

Combined Impact of Excess Zinc and Cadmium on Elemental Uptake, Leaf Anatomy and Pigments, Antioxidant Capacity, and Function of Photosynthetic Apparatus in Clary Sage (Salvia sclarea L.)

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Growth Conditions, and Heavy Metal Treatments
2.2. Determination of Trace Element Concentrations by Inductively Coupled Plasma Mass Spectrometry (ICP-MS)
2.3. Light Microscopy and Leaf Thickness
2.4. Determination of Membrane Stability Index, Lipid Peroxidation, and Total Antioxidant Capacity
2.5. Analysis of Leaf Pigments and Total Phenolic Content
2.6. Chlorophyll Fluorescence Analysis
2.7. Oxygen Evolution Measurements
2.8. Measurements of P700 Redox State and Microviscosity
2.9. Statistics
3. Results
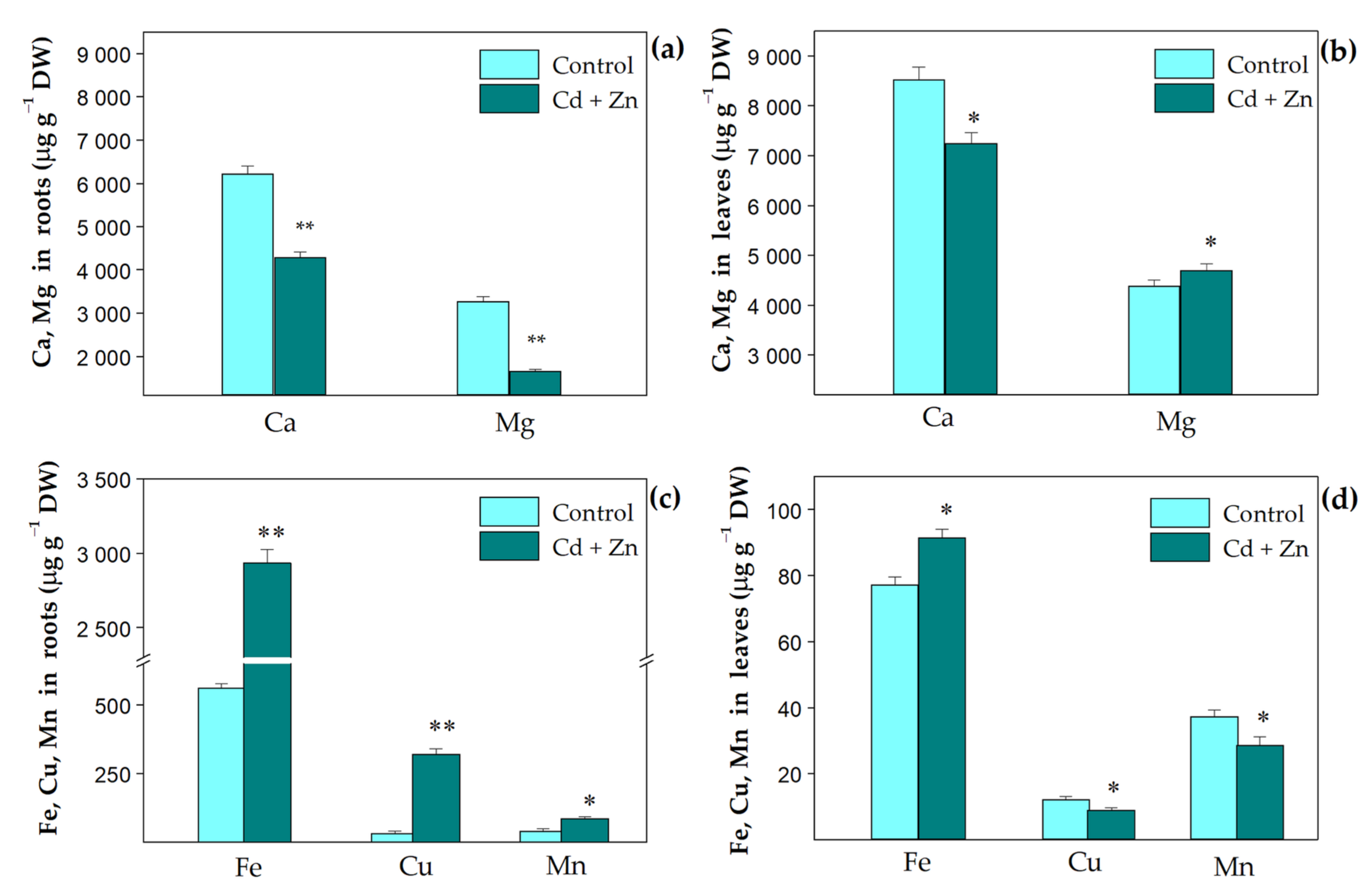
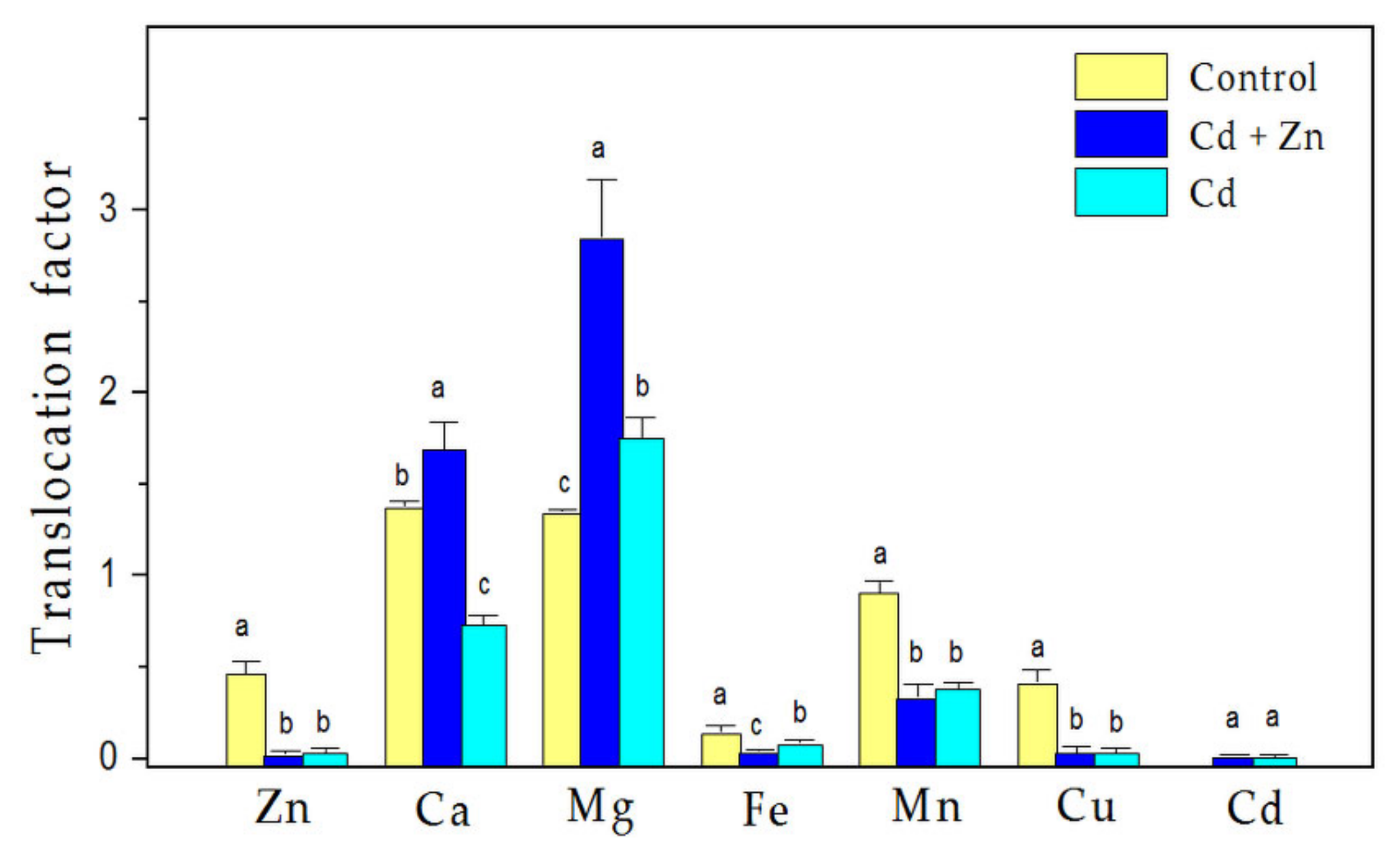
3.1. Trace Elemental Content in Plant Tissues
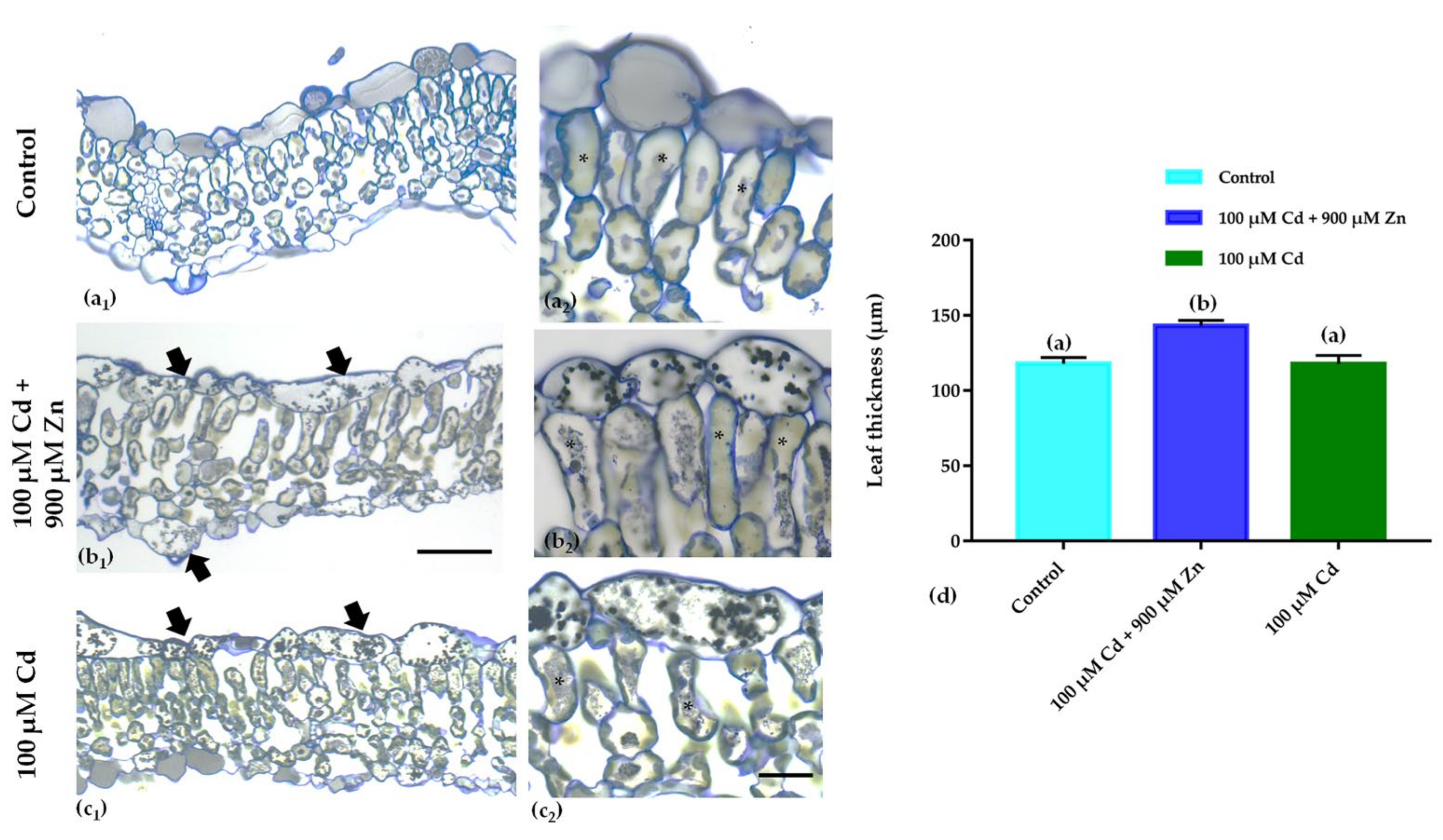
3.2. Leaf Anatomical Features
3.3. Membrane Damage and Antioxidant Capacity
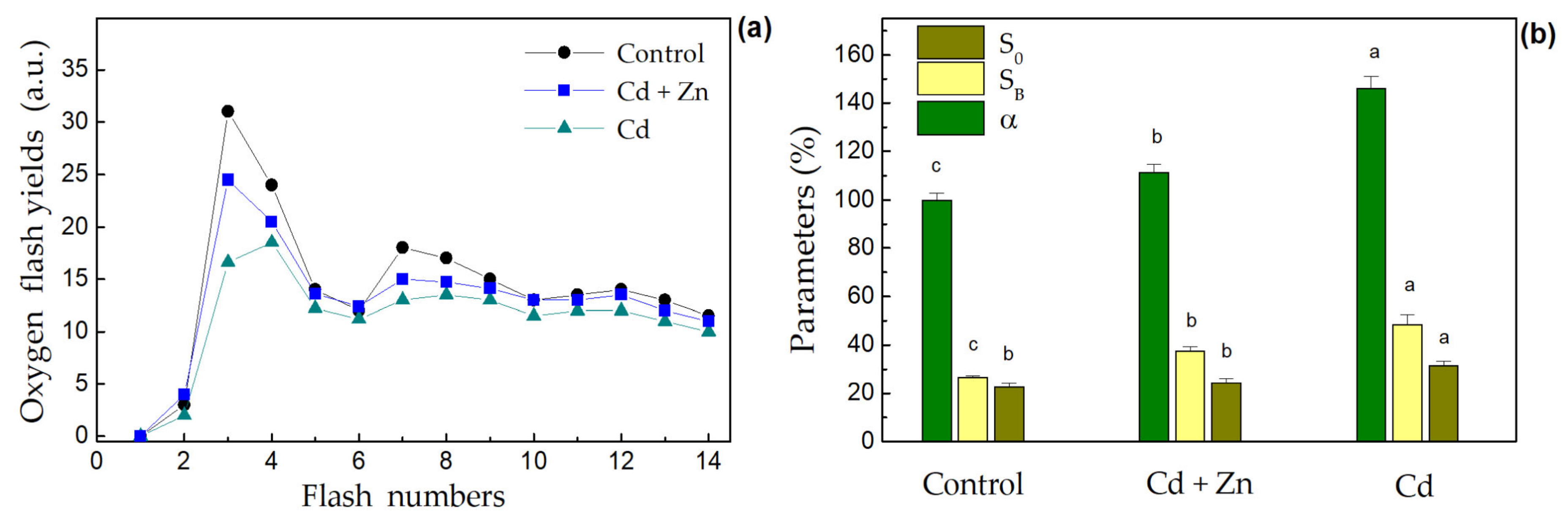
3.4. Photosynthetic Oxygen Evolution
3.5. Leaf Pigments and Total Phenolic Content
3.6. Changes in the Chlorophyll Fluorescence Parameters
3.7. P700 Photooxidation and Reduction Kinetics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alloway, B.J. Heavy Metals in Soils: Trace Metals and Metalloids in Soils and Their Bioavailability; Springer: Dordrecht, The Netherlands, 2013; pp. 1–614. [Google Scholar]
- Wu, F.; Fan, J.; Ye, X.; Yang, L.; Hu, R.; Ma, J.; Ma, S.; Li, D.; Zhou, J.; Nie, G.; et al. Unraveling cadmium toxicity in Trifolium repens L. seedling: Insight into regulatory mechanisms using comparative transcriptomics combined with physiological analyses. Int. J. Mol. Sci. 2022, 23, 4612. [Google Scholar] [CrossRef] [PubMed]
- Dobrikova, A.G.; Apostolova, E.L. Damage and protection of the photosynthetic apparatus under cadmium stress. In Cadmium Toxicity and Tolerance in Plants: From Physiology to Remediation, 1st ed.; Hasanuzzaman, M., Prasad, M.N.V., Fujita, M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 275–298. [Google Scholar]
- Zulfiqar, U.; Jiang, W.; Xiukang, W.; Hussain, S.; Ahmad, M.; Maqsood, M.F.; Ali, N.; Ishfaq, M.; Kaleem, M.; Haider, F.U.; et al. Cadmium Phytotoxicity, Tolerance, and Advanced Remediation Approaches in Agricultural Soils; A Comprehensive Review. Front. Plant Sci. 2022, 13, 773815. [Google Scholar] [CrossRef] [PubMed]
- Sandalio, L.M.; Dalurzo, H.C.; Gomez, M.; Romero-Puertas, M.C.; Del Rio, L.A. Cadmium-induced changes in the growth and oxidative metabolism of pea plants. J. Exp. Bot. 2001, 52, 2115–2126. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Abbas, T.; Zia-Ur-Rehman, M.; Hannan, F.; Keller, C.; Al-Wabel, M.I.; Ok, Y.S. Cadmium minimization in wheat: A critical review. Ecotoxicol. Environ. Saf. 2016, 130, 43–53. [Google Scholar] [CrossRef]
- Dobrikova, A.G.; Yotsova, E.K.; Börner, A.; Landjeva, S.P.; Apostolova, E.L. The wheat mutant DELLA-encoding gene (Rht-B1c) affects plant photosynthetic responses to cadmium stress. Plant Physiol. Biochem. 2017, 114, 10–18. [Google Scholar] [CrossRef]
- Adamakis, I.-D.S.; Sperdouli, I.; Hanc, A.; Dobrikova, A.; Apostolova, E.; Moustakas, M. Rapid hormetic responses of photosystem II photochemistry of clary sage to cadmium exposure. Int. J. Mol. Sci. 2021, 22, 41. [Google Scholar] [CrossRef]
- Djebali, W.; Zarrouk, M.; Brouquisse, R.; El Kahoui, S.; Limam, F.; Ghorbel, M.H.; Chaïbi, W. Ultrastructure and lipid alterations induced by cadmium in tomato (Lycopersicon esculentum) chloroplast membranes. Plant Biol. 2005, 7, 358–368. [Google Scholar] [CrossRef]
- Pal, M.; Lesko, K.; Janda, T.; Paldi, E.; Szalai, G. Cadmium-induced changes in the membrane lipid composition of maize plants. Cereal Res. Commun. 2007, 35, 1631–1642. [Google Scholar] [CrossRef]
- Pereira de Araújo, R.; Furtado de Almeida, A.A.; Silva Pereira, L.; Mangabeira, P.A.O.; Olimpio Souza, J.; Pirovani, C.P.; Ahnert, D.; Baligar, V.C. Photosynthetic, antioxidative, molecular and ultrastructural responses of young cacao plants to Cd toxicity in the soil. Ecotoxicol. Environ. Saf. 2017, 144, 148–157. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Rodríguez-Serrano, M.; Corpas, F.J.; Gómez, M.; del Río, L.A.; Sandalio, L.M. Cd-induced subcellular accumulation of O2.− and H2O2 in pea leaves. Plant Cell Environ. 2004, 27, 1122–1134. [Google Scholar] [CrossRef]
- Ahmad, A.; Hadi, F.; Ali, N. Effective phytoextraction of cadmium (Cd) with increasing concentration of total phenolics and free proline in Cannabis sativa (L) plant under various treatments of fertilizers, plant growth regulators and sodium salt. Int. J. Phytoremediat. 2015, 17, 56–65. [Google Scholar] [CrossRef]
- Clairvil, E.; Martins, J.P.R.; Braga, P.C.S.; Moreira, S.W.; Conde, L.T.; Cipriano, R.; Falqueto, A.R.; Gontijo, A.B.P.L. Zinc and cadmium as modulating factors of the morphophysiological responses of Alternanthera tenella Colla (Amaranthaceae) under in vitro conditions. Photosynthetica 2021, 59, 652–663. [Google Scholar] [CrossRef]
- Karahan, F.; Ozyigit, I.I.; Saracoglu, I.A.; Yalcin, I.E.; Ozyigit, A.H.; Ilcim, A. Heavy metal levels and mineral nutrient status in different parts of various medicinal plants collected from Eastern Mediterranean region of Turkey. Biol. Trace Elem. Res. 2020, 197, 316–329. [Google Scholar] [CrossRef]
- Kirkby, E. Introduction, definition and classification of nutrients. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, H., Ed.; Academic Press: Amsterdam, The Netherlands, 2012; pp. 3–5. [Google Scholar]
- Tsonev, T.; Lidon, F.J.C. Zinc in plants—An overview. Emir. J. Food Agric. 2012, 24, 322–333. [Google Scholar]
- Verma, S.K.; Sahin, G.; Das, A.K.; Gurel, E. In vitro plant regeneration of Ocimum basilicum L. is accelerated by zinc sulfate. Vitr. Cell. Dev. Biol.-Plant 2016, 52, 20–27. [Google Scholar] [CrossRef]
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc, zinc nanoparticles and plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef]
- Bayçu, G.; Gevrek-Kürüm, N.; Moustaka, J.; Csatári, I.; Rognes, S.E.; Moustakas, M. Cadmium-zinc accumulation and photosystem II responses of Noccaea caerulescens to Cd and Zn exposure. Environ. Sci. Pollut. Res. 2017, 24, 2840–2850. [Google Scholar] [CrossRef]
- Anjum, N.A.; Singh, H.P.; Khan, M.I.R.; Masood, A.; Per, T.S.; Negi, A.; Ahmad, I. Too much is bad—An appraisal of phytotoxicity of elevated plant-beneficial heavy metal ions. Environ. Sci. Pollut. Res. 2015, 22, 3361–3382. [Google Scholar] [CrossRef]
- Szopiński, M.; Sitko, K.; Gieroń, Ż.; Rusinowski, S.; Corso, M.; Hermans, C.; Verbruggen, N.; Małkowski, E. Toxic Effects of Cd and Zn on the photosynthetic apparatus of the Arabidopsis halleri and Arabidopsis arenosa pseudo-metallophytes. Front. Plant Sci. 2019, 10, 748. [Google Scholar] [CrossRef]
- Sarwar, N.; Ishaq, W.; Farid, G.; Shaheen, M.R.; Imran, M.; Geng, M.; Hussain, S. Zinc-cadmium interactions: Impact on wheat physiology and mineral acquisition. Ecotoxicol. Environ. Saf. 2015, 122, 528–536. [Google Scholar] [CrossRef]
- Saifullah, S.N.; Bibi, S.; Ahmad, M.; Ok, Y.S. Effectiveness of zinc application to minimize cadmium toxicity and accumulation in wheat (Triticum aestivum L.). Environ. Earth Sci. 2014, 71, 1663–1672. [Google Scholar] [CrossRef]
- Rashid, M.H.; Mahmudur, R.M.; Abdul, H.M.; Ravi, N. Growth, metal partitioning and antioxidant enzyme activities of mung beans as influenced by zinc oxide nanoparticles under cadmium stress. Crop Pasture Sci. 2022, 73, 862–876. [Google Scholar] [CrossRef]
- Adil, M.F.; Sehar, S.; Han, Z.; Lwalaba, J.L.W.; Jilani, G.; Zeng, F.; Chen, Z.H.; Shamsi, I.H. Zinc alleviates cadmium toxicity by modulating photosynthesis, ROS homeostasis, and cation flux kinetics in rice. Environm. Pollut. 2020, 265, 114979. [Google Scholar] [CrossRef]
- Mishra, B.; Chandra, M. Evaluation of phytoremediation potential of aromatic plants: A systematic review. J. Appl. Res. Med. Aromat. Plants 2022, 31, 100405. [Google Scholar] [CrossRef]
- Peana, A.T.; Moretti, M.D.L. Pharmacological activities and applications of Salvia sclarea and Salvia desoleana essential oils. Stud. Nat. Prod. Chem. 2002, 26, 391–423. [Google Scholar]
- Kuźma, L.; Kalemba, D.; Rózalski, M.; Rózalska, B.; Wieckowska-Szakiel, M.; Krajewska, U.; Wysokińska, H. Chemical composition and biological activities of essential oil from Salvia sclarea plants regenerated in vitro. Molecules 2009, 14, 1438–1447. [Google Scholar] [CrossRef]
- Gezek, G.; Hashemi, P.; Kalaycıoğlu, Z.; Kaygusuz, H.; Sarıoğlu, G.; Döker, S.; Dirmenci, T.; Bedia Erim, F. Evaluation of some Turkish Salvia species by principal component analysis based on their vitamin B2, mineral composition, and antioxidant properties. LWT 2019, 100, 287–293. [Google Scholar] [CrossRef]
- Dobrikova, A.; Apostolova, E.; Hanć, A.; Yotsova, E.; Borisova, P.; Sperdouli, I.; Adamakis, I.D.S.; Moustakas, M. Cadmium toxicity in Salvia sclarea L.: An integrative response of element uptake, oxidative stress markers, leaf structure and photosynthesis. Ecotoxicol. Environ. Saf. 2021, 209, 111851. [Google Scholar] [CrossRef] [PubMed]
- Dobrikova, A.; Apostolova, E.; Hanć, A.; Yotsova, E.; Borisova, P.; Sperdouli, I.; Adamakis, I.D.S.; Moustakas, M. Tolerance mechanisms of the aromatic and medicinal plant Salvia sclarea L. to excess zinc. Plants 2021, 10, 194. [Google Scholar]
- Sperdouli, I.; Adamakis, I.-D.S.; Dobrikova, A.; Apostolova, E.; Hanć, A.; Moustakas, M. Excess zinc supply reduces cadmium uptake and mitigates cadmium toxicity effects on chloroplast structure, oxidative stress and photosystem II photochemical efficiency in Salvia sclarea plants. Toxics 2022, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Angelova, V.R.; Ivanova, R.V.; Todorov, G.M.; Ivanov, K.I. Potential of Salvia sclarea L. for phytoremediation of soils contaminated with heavy metals. Int. J. Agric. Biosyst. Eng. 2016, 10, 780–790. [Google Scholar]
- Tufail, A.; Li, H.; Naeem, A.; Li, T.X. Leaf cell membrane stability-based mechanisms of zinc nutrient in mitigating salinity stress in rice. Plant Biol. 2018, 20, 338–345. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Fujita, M. Salicylic acid alleviates copper toxicity in rice (Oryza sativa L.) seedlings by up-regulating antioxidative and glyoxalase systems. Ecotoxicology 2013, 22, 959–973. [Google Scholar] [CrossRef]
- Vassilevska-Ivanova, R.L.; Shtereva, L.; Stancheva, I.; Geneva, M.; Hristozkova, M. Determination of the antioxidant capacity of Sideritis scardica specimens collected at different regions, C.R. Acad. Bulg. Sci. 2010, 69, 1307–1315. [Google Scholar]
- Sripakdee, T.; Sriwicha, A.; Jansam, N.; Mahachai, R.; Chanthai, S. Determination of total phenolics and ascorbic acid related to an antioxidant activity and thermal stability of the Mao fruit juice. Int. Food Res. J. 2015, 22, 618–624. [Google Scholar]
- Moustaka, J.; Panteris, E.; Adamakis, I.D.S.; Tanou, G.; Giannakoula, A.; Eleftheriou, E.P.; Moustakas, M. High anthocyanin accumulation in poinsettia leaves is accompanied by thylakoid membrane unstacking, acting as a photoprotective mechanism, to prevent ROS formation. Environ. Exp. Bot. 2018, 154, 44–55. [Google Scholar] [CrossRef]
- Schreiber, U.; Bilger, W.; Neubauer, C. Chlorophyll fluorescence as a non-intrusive indicator for rapid assessment of in vivo photosynthesis. In Ecophysiology of Photosynthesis; Schulze, E.D., Caldwell, M.M., Eds.; Series Ecological Studies; Springer: Berlin/Heidelberg, Germany, 1994; Volume 100, pp. 49–70. [Google Scholar]
- Moustakas, M.; Bayçu, G.; Sperdouli, I.; Eroğlu, H.; Eleftheriou, E.P. Arbuscular mycorrhizal symbiosis enhances photosynthesis in the medicinal herb Salvia fruticosa by improving photosystem II photochemistry. Plants 2020, 9, 962. [Google Scholar] [CrossRef]
- Sperdouli, I.; Andreadis, S.; Moustaka, J.; Panteris, E.; Tsaballa, A.; Moustakas, M. Changes in light energy utilization in photosystem II and reactive oxygen species generation in potato leaves by the pinworm Tuta absoluta. Molecules 2021, 26, 2984. [Google Scholar] [CrossRef]
- Zeinalov, Y. An equipment for investigations of photosynthetic oxygen production reactions. Bulg. J. Plant Physiol. 2002, 28, 57–67. [Google Scholar]
- Lubitz, W.; Chrysina, M.; Cox, N. Water oxidation in photosystem II. Photosynth. Res. 2019, 142, 105–125. [Google Scholar] [CrossRef]
- Zeinalov, Y. Photosynthesis—Behind the Fundamental Concepts; LAP Lambert Academic Publishing AG & Co. KG: Saarbrucken, Germany, 2010; pp. 1–128. [Google Scholar]
- Yotsova, E.K.; Dobrikova, A.G.; Stefanov, M.A.; Kouzmanova, M.; Apostolova, E.L. Improvement of the rice photosynthetic apparatus defence under cadmium stress modulated by salicylic acid supply to roots. Theor. Exp. Plant Physiol. 2018, 30, 57–70. [Google Scholar] [CrossRef]
- Dobrikova, A.G.; Vladkova, R.S.; Rashkov, G.D.; Todinova, S.J.; Krumova, S.B.; Apostolova, E.L. Effects of exogenous 24-epibrassinolide on the photosynthetic membranes under non-stress conditions. Plant Physiol. Biochem. 2014, 80, 75–82. [Google Scholar] [CrossRef]
- Dobrikova, A.G.; Apostolova, E.L. Damage and protection of the photosynthetic apparatus from UV-B radiation. II. Effect of quercetin at different pH. J. Plant Physiol. 2015, 184, 98–105. [Google Scholar] [CrossRef]
- Chaffai, R.; Koyama, H. Heavy metal tolerance in Arabidopsis thaliana. Adv. Bot. Res. 2011, 60, 1–49. [Google Scholar]
- Andresen, E.; Peiter, E.; Küpper, H. Trace metal metabolism in plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef]
- Sarwar, N.; Imran, M.; Shaheen, M.R.; Ishaque, W.; Kamran, M.A.; Matloob, A.; Rehim, A.; Hussain, S. Phytoremediation strategies for soils contaminated with heavy metals: Modifications and future perspectives. Chemosphere 2017, 171, 710–721. [Google Scholar] [CrossRef]
- Yang, Y.; Sun, C.; Yao, Y.; Zhang, Y.; Achal, V. Growth and physiological responses of grape (Vitis vinifera “Combier”) to excess zinc. Acta Physiol. Plant 2011, 33, 1483–1491. [Google Scholar] [CrossRef]
- Melki, F.; Zribi, O.T.; Jeder, S.; Louati, F.; Nouairi, I.; Mhadhbi, H.; Zribi, K. Effect of increasing zinc levels on Trigonella foenum-graecum growth and photosynthesis activity. J. Appl. Bot. Food Qual. 2022, 95, 23–30. [Google Scholar] [CrossRef]
- Szabò, I.; Spetea, C. Impact of the ion transportome of chloroplasts on the optimization of photosynthesis. J. Exp. Bot. 2017, 68, 3115–3128. [Google Scholar] [CrossRef]
- Saito, A.; Iino, T.; Sonoike, K.; Miwa, E.; Higuchi, K. Remodeling of the major light-harvesting antenna protein of PSII protects the young leaves of barley (Hordeum vulgare L.) from photoinhibition under prolonged iron deficiency. Plant Cell Physiol. 2010, 51, 2013–2030. [Google Scholar] [CrossRef]
- Sebastian, A.; Prasad, M.N.V. Iron- and manganese-assisted cadmium tolerance in Oryza sativa L.: Lowering of rhizotoxicity next to functional photosynthesis. Planta 2015, 241, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Hochmal, A.K.; Schulze, S.; Trompelt, K.; Hippler, M. Calcium dependent regulation of photosynthesis. Biochim. Biophys. Acta 2015, 1847, 993–1003. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Sarwat, M.; Bhat, N.A.; Wani, M.R.; Kazi, A.G.; Tran, L.S.P. Alleviation of cadmium toxicity in Brassica juncea L. (Czern. & Coss.) by calcium application involves various physiological and biochemical strategies. PLoS ONE 2015, 10, e0114571. [Google Scholar]
- Aravind, P.; Prasad, M.N.V. Zinc alleviates cadmium-induced oxidative stress in Ceratophyllum demersum L.: A free floating freshwater macrophyte. Plant Physiol. Biochem. 2003, 41, 391–397. [Google Scholar] [CrossRef]
- Wei, C.; Jiao, Q.; Agathokleous, E.; Liu, H.; Li, G.; Zhang, J.; Fahad, S.; Jiang, Y. Hormetic effects of zinc on growth and antioxidant defense system of wheat plants. Sci. Total Environ. 2022, 807, 150992. [Google Scholar] [CrossRef]
- Roby, M.H.H.; Sarhan, M.A.; Selim, K.A.-H.; Khalel, K.I. Evaluation of antioxidant activity, total phenols and phenolic compounds in thyme (Thymus vulgaris L.), sage (Salvia officinalis L.), and marjoram (Origanum majorana L.) extracts. Ind. Crops Prod. 2013, 43, 827–831. [Google Scholar] [CrossRef]
- Unuigbe, C.; Enahoro, J.; Erharuyi, O.; Okeri, H.A. Phytochemical analysis and antioxidant evaluation of lemon grass (Cymbopogon citratus DC.) Stapf leaves. J. Appl. Sci. Environ. Manag. 2019, 23, 223–228. [Google Scholar] [CrossRef]
- Francik, S.; Francik, R.; Sadowska, U.; Bystrowska, B.; Zawiślak, A.; Knapczyk, A.; Nzeyimana, A. Identification of phenolic compounds and determination of antioxidant activity in extracts and infusions of Salvia leaves. Materials 2020, 13, 5811. [Google Scholar] [CrossRef]
- Salinitro, M.; Hoogerwerf, S.; Casolari, S.; Zappi, A.; Melucci, D.; Tassoni, A. Production of antioxidant molecules in Polygonum aviculare (L.) and Senecio vulgaris (L.) under metal stress: A possible tool in the evaluation of plant metal tolerance. Int. J. Mol. Sci. 2020, 21, 7317. [Google Scholar] [CrossRef]
- Vidal, C.; Ruiz, A.; Ortiz, J.; Larama, G.; Perez, R.; Santander, C.; Ferreira, P.; Cornejo, P. Antioxidant responses of phenolic compounds and immobilization of copper in Imperata Cylindrica, a plant with potential use for bioremediation of Cu contaminated environments. Plants 2020, 9, 1397. [Google Scholar] [CrossRef]
- Lavid, N.; Schwartz, A.; Yarden, O.; Tel-Or, A. The involvement of polyphenols and peroxidase activities in heavy metal accumulation by epidermal glands of waterlily (Nympheaceae). Planta 2001, 212, 323–331. [Google Scholar] [CrossRef]
- Salinitro, M.; van der Ent, A.; Tognacchini, A.; Tassoni, A. Stress responses and nickel and zinc accumulation in different accessions of Stellaria media (L.) Vill. in response to solution pH variation in hydroponic culture. Plant Physiol. Biochem. 2020, 148, 133–141. [Google Scholar] [CrossRef]
- Sytar, O.; Kumar, A.; Latowski, D.; Kuczynska, P.; Strzałka, K.; Prasad, M. Heavy metal-induced oxidative damage, defense reactions, and detoxification mechanisms in plants. Acta Physiol. Plant. 2013, 35, 985–999. [Google Scholar] [CrossRef]
- Moustaka, J.; Tanou, G.; Giannakoula, A.; Adamakis, I.D.S.; Panteris, E.; Eleftheriou, E.P.; Moustakas, M. Anthocyanin accumulation in poinsettia leaves and its functional role in photo-oxidative stress. Environ. Exp. Bot. 2020, 175, 104065. [Google Scholar] [CrossRef]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Env. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Zoufan, P.; Azad, Z.; Ghahfarokhie, A.R.; Kolahi, M. Modification of oxidative stress through changes in some indicators related to phenolic metabolism in Malva parviflora exposed to cadmium. Ecotoxicol. Environ. Saf. 2020, 187, 109811. [Google Scholar] [CrossRef]
- Lu, Y.; Foo, L.Y. Antioxidant activities of polyphenols from sage (Salvia officinalis). Food Chem. 2001, 75, 197–202. [Google Scholar] [CrossRef]
- Goupy, P.; Dufour, C.; Loonis, M.; Dangles, O. Quantitative kinetic analysis of hydrogen transfer reactions from dietary polyphenols to the DPPH radical. J. Agric. Food Chem. 2003, 51, 615. [Google Scholar] [CrossRef]
- Goli, A.H.; Barzegar, M.; Sahari, M.A. Antioxidant activity, total phenolic compounds of pistachio (Pistachia vera) hull extracts. Food Chem. 2004, 92, 521–525. [Google Scholar] [CrossRef]
- Eyob, S.; Martinsen, B.K.; Tsegaye, A. Antioxidant and antimicrobial activities of extract and essential oil of korarima Aframomum corrorima (Braun) P.C.M. Jansen). Afr. J. Biotechnol. 2008, 7, 2585–2592. [Google Scholar]
- Maizura, M.; Aminah, A.; Wan Aida, W.M. Total phenolic content and antioxidant activity of kesum (Polygonum minus), ginger (Zingiber officinale) and turmeric (Curcuma longa) extract. Int. Food Res. J. 2001, 18, 534–538. [Google Scholar]
- Amarowicz, R.; Pegg, R.B.; Rahimi-Moghaddam, P.; Barl, B.; Weil, J.A. 4Free-radical scavenging capacity and antioxidant activity of selected plant species from the Canadian prairies. Food Chem. 2004, 84, 551–562. [Google Scholar] [CrossRef]
- Phang, C.; Malek, S.N.A.; Ibrahim, H.; Wahab, N.A. Antioxidant properties of crude and fractionated extracts of Alpinia mutica rhizomes and their total phenolic content. Afr. J. Pharm. Pharmacol. 2011, 5, 842–852. [Google Scholar]
- Stancheva, I.; Geneva, M.; Hristozkova, M.; Markovska, Y.; Salamon, I. Antioxidant capacity of sage grown on heavy metal-polluted soil. Russ. J. Plant Physiol. 2010, 57, 799–805. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Nisha, N.; Ejaz, B.; Khan, M.I.R.; Kumar, M.; Ramteke, P.W.; Gupta, R. A comprehensive review on the heavy metal toxicity and sequestration in plants. Biomolecules 2021, 12, 43. [Google Scholar] [CrossRef]
- Vollenweider, P.; Cosio, C.; Günthardt-Goerg, M.S.; Keller, C. Localization and effects of cadmium in leaves of a cadmium-tolerant willow (Salix viminalis L.): Part II Microlocalization and cellular effects of cadmium. Environ. Exp. Bot. 2006, 58, 25–40. [Google Scholar] [CrossRef]
- Fukuda, N.; Hokura, A.; Kitajima, N.; Terada, Y.; Saito, H.; Abe, T.; Nakai, I. Micro x-ray fluorescence imaging and micro x-ray absorption spectroscopy of cadmium hyper-accumulating plant, Arabidopsis halleri ssp. gemmifera, using high-energy synchrotron radiation. J. Anal. Atom. Spectrom. 2008, 23, 1068–1075. [Google Scholar] [CrossRef]
- Vogel-Mikus, K.; Regvar, M.; Mesjasz-Przybyłowicz, J.; Przybyłowicz, W.J.; Simcic, J.; Pelicon, P.; Budnar, M. Spatial distribution of cadmium in leaves of metal hyperaccumulating Thlaspi praecox using micro-PIXE. New Phytol. 2008, 179, 712–721. [Google Scholar] [CrossRef]
- Tian, S.; Lu, L.; Labavitch, J.; Yang, X.; He, Z.; Hu, H.; Sarangi, R.; Newville, M.; Commisso, J.; Brown, P. Cellular sequestration of cadmium in the hyperaccumulator plant species Sedum alfredii. Plant Physiol. 2011, 157, 1914–1925. [Google Scholar] [CrossRef]
- Petrovic, D.; Krivokapic, S. The effect of Cu, Zn, Cd, and Pb accumulation on biochemical parameters (proline, chlorophyll) in the water caltrop (Trapa natans L.), Lake Skadar, Montenegro. Plants 2020, 9, 1287. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, L.; Huang, X.; Zhou, Y.; Quan, Q.; Li, Y.; Zhu, X. Response of photosynthesis to different concentrations of heavy metals in Davidia involucrata. PLoS ONE 2020, 15, e0228563. [Google Scholar] [CrossRef]
- Björkman, O.; Demmig, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 1987, 170, 489–504. [Google Scholar] [CrossRef]
- Yi, Z.; Cui, J.; Fu, Y.; Liu, H. Effect of different light intensity on physiology, antioxidant capacity and photosynthetic characteristics on wheat seedlings under high CO2 concentration in a closed artificial ecosystem. Photosynth. Res. 2020, 144, 23–34. [Google Scholar] [CrossRef]
- Schansker, G.; Goussias, C.; Petrouleas, V.; Rutherford, A.W. Reduction of the Mn cluster of the water-oxidizing enzyme by nitric oxide: Formation of an S-2 state. Biochemistry 2002, 41, 3057–3064. [Google Scholar] [CrossRef]
- Bukhov, N.; Egorova, E.; Carpentier, R. Electron flow to photosystem I from stromal reductants in vivo: The size of the pool of stromal reductants controls the rate of electron donation to both rapidly and slowly reducing photosystem I units. Planta 2002, 215, 812–820. [Google Scholar] [CrossRef]
- Albertsson, P.-A. The structure and function of the chloroplast photosynthetic membrane—A model for the domain organization. Photosynth. Res. 1995, 46, 141–149. [Google Scholar] [CrossRef]
- Nouairi, I.; Ghnaya, T.; Youssef, N.B.; Zarrouk, M.; Ghorbel, M.H. Changes in content and fatty acid profiles of total lipids of two halophytes: Sesuvium portulacastrum and Mesembryanthemum crystallinum under cadmium stress. J. Plant Physiol. 2006, 163, 1198–1202. [Google Scholar] [CrossRef]
- Chaffai, R.; Cherif, A. The cadmium-induced changes in the polar and neutral lipid compositions suggest the involvement of triacylglycerol in the defense response in maize. Physiol. Mol. Biol. Plants 2020, 26, 15–23. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment (8 Days) | MSI (%) | LP (%) | FRAP (μmol Fe2+ g−1 DW) | DPPH (%) |
|---|---|---|---|---|
| Control | 89.8 ± 0.5 a | 100 c | 61.87 ± 2.28 c | 48.38 ± 2.26 c |
| Cd + Zn | 74.2 ± 1.7 b | 124 ± 2 b | 97.26 ± 2.35 a | 67.03 ± 1.18 a |
| Cd | 56.9 ± 2.3 c | 141 ± 3 a | 83.15 ± 2.86 b | 64.35 ± 1.04 b |
| Treatment (8 Days) | Maximum Efficiency of PSII Photochemistry (Fv/Fm) | Efficiency of the Oxygen Evolving Complex (Fv/Fo) | Relative PSII Electron Transport Rate (ETR) |
|---|---|---|---|
| Control | 0.827 ± 0.007 a | 4.80 ± 0.25 a | 57.57 ± 1.53 b |
| Cd + Zn | 0.828 ± 0.012 a | 4.83 ± 0.41 a | 59.11 ± 2.76 a |
| Cd | 0.803 ± 0.016 b | 4.09 ± 0.39 b | 55.09 ± 3.52 c |
| Treatment (8 Days) | P700+ (ΔA/A830 × 10−3) | k1 (s−1) | k2 (s−1) | η |
|---|---|---|---|---|
| Control | 11.71 ± 0.38 b | 0.55 ± 0.07 c | 0.12 ± 0.02 c | 2.946 ± 0.11 c |
| Cd + Zn | 12.78 ± 0.44 a | 0.87 ± 0.09 b | 0.16 ± 0.01 b | 3.257 ± 0.15 b |
| Cd | 10.47 ± 0.27 c | 1.29 ± 0.05 a | 0.21 ± 0.01 a | 3.680 ± 0.17 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dobrikova, A.; Apostolova, E.; Adamakis, I.-D.S.; Hanć, A.; Sperdouli, I.; Moustakas, M. Combined Impact of Excess Zinc and Cadmium on Elemental Uptake, Leaf Anatomy and Pigments, Antioxidant Capacity, and Function of Photosynthetic Apparatus in Clary Sage (Salvia sclarea L.). Plants 2022, 11, 2407. https://doi.org/10.3390/plants11182407
Dobrikova A, Apostolova E, Adamakis I-DS, Hanć A, Sperdouli I, Moustakas M. Combined Impact of Excess Zinc and Cadmium on Elemental Uptake, Leaf Anatomy and Pigments, Antioxidant Capacity, and Function of Photosynthetic Apparatus in Clary Sage (Salvia sclarea L.). Plants. 2022; 11(18):2407. https://doi.org/10.3390/plants11182407
Chicago/Turabian StyleDobrikova, Anelia, Emilia Apostolova, Ioannis-Dimosthenis S. Adamakis, Anetta Hanć, Ilektra Sperdouli, and Michael Moustakas. 2022. "Combined Impact of Excess Zinc and Cadmium on Elemental Uptake, Leaf Anatomy and Pigments, Antioxidant Capacity, and Function of Photosynthetic Apparatus in Clary Sage (Salvia sclarea L.)" Plants 11, no. 18: 2407. https://doi.org/10.3390/plants11182407
APA StyleDobrikova, A., Apostolova, E., Adamakis, I.-D. S., Hanć, A., Sperdouli, I., & Moustakas, M. (2022). Combined Impact of Excess Zinc and Cadmium on Elemental Uptake, Leaf Anatomy and Pigments, Antioxidant Capacity, and Function of Photosynthetic Apparatus in Clary Sage (Salvia sclarea L.). Plants, 11(18), 2407. https://doi.org/10.3390/plants11182407