigoni, P.V.; Vannelli, S. La «Genista aetnensis» (Raf.) DC. in Sardegna. Webbia 1967, 22, 1–20. [Google Scholar] [CrossRef]
- Valsecchi, F. Il genere Genista L. in Italia: I. Le specie delle sezioni Erinacoides Spach, Ephedrospartum Spach, Aureospartum sect. nova. Webbia 1993, 48, 779–884. [Google Scholar]
- Spada, F.; Cutini, M.; Paura, B. Considerazioni fitostoriche sulla zonazione altitudinale della vegetazione di alcuni rilievi dell’Appennino meridionale e della Sicilia. Biogeographia 2011, 30, 95–112. [Google Scholar] [CrossRef]
- Vivant, J. Quelques notes à propos de plantes vasculaires de la Corse. Bull. Soc. Bot. Fr. 1974, 121, 27–36. [Google Scholar] [CrossRef][Green Version]
- Deschâtres, R. À propos du Genista etnensis (Biv.) DC. en Corse. Rev. Sci. Bourbon. Cent. Fr. 1980, 1979, 4–10. [Google Scholar]
- Verlaque, R.; Contandriopoulos, J.; Aboucaya, A. Cytotaxonomie et conservation de la flore insulaire: Les espèces endémiques ou rares de Corse. Ecol. Mediterr. 1995, 21, 257–268. [Google Scholar] [CrossRef]
- Jeanmonod, D.; Gamisans, J. Flora corsica, ed. 2. Bull. Soc. Bot. Cent. Ouest 2013, 39, 1–1074. [Google Scholar]
- Nery, L.; Delage, A. Notes à la Flore de Corse, XXV. Candollea 2015, 70, 109–140. [Google Scholar]
- Spach, E. Revisio generis Genista. Ann. Des Sci. Nat. Bot. Ser. 3. 1845, 3, 102–158. [Google Scholar]
- Presl, C.B. Botanische Bemerkungen. Abh. Königl. Böhm. Ges. Wiss. ser. 5. 1845, 3, 431–584. [Google Scholar]
- Cristofolini, G.; Feoli Chiapella, L. Serological systematics of the tribe Genisteae (Fabaceae). Taxon 1977, 26, 43–56. [Google Scholar] [CrossRef]
- De Castro, O.; Cozzolino, S.; Jury, S.L.; Caputo, P. Molecular relationships in Genista L. sect. Spartocarpus Spach (Fabaceae). Plant Syst. Evol. 2002, 231, 91–108. [Google Scholar] [CrossRef]
- Pardo, C.; Cubas, P.; Tahiri, H. Molecular phylogeny and systematics of Genista (Leguminosae) and related genera based on nucleotide sequences of nrDNA (ITS region) and cpDNA (trnL-trnF intergenic spacer). Plant Syst. Evol. 2004, 244, 93–119. [Google Scholar] [CrossRef]
- Bedini, G.; Peruzzi, L. Chrobase.it—Chromosome Numbers for the Italian Flora v. 2.0. Available online: http://bot.biologia.unipi.it/chrobase/ (accessed on 4 February 2022).
- Cusma Velari, T.; Feoli Chiapella, L.; Kosovel, V. A karyological study of Genista sect. Spartocarpus Spach (Cytiseae-Fabaceae). Webbia 2013, 66, 57–68. [Google Scholar] [CrossRef]
- Walpers, G.G. Annales Botanices Systematicae; Sumtibus F. Hofmeister: Lipsiae, Germany, 1848; Volume 1. [Google Scholar]
- Rizzi Longo, L.; Feoli Chiapella, L. Contribution to the systematics of Genista L. sect. Spartocarpus Spach (Genisteae, Fabaceae) with emphasis on palynological data. Studia Geobot. 1994, 14, 41–62. [Google Scholar]
- Cennamo, P.; Del Guacchio, E.; Jury, S.L.; Caputo, P. Molecular markers in Viola L. subsect. Viola: Application and taxonomic implications for the identification of dubious herbarium specimens. Plant Biosyst. 2011, 145, 306–323. [Google Scholar] [CrossRef]
- Cennamo, P.; Del Guacchio, E.; Paino, L.; De Castro, O.; Menale, B.; Vazquez-Torres, M.; Caputo, P. Genetic structure of Ipomoea imperati (Convolvulaceae) in the Mediterranean region and implications for its conservation. Phytotaxa 2013, 141, 40–54. [Google Scholar] [CrossRef][Green Version]
- De Castro, O.; Innangi, M.; Di Maio, A.; Menale, B.; Bacchetta, G.; Pires, M.; Noble, V.; Gestri, G.; Conti, F.; Peruzzi, L. Disentangling phylogenetic relationships in a hotspot of diversity: The butterworts (Pinguicula L., Lentibulariaceae) endemic to Italy. PLoS ONE 2016, 11, e0167610. [Google Scholar] [CrossRef] [PubMed]
- De Castro, O.; Geraci, A.; Mannino, A.M.; Mormile, N.; Santangelo, A.; Troia, A. A Contribution to the Characterization of Ruppia drepanensis (Ruppiaceae), a Key Species of Threatened Mediterranean Wetlands. Ann. Mo. Bot. Gard. 2021, 106, 1–9. [Google Scholar] [CrossRef]
- De Castro, O.; Innangi, M.; Menale, B. Message in a bottle: The Mediterranean Sea currents acted as protagonists in shaping the distribution of the sea daffodil (Pancratium maritimum, Amaryllidaceae). Bot. J. Linn. Soc. 2020, 194, 207–220. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef]
- Prince, L.M. Plastid Primers for Angiosperm Phylogenetics and Phylogeography. Appl. Plant Sci. 2015, 3, 1400085. [Google Scholar] [CrossRef]
- Sang, T.; Crawford, D.J.; Stuessy, T.F. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). Am. J. Bot. 1997, 84, 1120–1136. [Google Scholar] [CrossRef] [PubMed]
- Tate, J.A.; Simpson, B.B. Paraphyly of Tarasa (Malvaceae) and diverse origins of the polyploid species. Syst. Bot. 2003, 28, 723–737. [Google Scholar] [CrossRef]
- Aceto, S.; Caputo, P.; Cozzolino, S.; Gaudio, L.; Moretti, A. Phylogeny and Evolution of Orchis and Allied Genera Based on ITS DNA Variation: Morphological Gaps and Molecular Continuity. Mol. Phylogenetics Evol. 1999, 13, 67–76. [Google Scholar] [CrossRef] [PubMed]
- De Castro, O.; Di Maio, A.; García, J.A.L.; Piacenti, D.; Vázquez-Torres, M.; De Luca, P. Plastid DNA sequencing and nuclear SNP genotyping help resolve the puzzle of central American Platanus. Ann. Bot. 2013, 112, 589–602. [Google Scholar] [CrossRef] [PubMed][Green Version]
- JJarne, P.; Lagoda, P.J. Microsatellites, from molecules to populations and back. Trends Ecol. Evol. 1996, 11, 424–429. [Google Scholar] [CrossRef]
- Goldstein, D.G.; Pollock, D.D. Launching microsatellites: A review of mutation processes and methods of phylogenetic inference. J. Hered. 1997, 88, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Provan, J.; Powell, W.; Hollingsworth, P.M. Chloroplast microsatellites: New tools for studies in plant ecology and evolution. Trends Ecol. Evol. 2001, 16, 142–147. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed]
- Akaike, H. A new look at the statistical model identification. System identification and time-series analysis. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Fiz-Palacios, O.; Valcárcel, V. From Messinian crisis to Mediterranean climate: A temporal gap of diversification recovered from multiple plant phylogenies. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 130–137. [Google Scholar] [CrossRef]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Fenu, G.; Fois, M.; Cañadas, E.M.; Bacchetta, G. Using endemic-plant distribution, geology and geomorphology in biogeography: The case of Sardinia (Mediterranean Basin). Syst. Biodiver. 2014, 12, 181–193. [Google Scholar] [CrossRef]
- Fois, M.; Farris, E.; Calvia, G.; Campus, G.; Fenu, G.; Porceddu, M.; Bacchetta, G. The Endemic Vascular Flora of Sardinia: A Dynamic Checklist with an Overview of Biogeography and Conservation Status. Plants 2022, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Poczai, P.; Hyvönen, J. Nuclear ribosomal spacer regions in plant phylogenetics: Problems and prospects. Mol. Biol. Rep. 2009, 37, 1897–1912. [Google Scholar] [CrossRef] [PubMed]
- Suc, J.P. Origin and evolution of the Mediterranean vegetation and climate in Europe. Nature 1984, 307, 429–432. [Google Scholar] [CrossRef]
- Tzedakis, P.C. Cenozoic climate and vegetation changes in the Mediterranean. In The Physical Geography of the Mediterranean; Woodward, J., Ed.; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Head, M.J.; Gibbard, P.L. Early-Middle Pleistocene transitions: An overview and recommendation for the defining boundary. In Early-Middle Pleistocene Transitions: The Land-Ocean Evidence; Head, M.J., Gibbard, P.L., Eds.; Geological Society, Special Publications; The Geological Society of London: London, UK, 2005; Volume 247, pp. 1–18. [Google Scholar]
- Magri, D.; Di Rita, F.; Aranbarri, J.; Fletcher, W.; González-Sampériz, P. Quaternary disappearance of tree taxa from Southern Europe: Timing and trends. Quat. Sci. Rev. 2017, 163, 23–55. [Google Scholar] [CrossRef]
- Magri, D.; Palombo, M.R. Early to Middle Pleistocene dynamics of plant and mammal communities in South West Europe. Quat. Int. 2013, 288, 63–72. [Google Scholar] [CrossRef]
- Arrigoni, P.V. Flora dell’Isola di Sardegna; Carlo Delfino Editore: Sassari, Italy, 2006; Volume 1. [Google Scholar]
- Boccaletti, M.; Ciaranfi, N.; Cosentino, D.; Deiana, G.; Gelati, R.; Lentini, F.; Massari, F.; Moratti, G.; Pescatore, T.; Lucchi, F.R.; et al. Palinspastic restoration and paleogeographic reconstruction of the peri-Tyrrhenian area during the Neogene. Palaeogeogr. Palaeoclim. Palaeoecol. 1990, 77, 1–41. [Google Scholar] [CrossRef]
- Rook, L.; Gallai, G.; Torre, D. Lands and endemic mammals in the Late Miocene of Italy: Constrains for paleogeographic outlines of Tyrrhenian area. Palaeogeogr. Palaeoclim. Palaeoecol. 2006, 238, 263–269. [Google Scholar] [CrossRef]
- Advokaat, E.L.; van Hinsbergen, D.J.; Maffione, M.; Langereis, C.G.; Vissers, R.L.; Cherchi, A.; Schroeder, R.; Madani, H.; Columbu, S. Eocene rotation of Sardinia, and the paleogeography of the western Mediterranean region. Earth Planet. Sci. Lett. 2014, 401, 183–195. [Google Scholar] [CrossRef]
- Schmitt, T.; Fritz, U.; Delfino, M.; Ulrich, W.; Habel, J.C. Biogeography of Italy revisited: Genetic lineages confirm major phylogeographic patterns and a pre-Pleistocene origin of its biota. Front. Zool. 2021, 18, 34. [Google Scholar] [CrossRef] [PubMed]
- Paiero, P.; Martini, F.; Colpi, C. Leguminose Arboree E Arbustive in Italia; Edizioni Lint: Trieste, Italy, 1993. [Google Scholar]
- Pemberton, R.W.; Irving, D.W. Elaiosomes on Weed Seeds and the Potential for Myrmecochory in Naturalized Plants. Weed Sci. 1990, 38, 615–619. [Google Scholar] [CrossRef]
- Devenish, A.J.M.; Gomez, C.; Bridle, J.R.; Newton, R.J.; Sumner, S. Invasive ants take and squander native seeds: Implications for native plant communities. Biol. Invasions 2018, 21, 451–466. [Google Scholar] [CrossRef]
- Van Dommelen, P. Colonial interactions and hybrid practices: Phoenician and Carthaginian Settlement in the Ancient Mediterranean. In The Archaeology of Colonial; Encounters Stein, G.J., Ed.; Comparative Perspectives: Oxford, UK, 2005; pp. 109–142. [Google Scholar]
- De Marco, G.; Altieri, A.; Estabrook, G.F. Relazioni evolutive e biogeografiche dei popolamenti ad areale disgiunto di Genista ephedroides DC. Biogeographia 1987, 11, 115–130. [Google Scholar] [CrossRef]
- De Castro, O.; Véla, E.; Vendramin, G.G.; Gargiulo, R.; Caputo, P. Genetic structure in the Genista ephedroides complex (Fabaceae) and implications for its present distribution. Bot. J. Linn. Soc. 2015, 177, 607–618. [Google Scholar] [CrossRef]
- Coulot, P.; Rabaute, P. Monographie des Leguminosae de France: Tribus des Fabeae, des Cicereae et des Genisteae. Bull. Soc. Bot. Centre-Ouest 2016, 46, 1–902. [Google Scholar]
- Paradis, G.; Chiappe, M. Origine du Genista de l’île Mezzu Mare (Corse): Une énigme résolue. Bull. Soc. Bot. Centre-Ouest 2017, 48, 143–146. [Google Scholar]
- Paradis, G.; Appietto, A.; Piazza, C. Répartition en 2018 sur l’île Mezzu Mare (Corse) du genêt introduit Genista tyrrhena subsp. pontiana. Monde ses Plantes 2015, 517, 3–5. [Google Scholar]
- Kress, W.J.; Erickson, D.L. A Two-Locus Global DNA Barcode for Land Plants: The Coding rbcL Gene Complements the Non-Coding trnH-psbA Spacer Region. PLoS ONE 2007, 2, e508. [Google Scholar] [CrossRef] [PubMed]
- Calviño, C.I.; Downie, S.R. Circumscription and phylogeny of Apiaceae subfamily Saniculoideae based on chloroplast DNA sequences. Mol. Phylogenet. Evol. 2007, 44, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Taberlet, P.; Gielly, L.; Pautou, G.; Bouvet, J. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Mol. Biol. 1991, 17, 1105–1109. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.B. Four primer pairs for the amplification of chloroplast intergenic regions with intraspecific variation. Mol. Ecol. 1999, 8, 521–523. [Google Scholar] [PubMed]



| Code | Taxon | Locality | N | |
|---|---|---|---|---|
| B | G. etnensis (Raf.) DC. | Sicily | Mount Baracca | 10 |
| F | G. etnensis | Sicily | Fornazzo | 10 |
| G | G. etnensis | Sardinia | Genna Silana | 10 |
| L | G. etnensis | Sardinia | Mount Limbara | 10 |
| N | G. etnensis | Sardinia | Villagrande Strisaili | 10 |
| Pf | G. etnensis | Corsica | Serra Fiumorbu | 10 |
| S + Ss | G. etnensis | Corsica | Solenzara + road between Sari and Solenzara | 6 + 4 |
| T | G. etnensis | Corsica | Solaro | 3 |
| V | G. etnensis | Southern Italy | Mount Vesuvius | 10 |
| GF | G. fasselata Decne. | Israel | Mount Carmel | 5 |
| Pr1 * | GenBank | Pr8 * | GenBank | Pr11 * | GenBank | Pr18 * | GenBank | Pr38 * | GenBank | H | Pop(n) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 213 | 214 | 320 | 11 | 332 | 149 | 349 | 112 | 188 | 189 | 190 | |||||||
| T | G | --- | OP830512 | --- | OP830517 | C | OP830520 | A | T | OP830523 | AATAAGA | A | G | A | OP830526 | P1 | B(9), F(5) |
| T | G | --- | --- | C | C | G | OP830524 | --- | A | G | A | OP830527 | P2 | B(1), F(5), V(5) | |||
| T | G | AATTATACATATATATAA | OP830513 | --- | T | OP830521 | A | T | --- | T | C | T | OP830528 | P3 | G(8), N(10), Ss(1) | ||
| T | G | --- | --- | C | A | T | --- | T | C | T | P4 | G(2), L(10) | |||||
| G | G | --- | OP830514 | --- | C | A | T | --- | T | C | T | P5 | P(10), S(6), Ss(3), T(3) | ||||
| T | G | --- | TATAC | OP830518 | C | C | G | --- | A | G | A | P6 | V(4) | ||||
| T | T | --- | OP830515 | --- | C | A | T | --- | A | G | A | P7 | V(1) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Castro, O.; Bacchetta, G.; Brullo, S.; Del Guacchio, E.; Di Iorio, E.; Piazza, C.; Caputo, P. Variability and Nativeness in the Mediterranean Taxa: Divergence and Phylogeography of Genista etnensis (Fabaceae) Inferred from Nuclear and Plastid Data. Plants 2022, 11, 3171. https://doi.org/10.3390/plants11223171
De Castro O, Bacchetta G, Brullo S, Del Guacchio E, Di Iorio E, Piazza C, Caputo P. Variability and Nativeness in the Mediterranean Taxa: Divergence and Phylogeography of Genista etnensis (Fabaceae) Inferred from Nuclear and Plastid Data. Plants. 2022; 11(22):3171. https://doi.org/10.3390/plants11223171
Chicago/Turabian StyleDe Castro, Olga, Gianluigi Bacchetta, Salvatore Brullo, Emanuele Del Guacchio, Emanuela Di Iorio, Carole Piazza, and Paolo Caputo. 2022. "Variability and Nativeness in the Mediterranean Taxa: Divergence and Phylogeography of Genista etnensis (Fabaceae) Inferred from Nuclear and Plastid Data" Plants 11, no. 22: 3171. https://doi.org/10.3390/plants11223171
APA StyleDe Castro, O., Bacchetta, G., Brullo, S., Del Guacchio, E., Di Iorio, E., Piazza, C., & Caputo, P. (2022). Variability and Nativeness in the Mediterranean Taxa: Divergence and Phylogeography of Genista etnensis (Fabaceae) Inferred from Nuclear and Plastid Data. Plants, 11(22), 3171. https://doi.org/10.3390/plants11223171