
How Does Quinoa (Chenopodium quinoa Willd.) Respond to Phosphorus Fertilization and Irrigation Water Salinity?

 ,
, 

Abstract
:1. Introduction
2. Results
2.1. Analysis of Variance of Growth Parameters and Leaf Nutrient Concentration
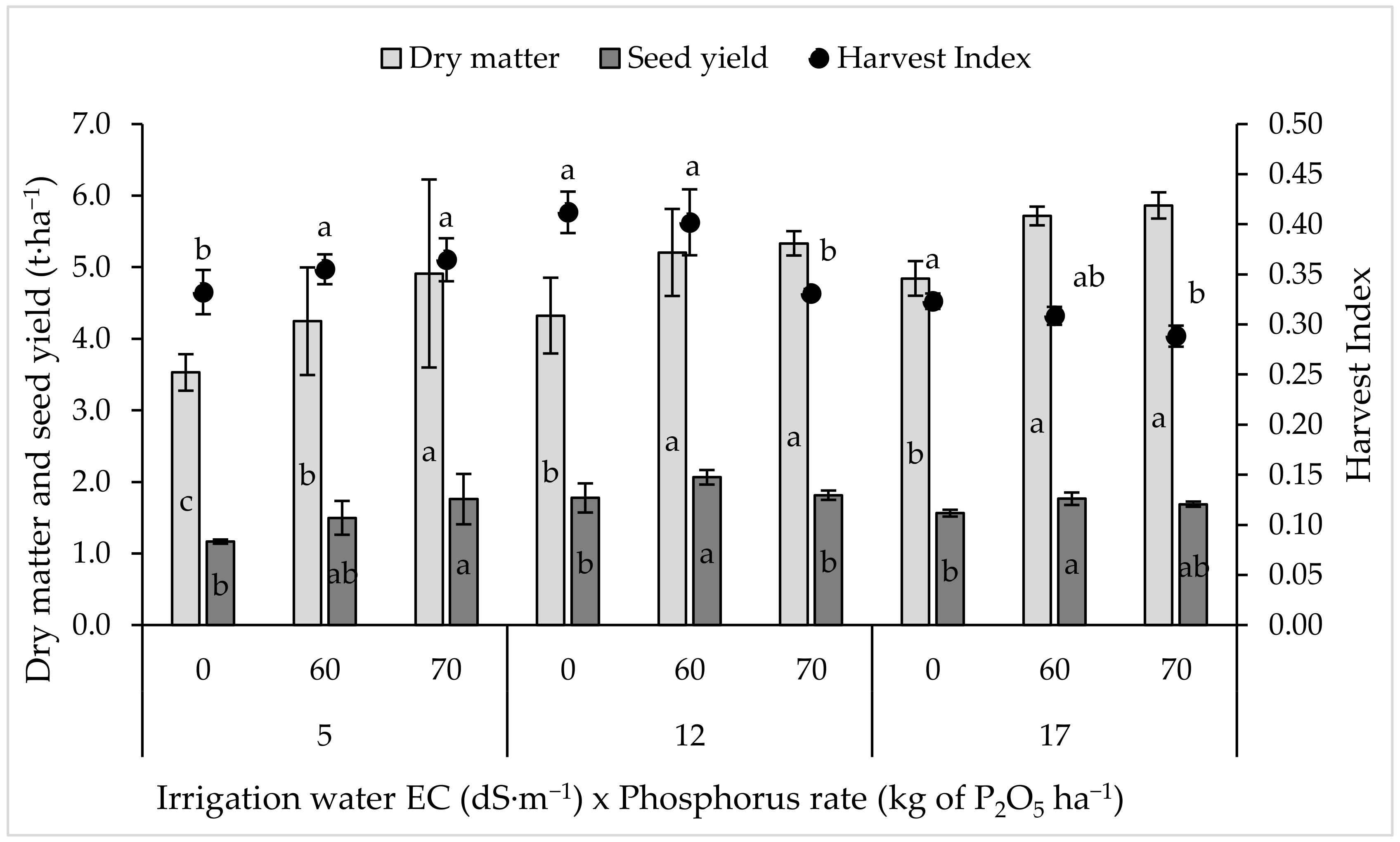
2.2. Seed and Biomass Yield and Harvest Index
2.3. Crop and Irrigation Water Productivity
2.4. Leaf Mineral Content
2.5. Correlation Matrix
2.6. Principal Component Analysis (PCA)
3. Discussion
3.1. Effect of Salinity Level on Biomass and Seed Yield of Quinoa
3.2. Effect of Phosphorus Rate on Biomass and Seed Yield
3.3. Leaf Mineral Content
4. Materials and Methods
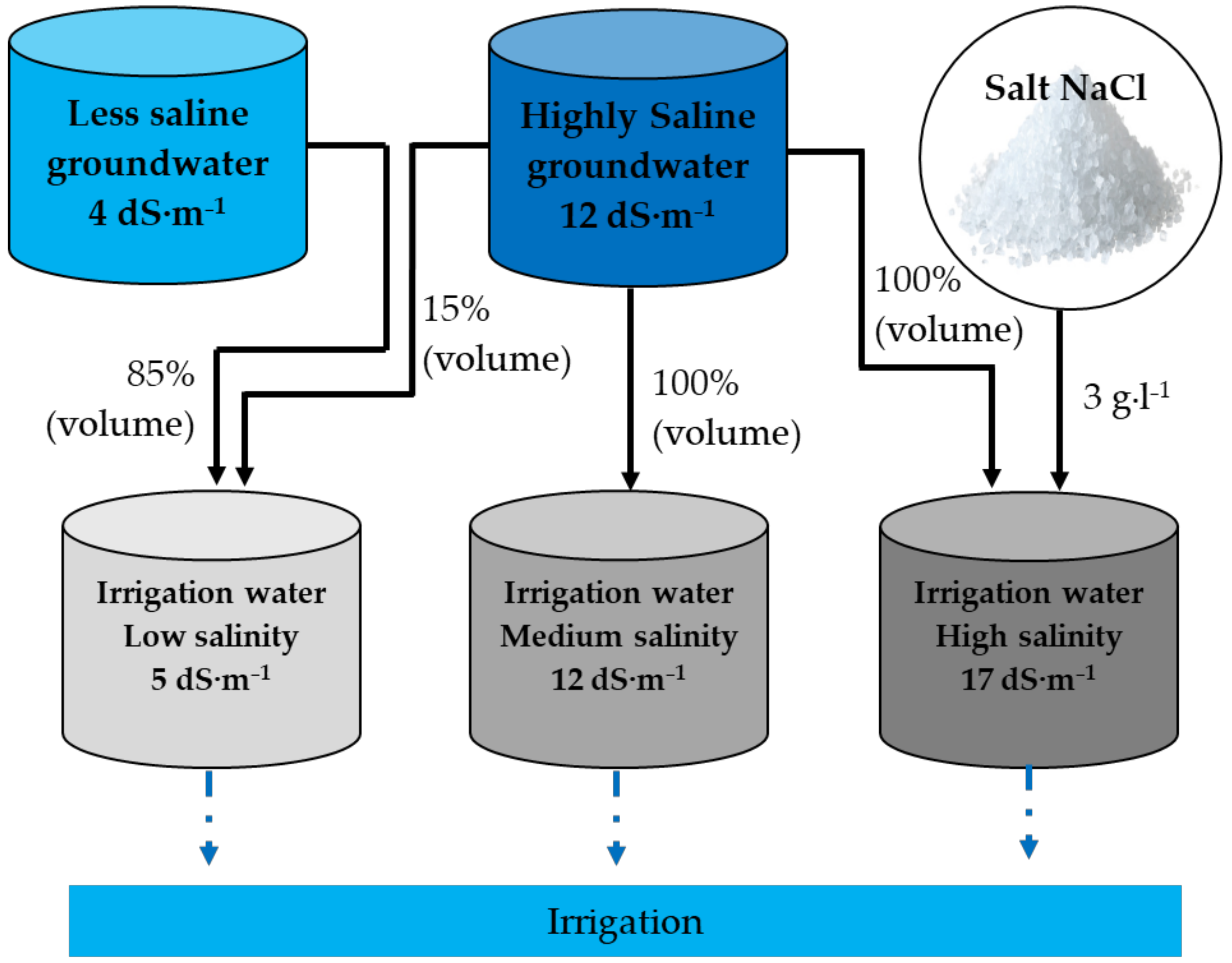
4.1. Experimental Site
4.2. Experimental Detail and Crop Management
4.3. Observations
4.3.1. Seed and Dry Matter Yield
4.3.2. Crop and Irrigation Water Productivity
4.3.3. Chemical Analysis
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shahid, S.A.; Zaman, M.; Heng, L. Soil Salinity: Historical Perspectives and a World Overview of the Problem. In Guideline for Salinity Assessment, Mitigation and Adaptation Using Nuclear and Related Techniques; Springer: Berlin/Heidelberg, Germany, 2018; pp. 43–53. [Google Scholar]
- Fao: Food and Agriculture Organization of the United Nations. Global Map of Salt-Affected Soils. 2021. Available online: https://www.fao.org/soils-portal/data-hub/soil-maps-and-databases/global-map-of-salt-affected-soils/en/ (accessed on 3 January 2022).
- Jaramillo Roman, V. Salt Tolerance Strategies of the Ancient Andean Crop Quinoa; Wageningen University: Wageningen, The Netherlands, 2021. [Google Scholar]
- Hopmans, J.W.; Qureshi, A.S.; Kisekka, I.; Munns, R.; Grattan, S.R.; Rengasamy, P.; Ben-Gal, A.; Assouline, S.; Javaux, M.; Minhas, P.S.; et al. Critical Knowledge Gaps and Research Priorities in Global Soil Salinity. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2021; Volume 169, pp. 1–191. ISBN 978-0-12-824590-3. [Google Scholar]
- Munns, R.; Day, D.A.; Fricke, W.; Watt, M.; Arsova, B.; Barkla, B.J.; Bose, J.; Byrt, C.S.; Chen, Z.-H.; Foster, K.J.; et al. Energy Costs of Salt Tolerance in Crop Plants. New Phytol. 2020, 225, 1072–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowers, T.J.; Colmer, T.D. Salinity Tolerance in Halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Choukr-Allah, R.; Rao, N.K.; Hirich, A.; Shahid, M.; Alshankiti, A.; Toderich, K.; Gill, S.; Butt, K.U.R. Quinoa for Marginal Environments: Toward Future Food and Nutritional Security in MENA and Central Asia Regions. Front. Plant Sci. 2016, 7, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezzouk, F.Z.; Shahid, M.A.; Elouafi, I.A.; Zhou, B.; Araus, J.L.; Serret, M.D. Agronomic Performance of Irrigated Quinoa in Desert Areas: Comparing Different Approaches for Early Assessment of Salinity Stress. Agric. Water Manag. 2020, 240, 106205. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Biondi, S.; Martínez, E.A.; Orsini, F.; Antognoni, F.; Jacobsen, S.-E. Quinoa—A Model Crop for Understanding Salt-Tolerance Mechanisms in Halophytes. Plant Biosyst.—Int. J. Deal. Asp. Plant Biol. 2016, 150, 357–371. [Google Scholar] [CrossRef]
- Huarancca Reyes, T.; Scartazza, A.; Castagna, A.; Cosio, E.G.; Ranieri, A.; Guglielminetti, L. Physiological Effects of Short Acute UVB Treatments in Chenopodium Quinoa Willd. Sci. Rep. 2018, 8, 371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, S.-E.; Monteros, C.; Christiansen, J.L.; Bravo, L.A.; Corcuera, L.J.; Mujica, A. Plant Responses of Quinoa (Chenopodium Quinoa Willd.) to Frost at Various Phenological Stages. Eur. J. Agron. 2005, 2, 131–139. [Google Scholar] [CrossRef]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa Abiotic Stress Responses: A Review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Hirich, A.; Choukr-Allah, R.; Jacobsen, S.-E. Deficit Irrigation and Organic Compost Improve Growth and Yield of Quinoa and Pea. J. Agron. Crop. Sci. 2014, 200, 390–398. [Google Scholar] [CrossRef]
- Razzaghi, F.; Jacobsen, S.-E.; Jensen, C.; Andersen, M. Ionic and Photosynthetic Homeostasis in Quinoa Challenged by Salinity and Drought-Mechanisms of Tolerance. Funct. Plant Biol. 2014, 42, 136–148. [Google Scholar] [CrossRef]
- Jaramillo Roman, V.; den Toom, L.A.; Castro Gamiz, C.; van der Pijl, N.; Visser, R.G.F.; van Loo, E.N.; van der Linden, C.G. Differential Responses to Salt Stress in Ion Dynamics, Growth and Seed Yield of European Quinoa Varieties. Environ. Exp. Bot. 2020, 177, 104146. [Google Scholar] [CrossRef]
- Adolf, V.I.; Jacobsen, S.-E.; Shabala, S. Salt Tolerance Mechanisms in Quinoa (Chenopodium Quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Hirich, A.; Jelloul, A.; Choukr-Allah, R.; Jacobsen, S.-E. Saline Water Irrigation of Quinoa and Chickpea: Seedling Rate, Stomatal Conductance and Yield Responses. J. Agron. Crop. Sci. 2014, 200, 378–389. [Google Scholar] [CrossRef]
- Bose, J.; Munns, R.; Shabala, S.; Gilliham, M.; Pogson, B.; Tyerman, S.D. Chloroplast Function and Ion Regulation in Plants Growing on Saline Soils: Lessons from Halophytes. J. Exp. Bot. 2017, 68, 3129–3143. [Google Scholar] [CrossRef] [PubMed]
- Tyerman, S.D.; Munns, R.; Fricke, W.; Arsova, B.; Barkla, B.J.; Bose, J.; Bramley, H.; Byrt, C.; Chen, Z.; Colmer, T.D.; et al. Energy Costs of Salinity Tolerance in Crop Plants. New Phytol. 2019, 221, 25–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grattan, S.R.; Grieve, C.M. Mineral Element Acquisition and Growth Response of Plants Grown in Saline Environments. Agric. Ecosyst. Environ. 1992, 38, 275–300. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops—What Is the Cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Maleki, P.; Bahrami, H.A.; Saadat, S.; Sharifi, F.; Dehghany, F.; Salehi, M. Salinity Threshold Value of Quinoa (Chenopodium Quinoa Willd.) at various Growth Stages and the Appropriate irrigation Method by Saline water. Commun. Soil. Sci. Plant Anal. 2018, 49, 1815–1825. [Google Scholar] [CrossRef]
- Orsini, F.; Accorsi, M.; Gianquinto, G.; Dinelli, G.; Antognoni, F.; Carrasco, K.B.R.; Martinez, E.A.; Alnayef, M.; Marotti, I.; Bosi, S.; et al. Beyond the Ionic and Osmotic Response to Salinity in Chenopodium Quinoa: Functional Elements of Successful Halophytism. Funct. Plant Biol. 2011, 38, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Colla, G.; Rouphael, Y.; Cardarelli, M.; Tullio, M.; Rivera, C.M.; Rea, E. Alleviation of Salt Stress by Arbuscular Mycorrhizal in Zucchini Plants Grown at Low and High Phosphorus Concentration. Biol. Fertil. Soils 2008, 44, 501–509. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus Acquisition and Use: Critical Adaptations by Plants for Securing a Nonrenewable Resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schroder, J.J.; Cordell, D.; Smit, A.L.; Rosemarin, A. Sustainable Use of Phosphorus: EU Tender ENV.B1/ETU/2009/0025; Plant Research International: Wageningen, The Netherlands, 2010. [Google Scholar]
- Baker, A.; Ceasar, S.A.; Palmer, A.J.; Paterson, J.B.; Qi, W.; Muench, S.P.; Baldwin, S.A. Replace, Reuse, Recycle: Improving the Sustainable Use of Phosphorus by Plants. J. Exp. Bot. 2015, 66, 3523–3540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerda, A.; Bingham, F.T.; Hoffman, G.J. Interactive Effect of Salinity and Phosphorus on Sesame. Soil Sci. Soc. Am. J. 1977, 41, 915–918. [Google Scholar] [CrossRef]
- Shibli, R.A.; Sawwan, J.; Swaidat, I.; Tahat, M. Increased Phosphorus Mitigates the Adverse Effects of Salinity in Tissue Culture. Commun. Soil Sci. Plant Anal. 2001, 32, 429–440. [Google Scholar] [CrossRef]
- Wagdi, E.M.; Metwally, S.M.; Matar, M.K.; Yousef, N.N. Effect of Phosphorus in Alleviation Adverse Impacts of Salinity on Wheat Grown on Different Soils. Commun. Soil Sci. Plant Anal. 2013, 44, 1921–1936. [Google Scholar] [CrossRef]
- Bouras, H.; Bouaziz, A.; Bouazzama, B.; Hirich, A.; Choukr-Allah, R. How Phosphorus Fertilization Alleviates the Effect of Salinity on Sugar Beet (Beta Vulgaris L.) Productivity and Quality. Agronomy 2021, 11, 1491. [Google Scholar] [CrossRef]
- Alzubaidi, A.; Aljanabi, A.S.; Al-Rawi, A. Interaction between Nitrogen and Phosphorus Fertilizers and Soil Salinity and Its Effect on Growth and Ionic Composition of Corn (Zea Mays L.). In Genetic Aspects of Plant Mineral Nutrition; El Bassam, N., Dambroth, M., Loughman, B.C., Eds.; Springer: Dordrecht, The Netherlands, 1990; pp. 195–202. ISBN 978-94-010-7421-6. [Google Scholar]
- Awad, A.S.; Edwards, D.G.; Campbell, L.C. Phosphorus Enhancement of Salt Tolerance of Tomato. Crop. Sci. 1990, 30, 123–128. [Google Scholar] [CrossRef]
- Malik, R.S.; Gupta, A.P.; Haneklaus, S.; El Bassam, N. Role of Phosphorus (P-32) in Inducing Salt Tolerance in Sunflower. Landbauforsch. Volkenrode 1999, 49, 169–176. [Google Scholar]
- Plaut, Z.; Grieve, C.M. Photosynthesis of Salt-Stressed Maize as Influenced by Ca: Na Ratios in the Nutrient Solution. Plant Soil. 1988, 105, 283–286. [Google Scholar] [CrossRef]
- Ghazi, N.; Al-Karaki, R. Barley Response to Salt Stress at Varied Levels of Phosphorus. J. Plant Nutr. 1997, 20, 1635–1643. [Google Scholar]
- Brakez, M.; Daoud, S.; Harrouni, M.C.; Tachbibi, N.; Brakez, Z. 3-Nutritional Value of Chenopodium Quinoa Seeds Obtained from an Open Field Culture Under Saline Conditions. In Halophytes for Food Security in Dry Lands; Khan, M.A., Ozturk, M., Gul, B., Ahmed, M.Z., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 37–47. ISBN 978-0-12-801854-5. [Google Scholar]
- Long, N.V. Effects of Salinity Stress on Growth and Yield of Quinoa. Vietnam. J. Agric. Sci. 2016, 14, 321–327. [Google Scholar]
- Talebnejad, R.; Sepaskhah, A.R. Effect of Different Saline Groundwater Depths and Irrigation Water Salinities on Yield and Water Use of Quinoa in Lysimeter. Agric. Water Manag. 2015, 148, 177–188. [Google Scholar] [CrossRef]
- Khosh Kholgh Sima, N.A.; Ahmad, S.T.; Alitabar, R.A.; Mottaghi, A.; Pessarakli, M. Interactive Effects of Salinity and Phosphorus Nutrition on Physiological Responses of Two Barley Species. J. Plant Nutr. 2012, 35, 1411–1428. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Dikilitas, M.; Tuna, A. Alleviation of Salt Stress-Induced Adverse Effects on Maize Plants by Exogenous Application of Indoleacetic Acid (IAA) and Inorganic Nutrients—A Field Trial. Aust. J. Crop. Sci. 2013, 7, 249–254. [Google Scholar]
- Bouras, H.; Bouaziz, A.; Choukr-Allah, R.; Hirich, A.; Devkota, K.P.; Bouazzama, B. Phosphorus Fertilization Enhances Productivity of Forage Corn (Zea Mays L.) Irrigated with Saline Water. Plants 2021, 10, 2608. [Google Scholar] [CrossRef]
- Jahan, M.N.; Barua, S.; Ali, H.; Ali, M.N.; Chowdhury, M.S.H.; Hasan, M.M.; Ferdous, T.; Eti, F.S.; Khayer, A.; Hossen, K. Effects of Phosphorus Fertilization on Hybrid Varieties of Mungbean [Vigna Radiata (L.) Wilczek] in a Salinity Prone Area of the Subtropics. Acta Agrobot. 2020, 73. [Google Scholar] [CrossRef]
- Gulmezoglu, N. The Interactive Effects of Phosphorus and Salt on Growth, Water Potenital and Phosphorus Uptake in Green Beans. Appl. Ecol. Environ. Res. 2017, 15, 1831–1842. [Google Scholar] [CrossRef]
- Sadji-Ait Kaci, H. Interactive Effects of Salinity and Two Phosphorus Fertilizers on Growth and Grain Yield of Cicer Arietinum L. Section B Soil and Plant Science. Acta Agric. Scand. 2017, 67, 208–216. [Google Scholar] [CrossRef]
- Abd El-Hamed, E.; Metwally, S.; Matar, M.; Yousef, N. Impact of Phosphorus Fertilization in Alleviating the Adverse Effects of Salinity on Wheat Grown on Different Soil Types. Acta Agron. Hung. 2012, 60, 265–281. [Google Scholar] [CrossRef]
- Malhotra, H.; Vandana; Sharma, S.; Pandey, R. Phosphorus Nutrition: Plant Growth in Response to Deficiency and Excess. In Plant Nutrients and Abiotic Stress Tolerance; Springer: Singapore, 2018; pp. 171–190. ISBN 978-981-10-9043-1. [Google Scholar]
- Chapman, H.D.; Pratt, P.F. Methods of Analysis for Soils, Plants and Waters; University of California: Berkeley, CA, USA, 1961; p. 309. [Google Scholar]
- Olsen, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; USA Department of Agriculture: Washington, DC, USA, 1954.
- Ryan, J.; Estefan, G.; Rashid, A. Soil and Plant Analysis Laboratory Manual; ICARDA: Beirut, Lebanon, 2001. [Google Scholar]
- Aboukila, E.; Abdelaty, E. Assessment of Saturated Soil Paste Salinity from 1:2.5 and 1:5 Soil-Water Extracts for Coarse Textured Soils. Alex. Sci. Exch. J. 2017, 38, 722–732. [Google Scholar] [CrossRef]
- Khorsandi, F.; Yazdi, F.A. Estimation of Saturated Paste Extracts’ Electrical Conductivity from 1:5 Soil/Water Suspension and Gypsum. Commun. Soil Sci. Plant Anal. 2011, 42, 315–321. [Google Scholar] [CrossRef]
- He, Y.; DeSutter, T.; Prunty, L.; Hopkins, D.; Jia, X.; Wysocki, D.A. Evaluation of 1:5 Soil to Water Extract Electrical Conductivity Methods. Geoderma 2012, 185–186, 12–17. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | DF | DM | SY | HI | CWP | IWP | Leaf Nutrient Content | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OC | N | P | K | Mg | Ca | Zn | Fe | Na | |||||||
| Irrigation water salinity (S) | 2 | 0.018 * | 0.014 * | 0.00 *** | 0.005 ** | 0.013 * | 0.047 * | 0.023 * | 0.032 * | 0.021 | 0.044 * | 0.047 * | 0.54 | 0.26 | 0.010 ** |
| Phosphorus rate (P) | 2 | 0.027 * | 0.032 * | 0.047 * | 0.033 * | 0.02 * | 0.045 * | 0.43 | 0.021 * | 0.036 * | 0.029 * | 0.23 | 0.18 | 0.31 | 0.024 * |
| Interaction (S × P) | 4 | 0.90 ns | 0.14 ns | 0.018 * | 0.148 ns | 0.975 ns | 0.029 * | 0.37 | 0.039 * | 0.043 * | 0.170 ns | 0.049 * | 0.270 ns | 0.63 ns | 0.025 * |
| Irrigation Water EC | P Rate (kg of P2O5·ha−1) | OC (%) | N (%) | P (%) | K (%) | Mg (%) | Ca (%) | Zn (ppm) | Fe (ppm) | Na (%) | K/Na | Ca/Na | Mg/Na |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5 dS·m−1 | 0 | 40.4 ± 8.4 a | 1.4 ± 0.1 a | 0.14 ± 0.03 a | 5.3 ± 1 b | 2.3 ± 0.2 a | 3.1 ± 1.1 a | 22.6 ± 3 b | 343 ± 29 b | 3.3 ± 0.6 b | 1.6 ± 0.2 a | 1.1 ± 0.6 a | 0.7 ± 0.2 a |
| 60 | 40.0 ± 6.3 a | 1.2 ± 0.1 a | 0.12 ± 0.03 b | 6.4 ± 2 a | 2.2 ± 0.3 a | 2.6 ± 1 b | 26.1 ± 6 a | 324 ± 17 b | 3.5 ± 0.2 a | 1.8 ± 0.4 a | 0.8 ± 0.4 b | 0.6 ± 0.1 b | |
| 70 | 36.2 ± 2.3 a | 1.4 ± 0.1 a | 0.15 ± 0.02 a | 5.7 ± 1 ab | 2.2 ± 0.1 a | 2.8 ± 0.6 ab | 22.1 ± 3 b | 433 ± 16 a | 3.6 ± 0.3 a | 1.6 ± 0.6 a | 0.8 ± 0.1 b | 0.6 ± 0 b | |
| Average | 38.9 ± 5.7 A | 1.3 ± 0.1 B | 0.14 ± 0.02 B | 6 ± 1.5 B | 2.2 ± 0.2 A | 3 ± 0.9 A | 24 ± 4 A | 366 ± 69 A | 3 ± 0.4 B | 2 ± 0.4 A | 0.9 ± 0.4 A | 0.7 ± 0.1 A | |
| 12 dS·m−1 | 0 | 35.1 ± 1.8 a | 1.5 ± 0.1 a | 0.30 ± 0.13 b | 7.6 ± 1 b | 2 ± 0.1 b | 1.7 ± 0.1 b | 28.8 ± 7 ab | 280 ± 13 a | 4.6 ± 1 a | 1.7 ± 0.3 b | 0.4 ± 0.1 c | 0.5 ± 0.1 b |
| 60 | 34.3 ± 0.7 b | 1.7 ± 0.2 a | 0.34 ± 0.07 b | 8.6 ± 1 a | 2 ± 0.3 b | 1.9 ± 0.3 ab | 24.1 ± 5 b | 251 ± 16 a | 4.3 ± 0.3 ab | 2 ± 0.4 a | 0.4 ± 0 b | 0.5 ± 0 b | |
| 70 | 35.2 ± 1.7 a | 1.6 ± 0.1 a | 0.41 ± 0.24 a | 8.1 ± 0 ab | 2.4 ± 0.2 a | 2.1 ± 0.2 a | 33.3 ± 17 a | 212 ± 6 a | 4 ± 0.1 b | 2 ± 0.1 a | 0.5 ± 0.1 a | 0.6 ± 0.1 a | |
| Average | 34.8 ± 1.4 AB | 1.6 ± 0.1 A | 0.35 ± 0.15 A | 8 ± 0.7 A | 2.1 ± 0.2 AB | 2 ± 0.2 B | 29 ± 10 A | 248 ± 57 A | 4 ± 0.5 A | 2 ± 0.3 A | 0.5 ± 0.1 B | 0.5 ± 0.1 B | |
| 17 dS·m−1 | 0 | 34.8 ± 1.1 a | 1.6 ± 0.1 a | 0.25 ± 0.03 a | 8.2 ± 1 c | 1.9 ± 0.1 b | 1.6 ± 0.1 b | 15.3 ± 2 b | 248 ± 15 a | 4.5 ± 0.3 b | 1.8 ± 0.1 ab | 0.4 ± 0 b | 0.4 ± 0 ab |
| 60 | 34.3 ± 1.3 a | 1.6 ± 0 a | 0.26 ± 0.06 a | 9.3 ± 0 a | 2.1 ± 0.2 a | 1.7 ± 0.2 a | 27.6 ± 3 a | 244 ± 15 a | 4.5 ± 0.3 b | 2.1 ± 0.2 a | 0.4 ± 0 a | 0.5 ± 0 a | |
| 70 | 31.7 ± 1.7 b | 1.5 ± 0.1 a | 0.23 ± 0.04 b | 9.1 ± 0 b | 1.9 ± 0.2 b | 1.6 ± 0.2 b | 22.7 ± 8 ab | 214 ± 12 a | 5.5 ± 0.6 a | 1.7 ± 0.2 b | 0.3 ± 0 b | 0.3 ± 0 b | |
| Average | 33.6 ± 1.8 B | 1.6 ± 0.1 A | 0.25 ± 0.04 AB | 9 ± 0.3 A | 1.9 ± 0.2 A | 2 ± 0.1 A | 22 ± 3 A | 235 ± 34 A | 5 ± 0.4 A | 2 ± 0.2 A | 0.3 ± 0.03 B | 0.4 ± 0 B |
| Depth (cm) | Sand (%) | Silt (%) | Clay (%) | Soil pH | EC1:5 (dS·m−1) | Cl (g·kg−1) | Na2O (g·kg−1) | OM (%) | N (%) | P2O5 (mg·kg−1) | K2O (g·kg−1) | MgO (g·kg−1) | CaO (g·kg−1) | Zn (mg·kg−1) | Fe (mg·kg−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0–20 | 61.8 | 18.6 | 18.6 | 8.47 | 1.91 | 2.12 | 2.00 | 0.47 | 0.03 | 44.12 | 0.33 | 0.92 | 9.73 | 0.80 | 1.23 |
| 20–40 | 71.3 | 12.9 | 23.8 | 8.47 | 1.80 | 1.43 | 1.51 | 0.40 | 0.03 | 36.29 | 0.31 | 0.85 | 9.46 | 0.80 | 1.23 |
| Climatic Parameters | During the Experimental Period | 3 Years Average | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| March | April | May | June | July | March | April | May | June | July | |
| Temperature (°C) | 18.3 | 19 | 20.1 | 21.6 | 23 | 23.8 | 24 | 25.4 | 26.4 | 28.7 |
| Rainfall (mm) | 7 | 2 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 |
| Humidity (%) | 59 | 62 | 64 | 68 | 69 | Data not available | ||||
| Sunshine hours (hour. day−1) | 8.9 | 9 | 9.3 | 9.6 | 9.7 | Data not available | ||||
| Water Content | EC | pH | Cations (meq·L−1) | Anions (meq·L−1) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (dS·m−1) | K+ | Na+ | Ca2+ | Mg2+ | Cl− | SO42− | NO3− | CO32− | HCO3− | ||
| Freshwater | 4.04 | 7.45 | 0.883 | 24.35 | 11.25 | 6.48 | 28.12 | 11.21 | 3.46 | 0.0 | 3.52 |
| Groundwater | 11.98 | 7.35 | 3.44 | 114.07 | 28.4 | 26.42 | 124.55 | 52.15 | 1.01 | 0.0 | 3.88 |
| Periods (10 Day) | March | April | May | June | July |
|---|---|---|---|---|---|
| 0–10 days | 20 | 18 | 27 | 27 | 9 |
| 11–20 days | -- (rain) | 18 | 27 | 18 | |
| 21–31 days | 18 | 27 | 27 | 9 | |
| Total irrigation (mm/month) | 38 | 63 | 81 | 54 | 9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouras, H.; Choukr-Allah, R.; Amouaouch, Y.; Bouaziz, A.; Devkota, K.P.; El Mouttaqi, A.; Bouazzama, B.; Hirich, A. How Does Quinoa (Chenopodium quinoa Willd.) Respond to Phosphorus Fertilization and Irrigation Water Salinity? Plants 2022, 11, 216. https://doi.org/10.3390/plants11020216
Bouras H, Choukr-Allah R, Amouaouch Y, Bouaziz A, Devkota KP, El Mouttaqi A, Bouazzama B, Hirich A. How Does Quinoa (Chenopodium quinoa Willd.) Respond to Phosphorus Fertilization and Irrigation Water Salinity? Plants. 2022; 11(2):216. https://doi.org/10.3390/plants11020216
Chicago/Turabian StyleBouras, Hamza, Redouane Choukr-Allah, Younes Amouaouch, Ahmed Bouaziz, Krishna Prasad Devkota, Ayoub El Mouttaqi, Bassou Bouazzama, and Abdelaziz Hirich. 2022. "How Does Quinoa (Chenopodium quinoa Willd.) Respond to Phosphorus Fertilization and Irrigation Water Salinity?" Plants 11, no. 2: 216. https://doi.org/10.3390/plants11020216
APA StyleBouras, H., Choukr-Allah, R., Amouaouch, Y., Bouaziz, A., Devkota, K. P., El Mouttaqi, A., Bouazzama, B., & Hirich, A. (2022). How Does Quinoa (Chenopodium quinoa Willd.) Respond to Phosphorus Fertilization and Irrigation Water Salinity? Plants, 11(2), 216. https://doi.org/10.3390/plants11020216