
Hydrogen Peroxide Mediates Premature Senescence Caused by Darkness and Inorganic Nitrogen Starvation in Physcomitrium patens

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
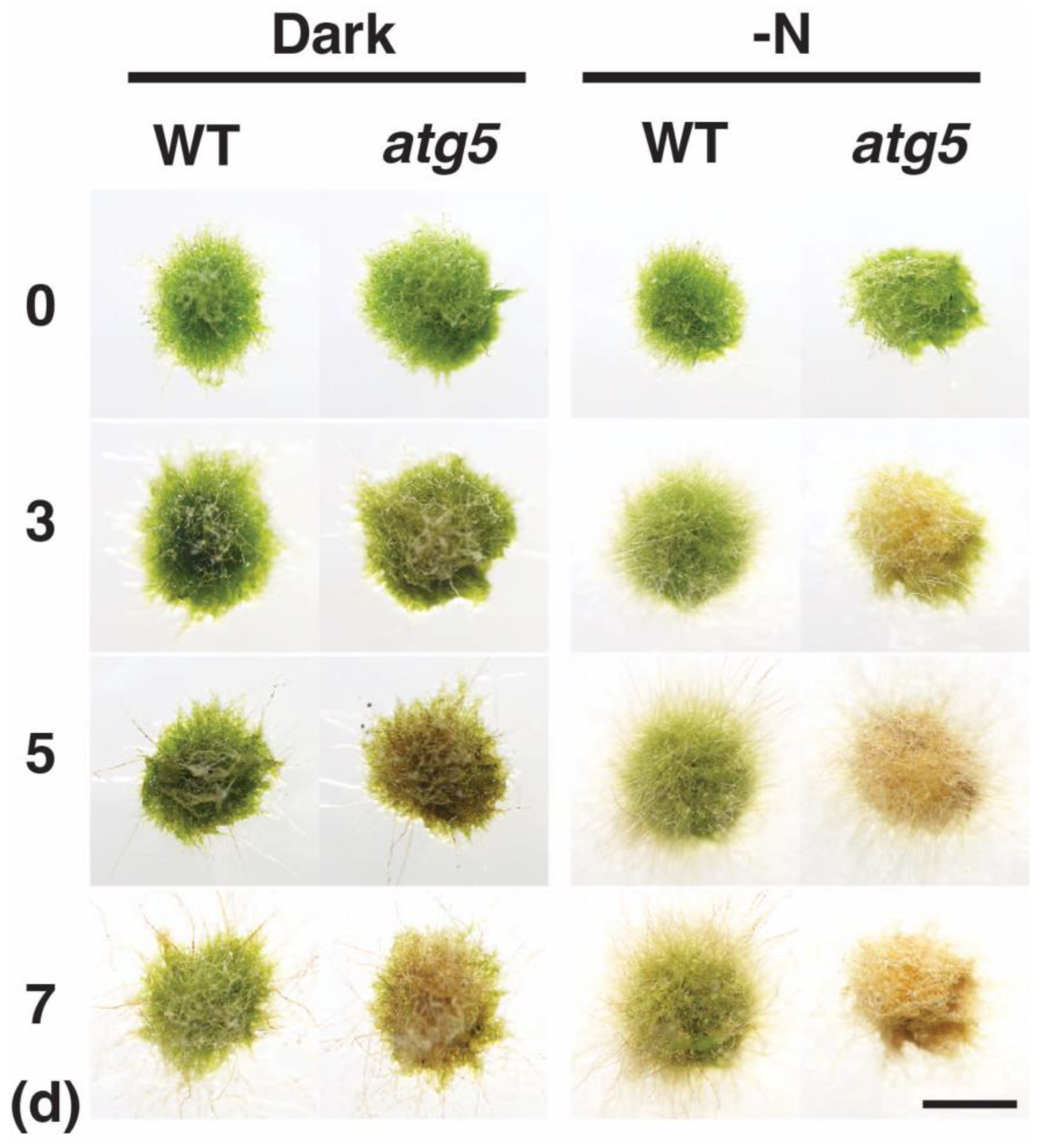
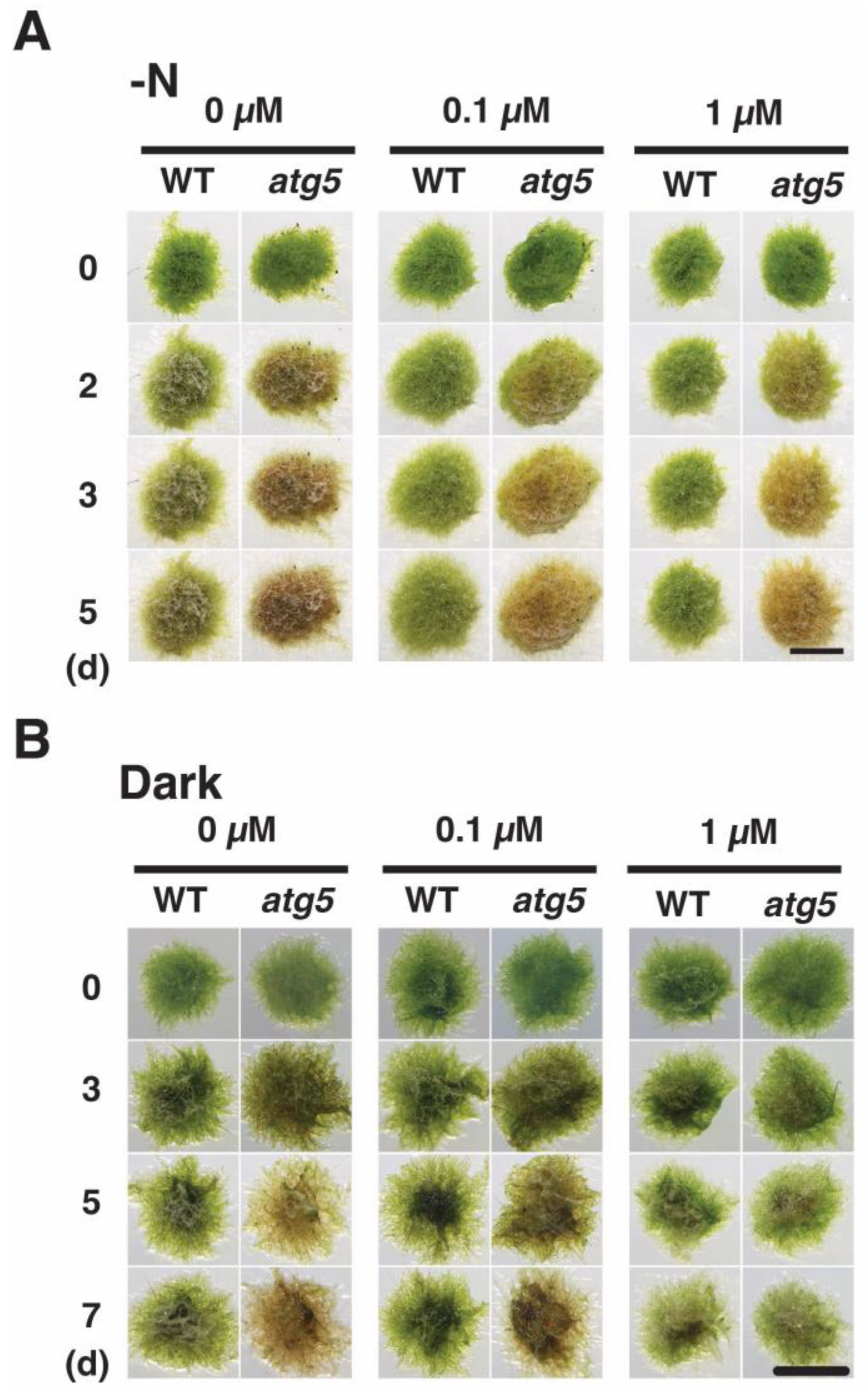
2.1. Premature Senescence Is Induced by Either Darkness or Inorganic Nitrogen Starvation in Physcomitrium
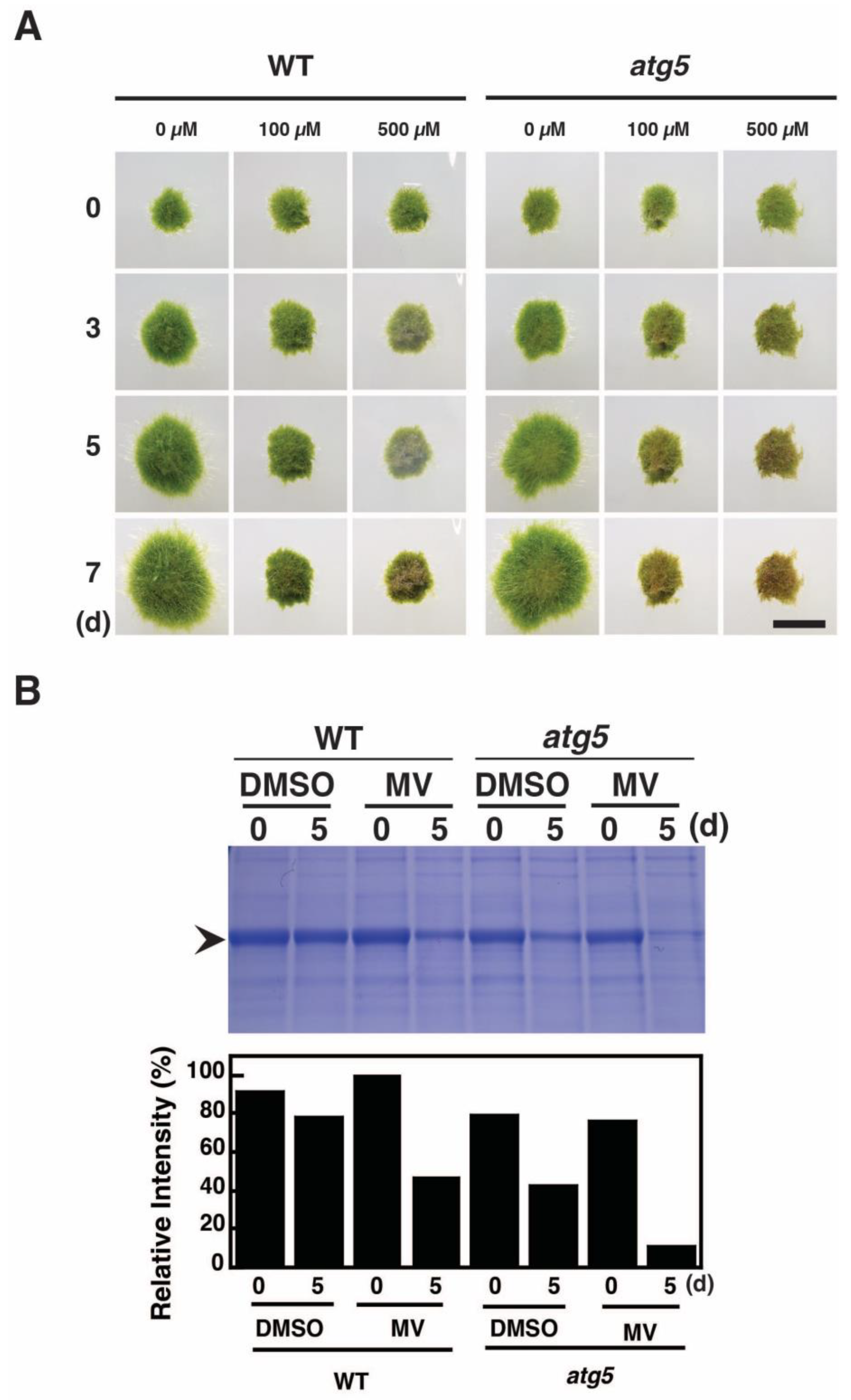
2.2. Methyl Viologen Induces Senescence-Like Symptoms
2.3. MV Treatment Increases Cellular H2O2 Levels
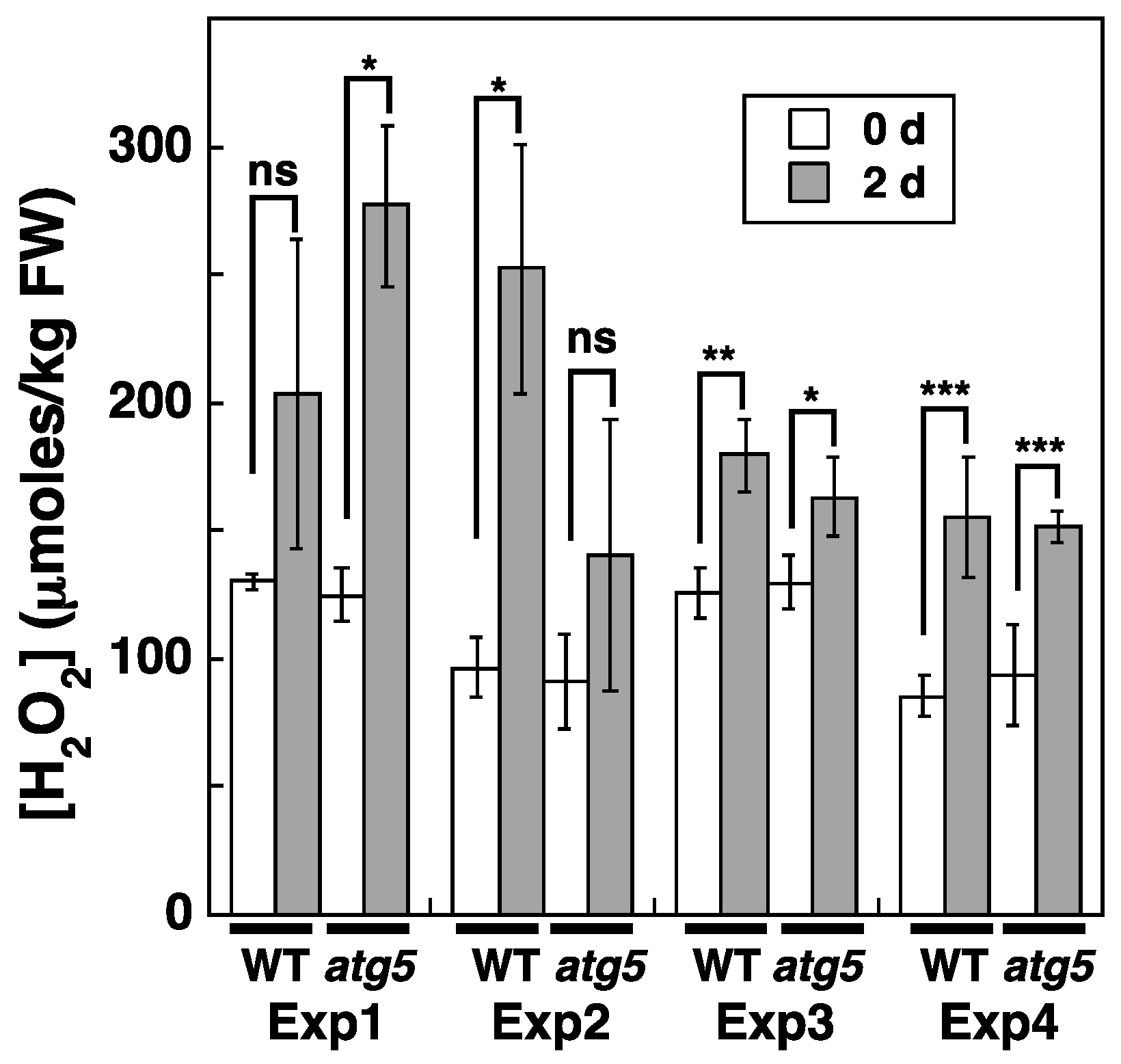
2.4. Cellular H2O2 Levels Increase under Dark Conditions
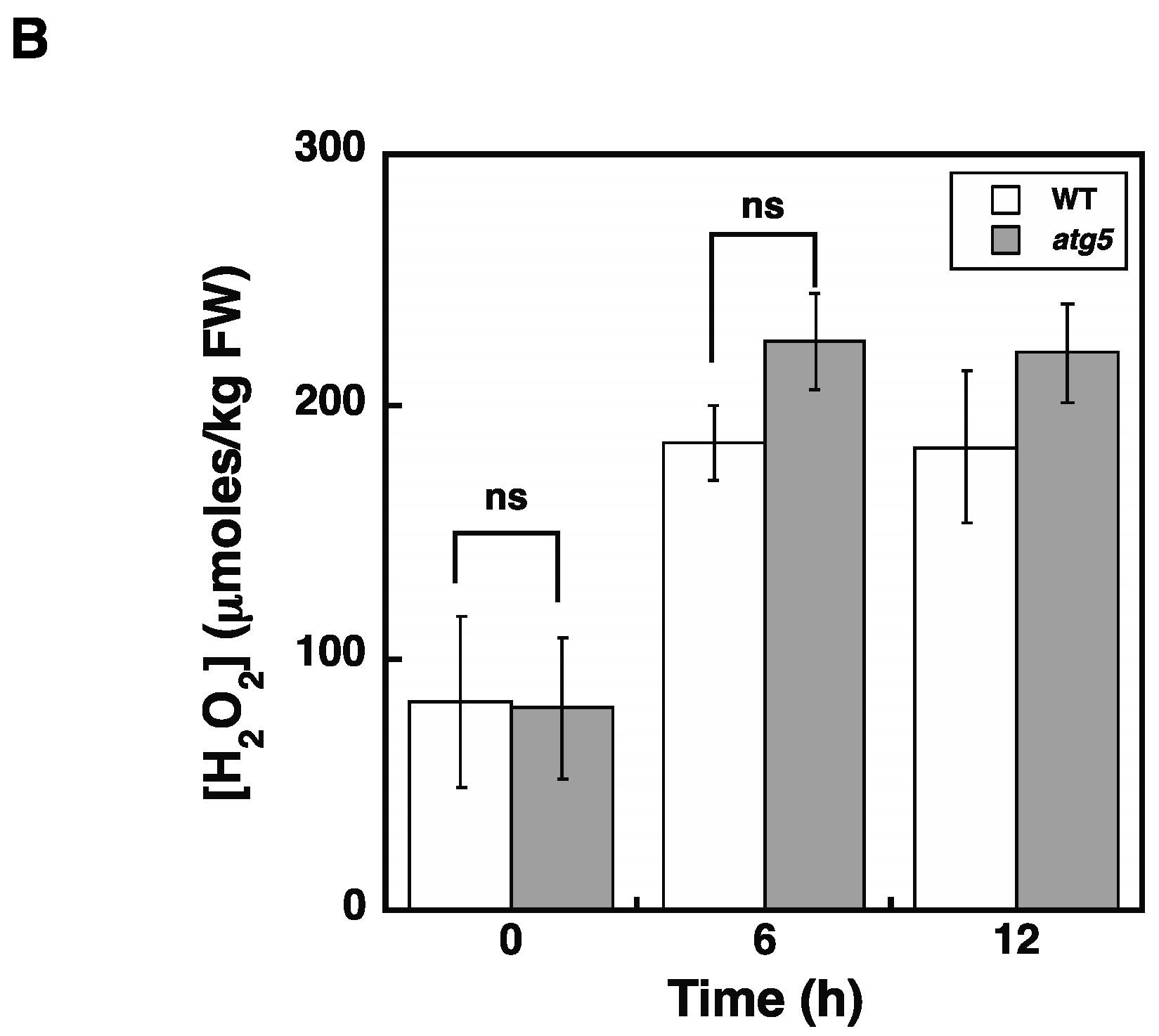
2.5. Cellular H2O2 Levels Increase Due to Inorganic Nitrogen Starvation
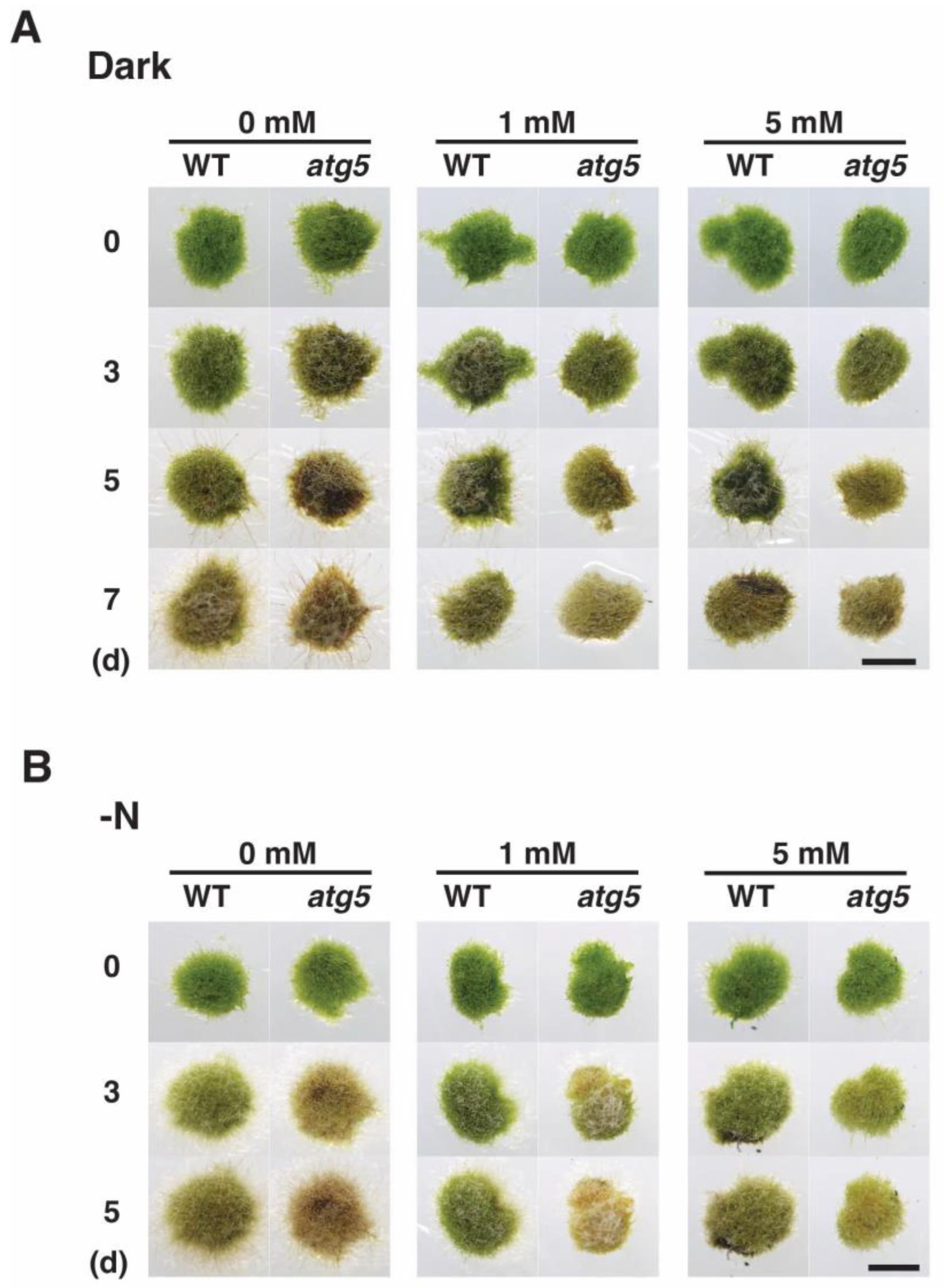
2.6. N-Acetylcysteine, an ROS Scavenger, Decelerates Yellowing
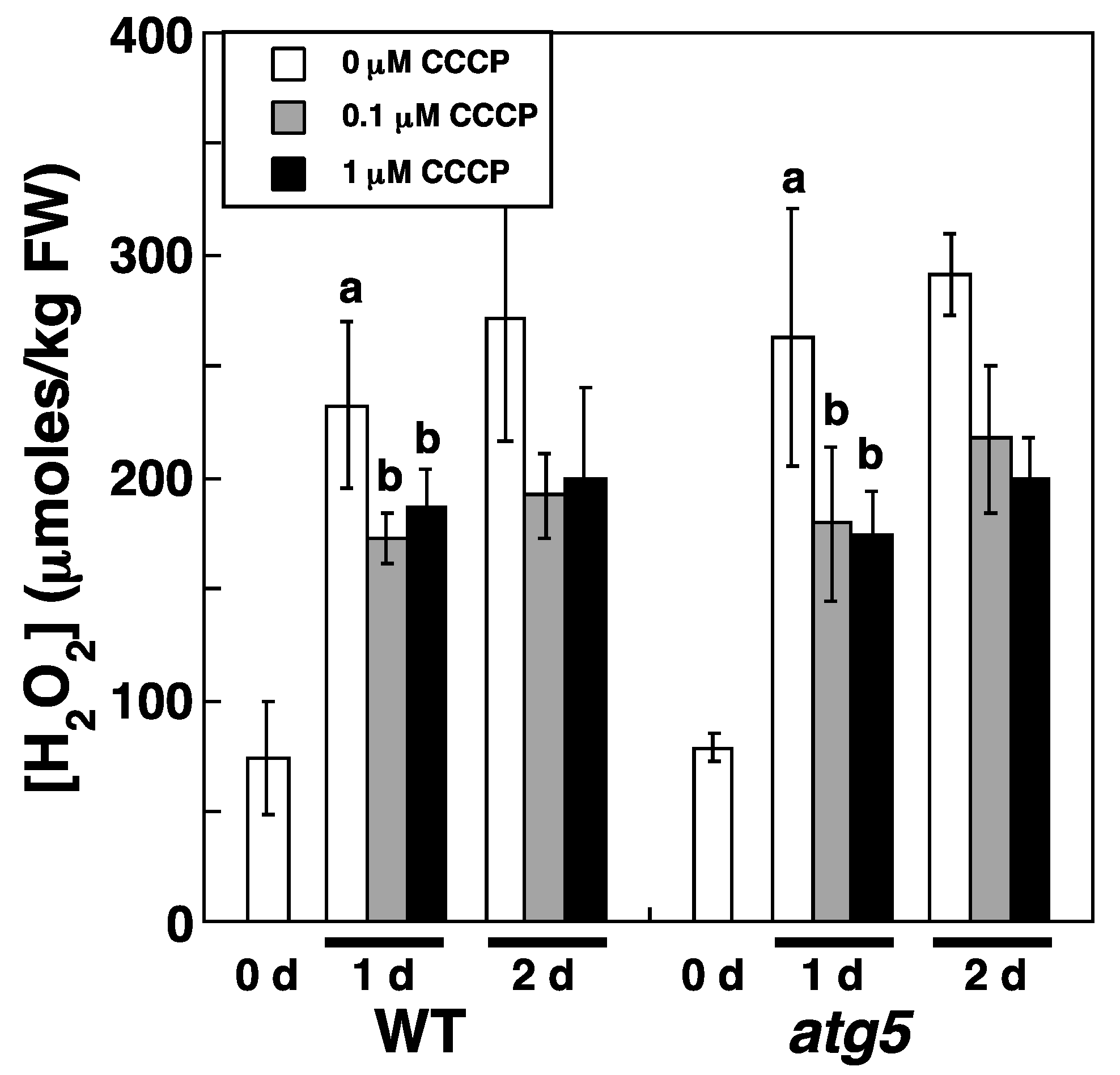
2.7. Carbonyl Cyanide m-Chlorophenylhydrazone Lowers H2O2 Levels and Decelerates Senescence under Nitrogen Starvation Conditions
2.8. Elevated H2O2 Levels Alone Cannot Explain the Difference in the Senescence Responses of WT and atg5 Colonies
3. Discussion
4. Materials and Methods
4.1. Biological Materials
4.2. H2O2 Measurement
4.3. SDS-PAGE
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Klionsky, D.J.; Ohsumi, Y. Vacuolar import of proteins and organelles from the cytoplasm. Annu. Rev. Cell Dev. Biol. 1999, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Levine, B.; Klionsky, D.J. Development by self-digestion: Molecular mechanisms and biological functions of autophagy. Dev. Cell 2004, 6, 463–477. [Google Scholar] [CrossRef]
- Lee, S.; Seo, P.J.; Lee, H.J.; Park, C.M. A NAC transcription factor NTL4 promotes reactive oxygen species production during drought-induced leaf senescence in Arabidopsis. Plant J. 2012, 70, 831–844. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-J.; Lin, Z.-W.; Huang, G.-J.; Lin, Y.-H. Sweet potato calmodulin SPCAM is involved in salt stress-mediated leaf senescence, H2O2 elevation and senescence-associated gene expression. J. Plant Physiol. 2012, 169, 1892–1902. [Google Scholar] [CrossRef] [PubMed]
- Bieker, S.; Riester, L.; Stahl, M.; Franzaring, J.; Zentgraf, U. Senescence-specific Alteration of Hydrogen Peroxide Levels in Arabidopsis thaliana and Oilseed Rape Spring Variety Brassica napus L. cv. MozartF. J. Integr. Plant Biol. 2012, 54, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Zentgraf, U.; Andrade-Galan, A.G.; Bieker, S. Specificity of H2O2 signaling in leaf senescence: Is the ratio of H2O2 contents in different cellular compartments sensed in Arabidopsis plants? Cell Mol. Biol. Lett. 2022, 27, 4. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, K.; Jikumaru, Y.; Kamiya, Y.; Kusano, M.; Consonni, C.; Panstruga, R.; Ohsumi, Y.; Shirasu, K. Autophagy negatively regulates cell death by controlling NPR1-dependent salicylic acid signaling during senescence and the innate immune response in Arabidopsis. Plant Cell 2009, 21, 2914–2927. [Google Scholar] [CrossRef] [PubMed]
- Lenz, H.D.; Haller, E.; Melzer, E.; Kober, K.; Wurster, K.; Stahl, M.; Bassham, D.C.; Vierstra, R.D.; Parker, J.E.; Bautor, J. Autophagy differentially controls plant basal immunity to biotrophic and necrotrophic pathogens. Plant J. 2011, 66, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.-X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Nxele, X.; Klein, A.; Ndimba, B. Drought and salinity stress alters ROS accumulation, water retention, and osmolyte content in sorghum plants. S. Afr. J. Bot. 2017, 108, 261–266. [Google Scholar] [CrossRef]
- Rensing, S.A.; Goffinet, B.; Meyberg, R.; Wu, S.Z.; Bezanilla, M. The moss Physcomitrium (Physcomitrella) patens: A model organism for non-seed plants. Plant Cell 2020, 32, 1361–1376. [Google Scholar] [CrossRef] [PubMed]
- Mukae, K.; Inoue, Y.; Moriyasu, Y. ATG5-knockout mutants of Physcomitrella provide a platform for analyzing the involvement of autophagy in senescence processes in plant cells. Plant Signal. Behav. 2015, 10, e1086859. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Brosché, M.; Shapiguzov, A.; He, X.-Q.; Vainonen, J.P.; Leppälä, J.; Trotta, A.; Kangasjärvi, S.; Salojärvi, J.; Kangasjärvi, J. Interaction of methyl viologen-induced chloroplast and mitochondrial signaling in Arabidopsis. Free Radic. Biol. Med. 2019, 134, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Ben Rejeb, K.; Lefebvre-De Vos, D.; Le Disquet, I.; Leprince, A.S.; Bordenave, M.; Maldiney, R.; Jdey, A.; Abdelly, C.; Savouré, A. Hydrogen peroxide produced by NADPH oxidases increases proline accumulation during salt or mannitol stress in Arabidopsis thaliana. New Phytol. 2015, 208, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, T.; Hiwatashi, Y.; Sakakibara, K.; Kato, M.; Hasebe, M. Tagged mutagenesis and gene-trap in the moss, Physcomitrella patens by shuttle mutagenesis. DNA Res. 2000, 7, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Gay, C.; Collins, J.; Gebicki, J.M. Determination of hydroperoxides by the ferric-xylenol orange method. Redox Rep. 1999, 4, 327–328. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roni, M.S.; Sakil, M.A.; Aktar, M.M.; Takatsuka, C.; Mukae, K.; Inoue-Aono, Y.; Moriyasu, Y. Hydrogen Peroxide Mediates Premature Senescence Caused by Darkness and Inorganic Nitrogen Starvation in Physcomitrium patens. Plants 2022, 11, 2280. https://doi.org/10.3390/plants11172280
Roni MS, Sakil MA, Aktar MM, Takatsuka C, Mukae K, Inoue-Aono Y, Moriyasu Y. Hydrogen Peroxide Mediates Premature Senescence Caused by Darkness and Inorganic Nitrogen Starvation in Physcomitrium patens. Plants. 2022; 11(17):2280. https://doi.org/10.3390/plants11172280
Chicago/Turabian StyleRoni, Md. Shyduzzaman, Md. Arif Sakil, Most Mohoshena Aktar, Chihiro Takatsuka, Kyosuke Mukae, Yuko Inoue-Aono, and Yuji Moriyasu. 2022. "Hydrogen Peroxide Mediates Premature Senescence Caused by Darkness and Inorganic Nitrogen Starvation in Physcomitrium patens" Plants 11, no. 17: 2280. https://doi.org/10.3390/plants11172280
APA StyleRoni, M. S., Sakil, M. A., Aktar, M. M., Takatsuka, C., Mukae, K., Inoue-Aono, Y., & Moriyasu, Y. (2022). Hydrogen Peroxide Mediates Premature Senescence Caused by Darkness and Inorganic Nitrogen Starvation in Physcomitrium patens. Plants, 11(17), 2280. https://doi.org/10.3390/plants11172280