
Marker-Trait Associations for Total Carotenoid Content and Individual Carotenoids in Durum Wheat Identified by Genome-Wide Association Analysis

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Genotyping
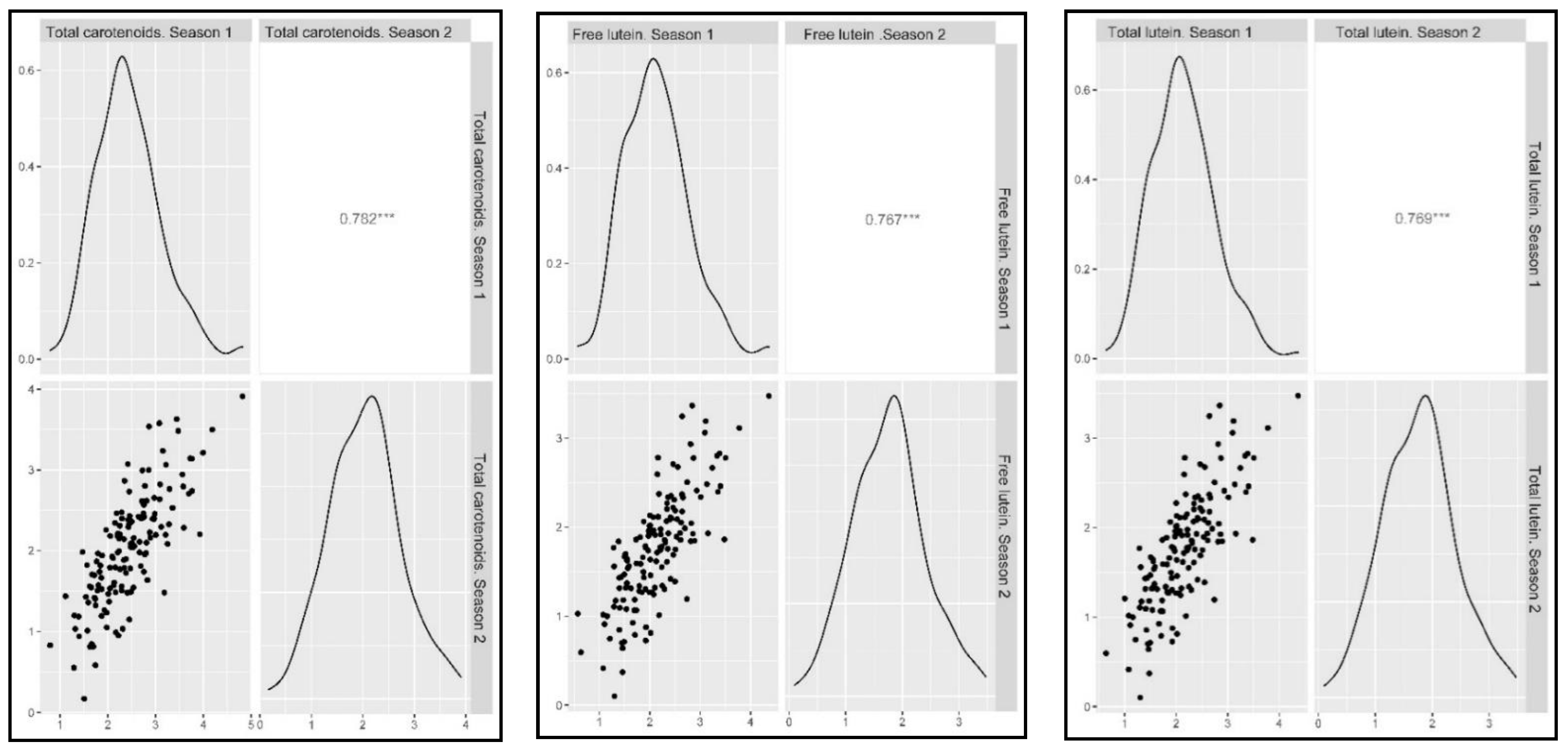
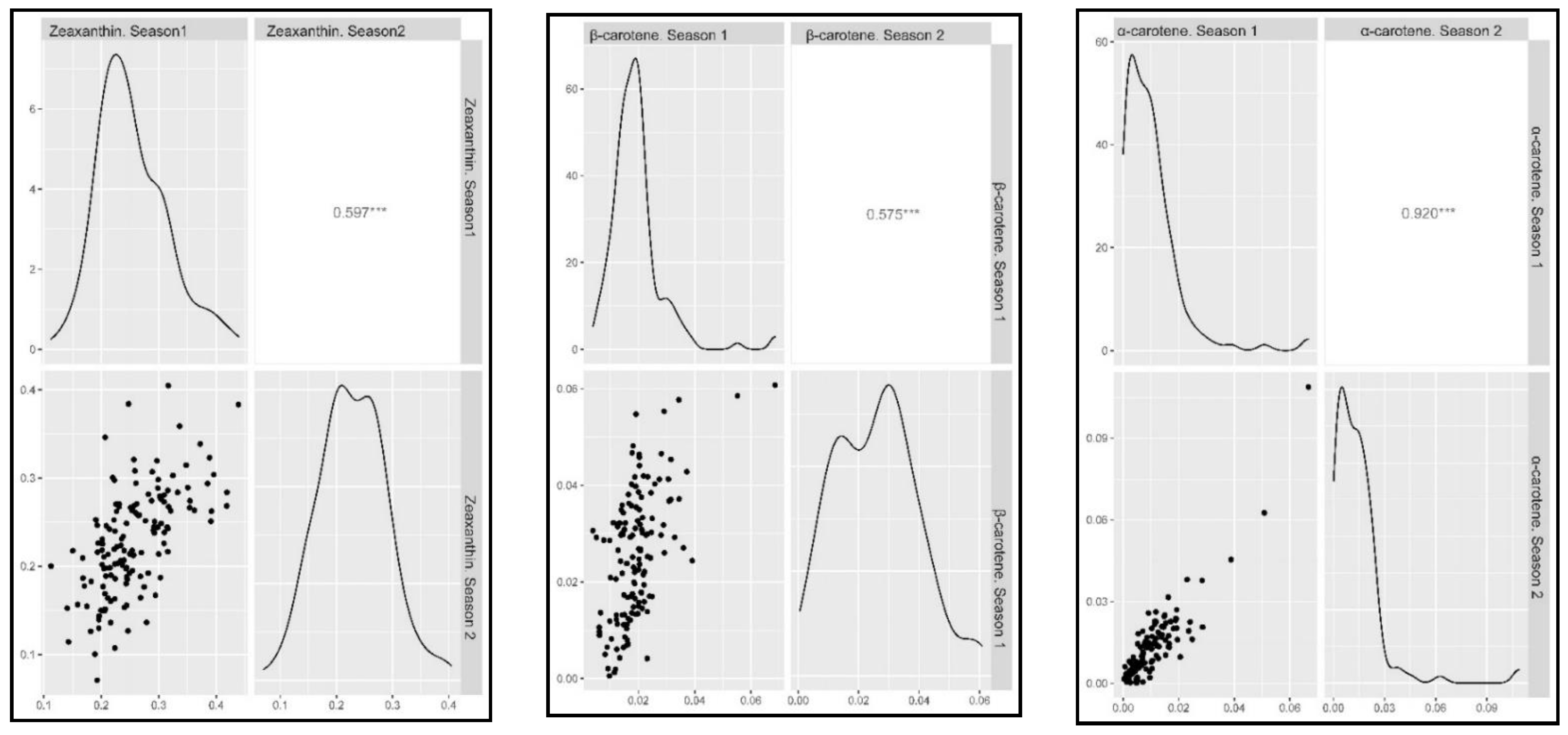
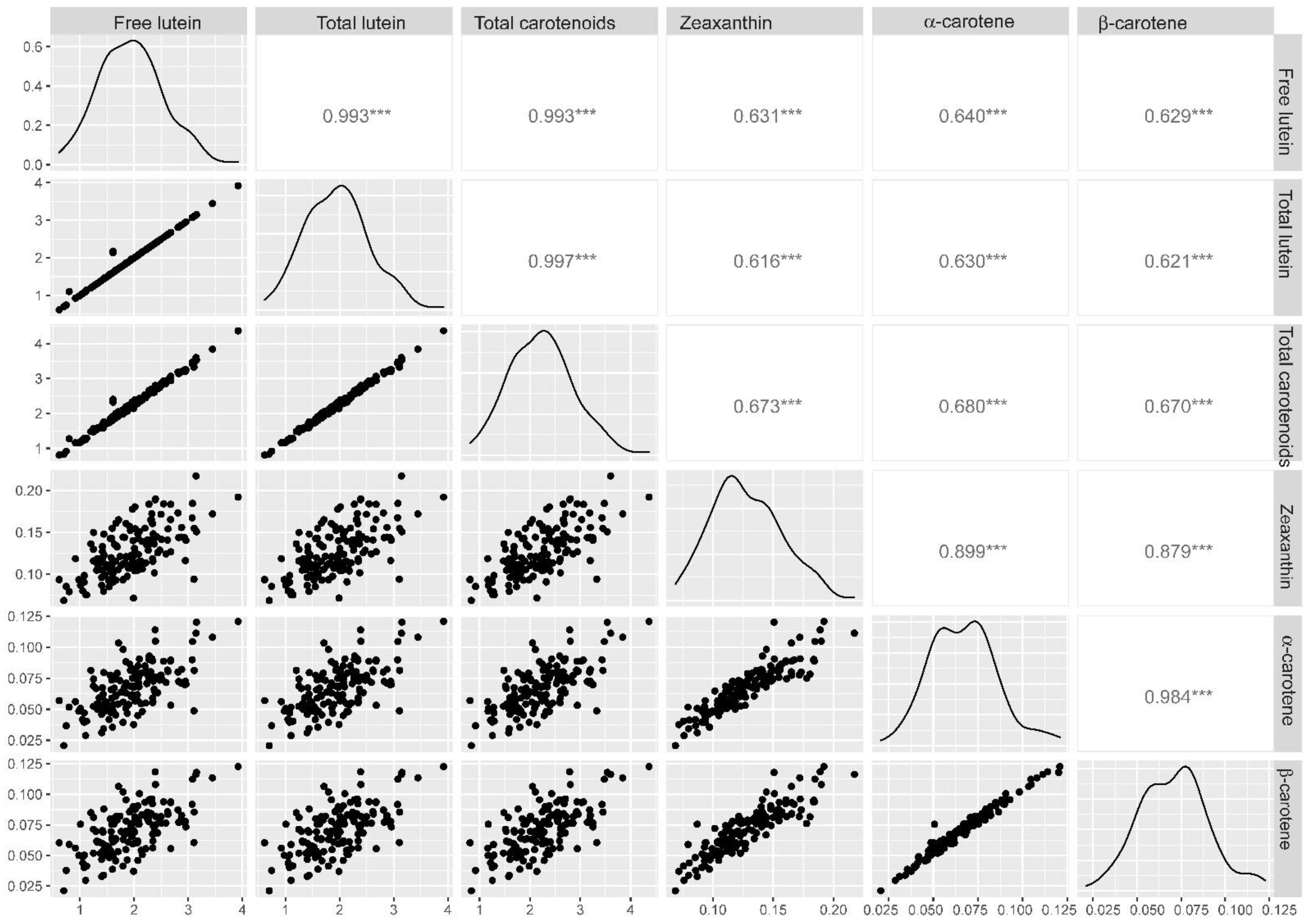
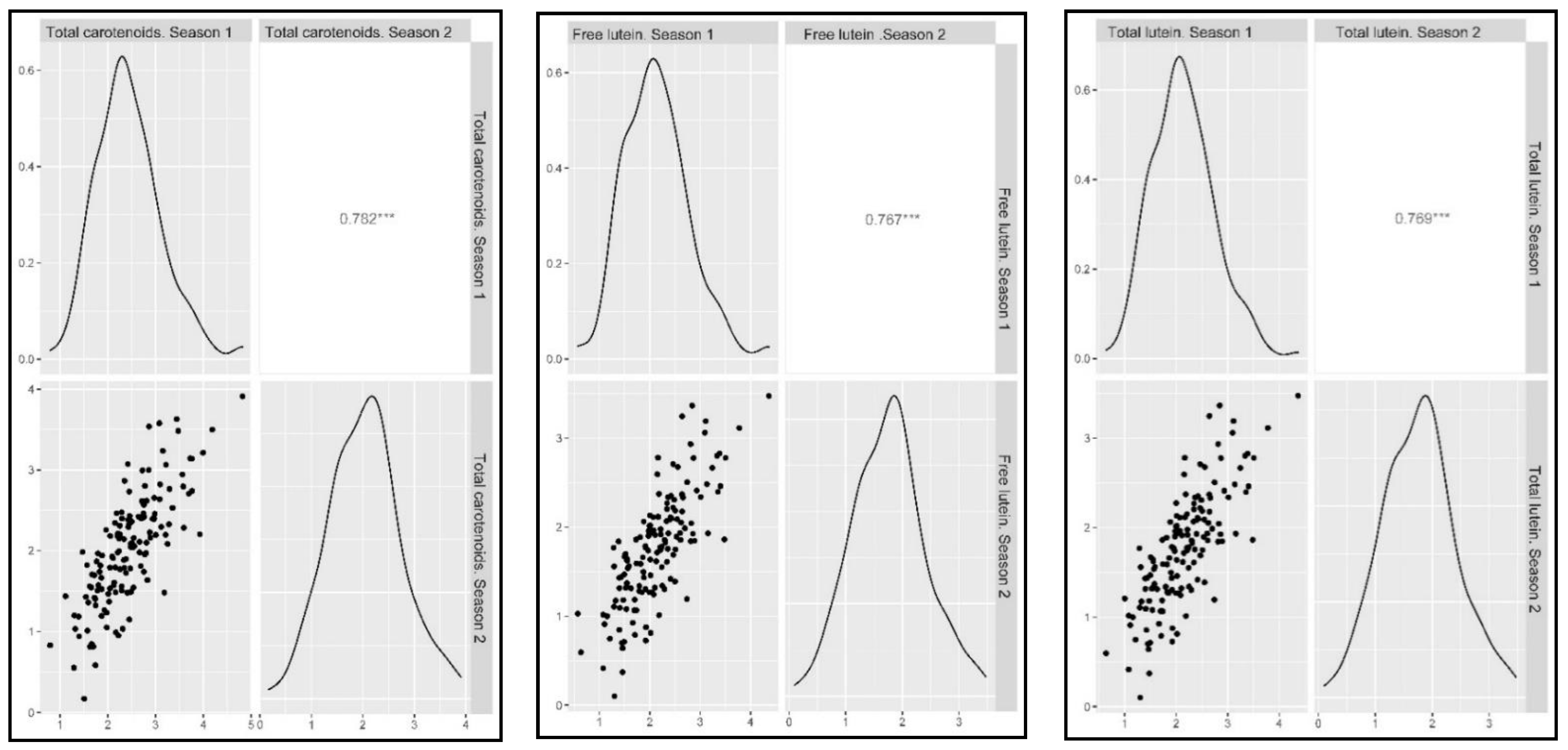
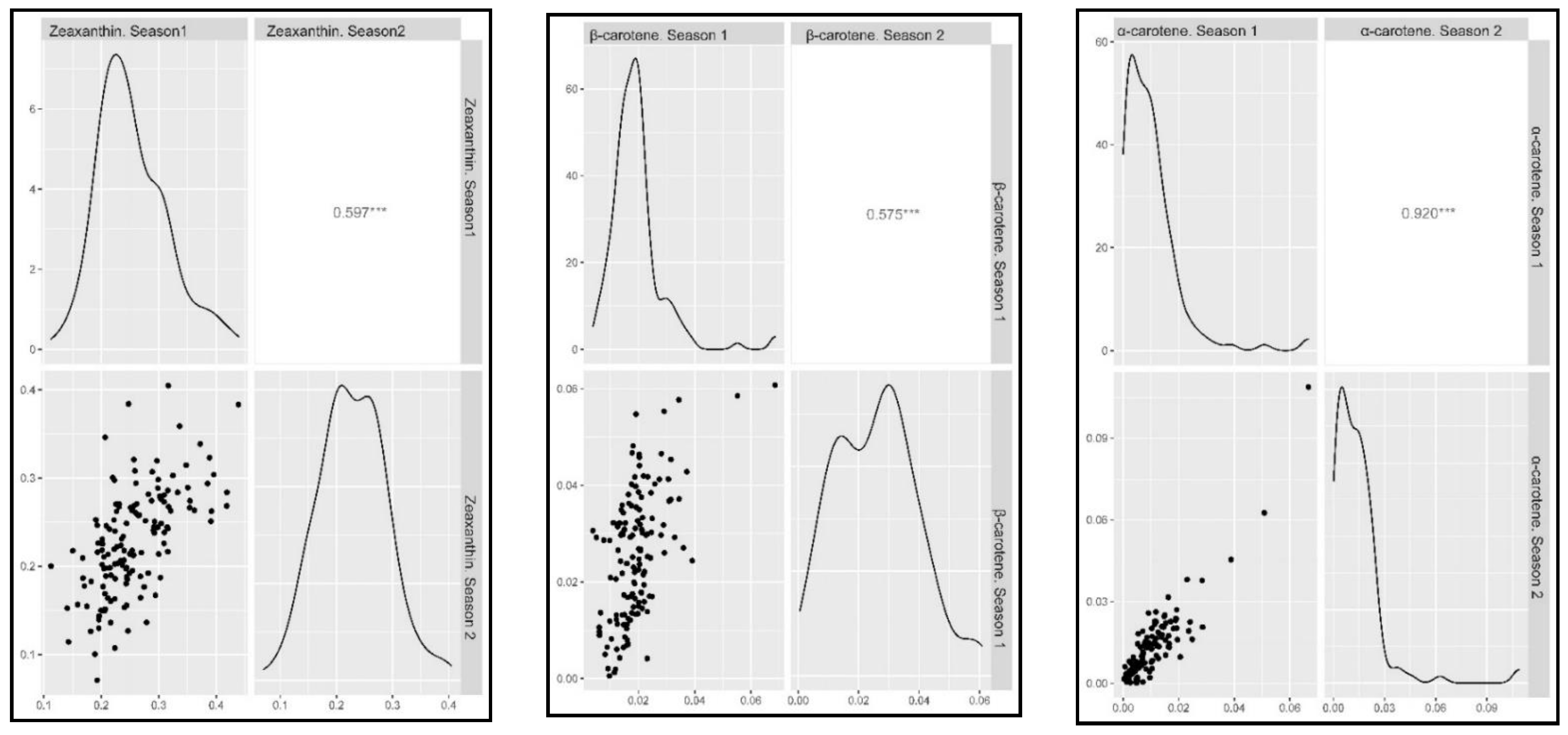
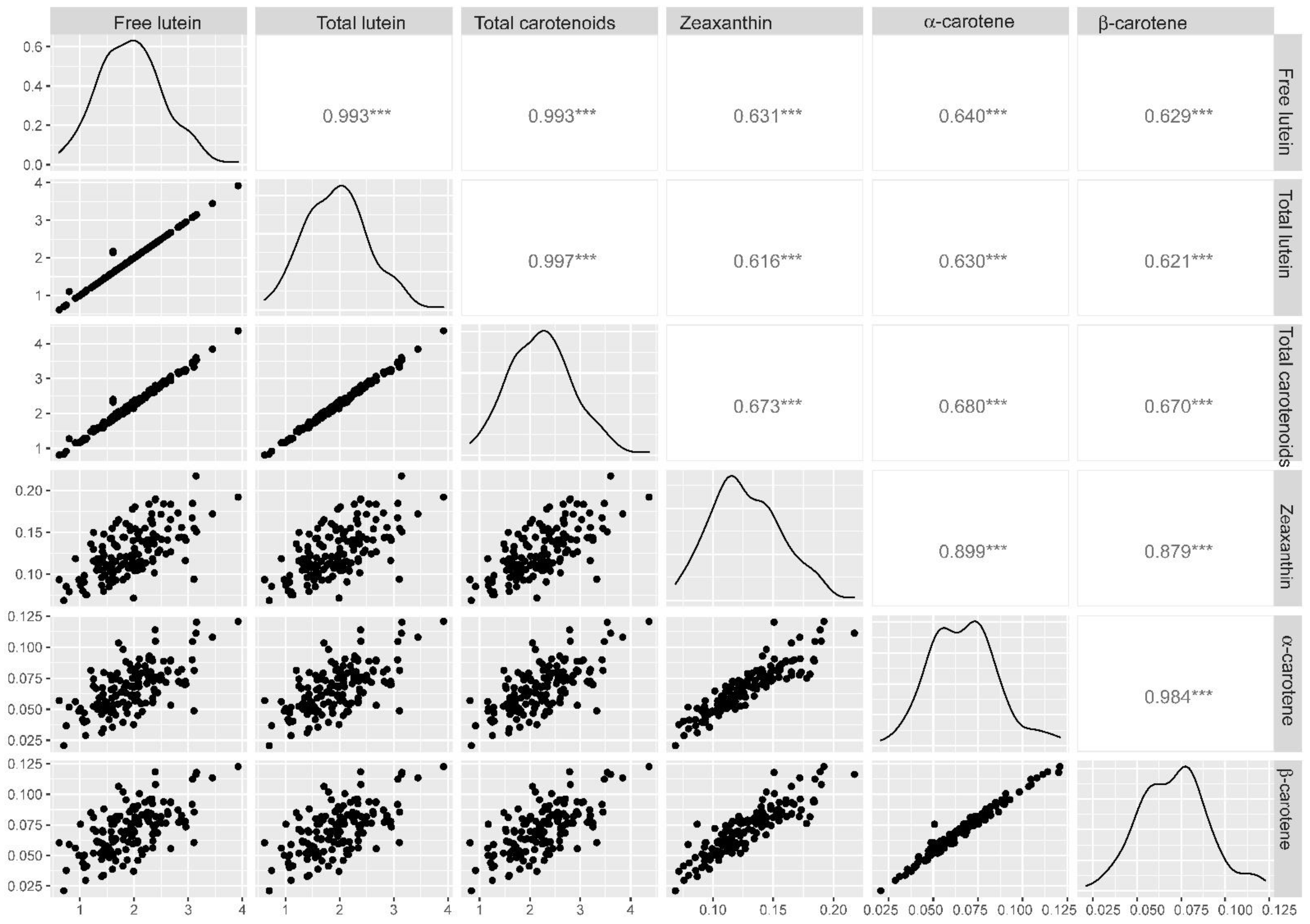
2.2. Phenotypic Assessment
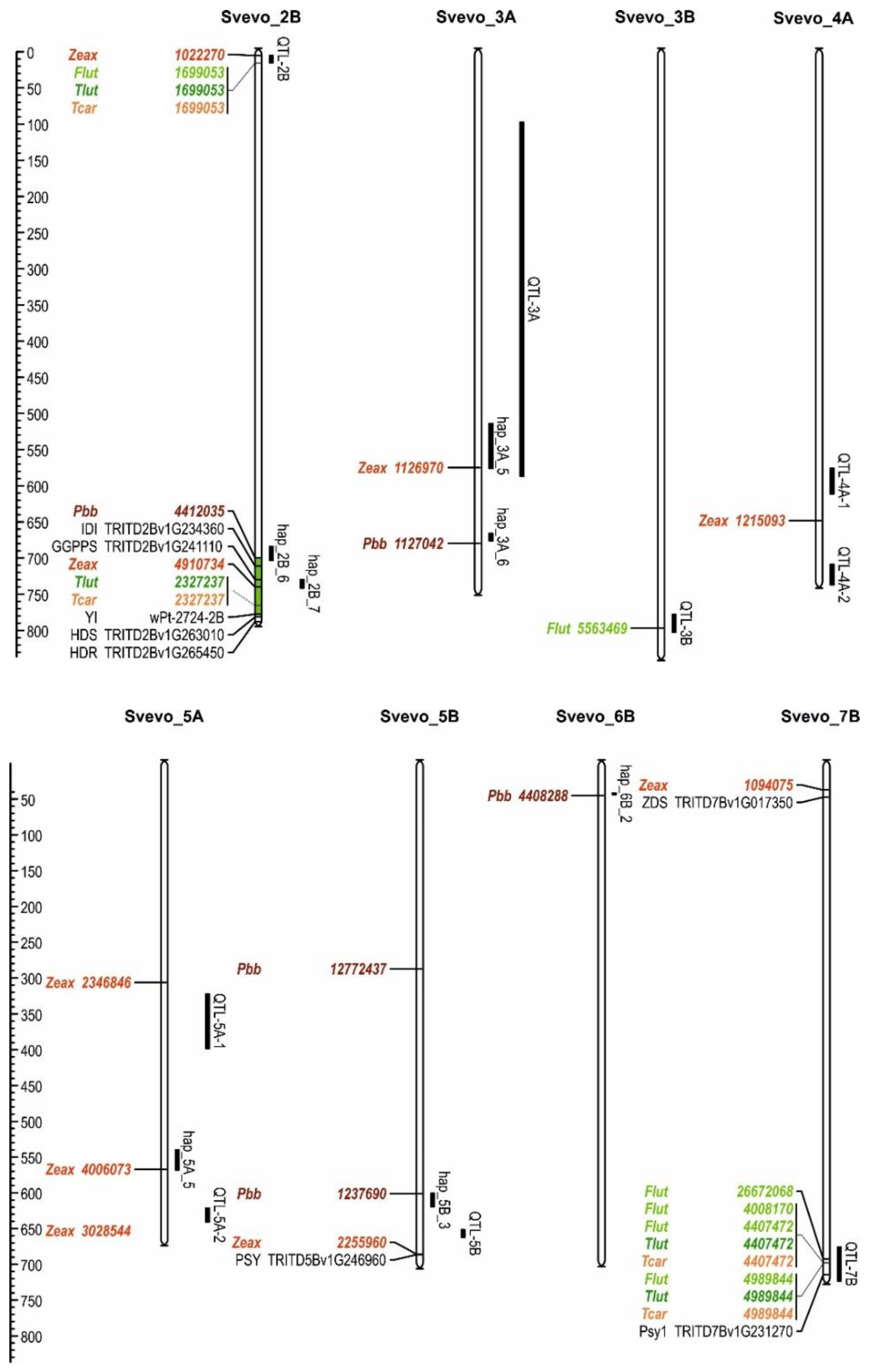
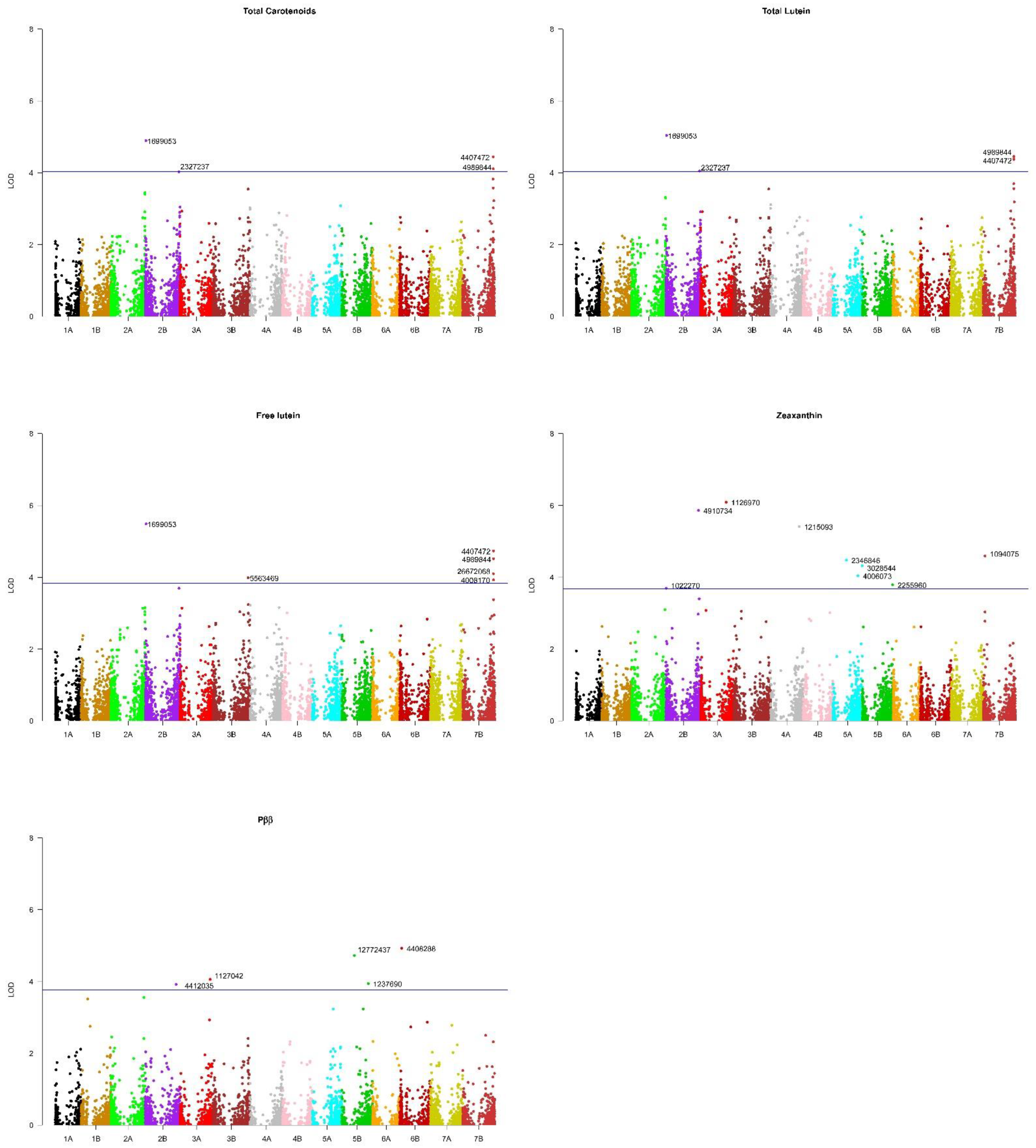
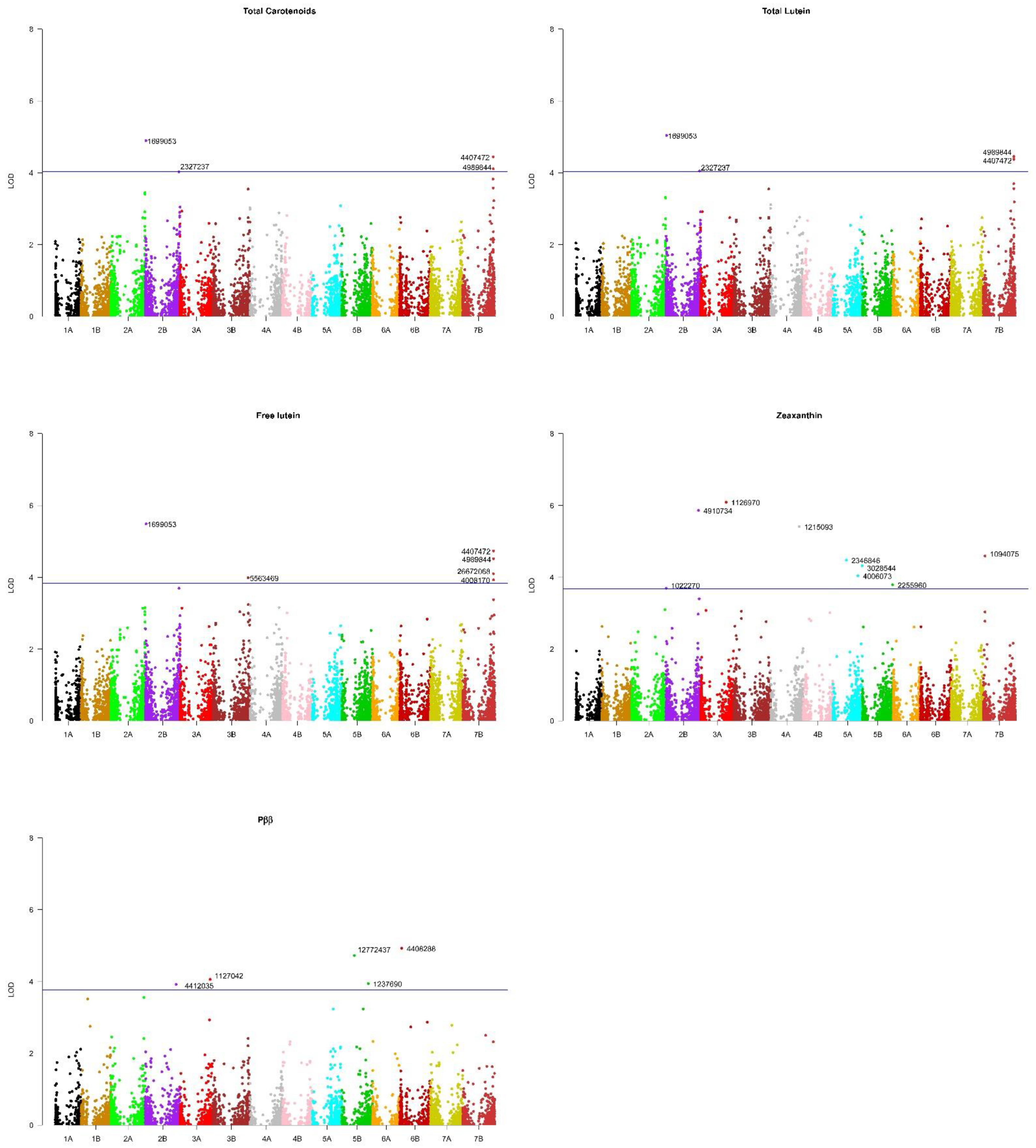
2.3. Marker-Trait Associations
3. Materials and Methods
3.1. Plant Material, Field Design and Statistical Analysis
3.2. Extraction of Carotenoids and HPLC Analysis
3.3. DNA Isolation, Genotyping and Marker-Trait Associations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agriculture and Agri-Food Canada. Canada: Outlook for Principal Field Crops. 1 March 2022. Available online: https://agriculture.canada.ca/en/canadas-agriculture-sectors/crops/canada-outlook-principal-field-crops-march-18-2022 (accessed on 5 July 2022).
- Eurostat Eurostat Database: https://ec.europa.eu/eurostat/web/main/data/database. Available online: https://ec.europa.eu/eurostat (accessed on 5 July 2022).
- Colasuonno, P.; Marcotuli, I.; Blanco, A.; Maccaferri, M.; Condorelli, G.E.; Tuberosa, R.; Parada, R.; de Camargo, A.C.; Schwember, A.R.; Gadaleta, A. Carotenoid pigment content in durum wheat (Triticum turgidum L. var durum): An overview of quantitative trait loci and candidate genes. Front. Plant Sci. 2019, 10, 1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ficco, D.B.M.; Mastrangelo, A.M.; Trono, D.; Borrelli, G.M.; De Vita, P.; Fares, C.; Beleggia, R.; Platani, C.; Papa, R. The colours of durum wheat: A review. Crop Pasture Sci. 2014, 65, 1–15. [Google Scholar] [CrossRef]
- Rodríguez-Suárez, C.; Giménez, M.J.; Atienza, S.G. Progress and perspectives for carotenoid accumulation in selected Triticeae species. Crop Pasture Sci. 2010, 61, 743. [Google Scholar] [CrossRef] [Green Version]
- Blanco, A.; Colasuonno, P.; Gadaleta, A.; Mangini, G.; Schiavulli, A.; Simeone, R.; Digesù, A.M.; De Vita, P.; Mastrangelo, A.M.; Cattivelli, L. Quantitative trait loci for yellow pigment concentration and individual carotenoid compounds in durum wheat. J. Cereal Sci. 2011, 54, 255–264. [Google Scholar] [CrossRef]
- Requena-Ramírez, M.D.; Hornero-Méndez, D.; Rodríguez-Suárez, C.; Atienza, S.G. Durum wheat (Triticum durum L.) landraces reveal potential for the improvement of grain carotenoid esterification in breeding programs. Foods 2021, 10, 757. [Google Scholar] [CrossRef] [PubMed]
- Atienza, S.G.; Ballesteros, J.; Martín, A.; Hornero-Méndez, D. Genetic variability of carotenoid concentration and degree of esterification among tritordeum (x Tritordeum Ascherson et Graebner) and durum wheat accessions. J. Agric. Food Chem. 2007, 55, 4244–4251. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.J. Role of lutein and zeaxanthin in visual and cognitive function throughout the lifespan. Nutr. Rev. 2014, 72, 605–612. [Google Scholar] [CrossRef]
- Nishino, H.; Murakoshi, M.; Tokuda, H.; Satomi, Y. Cancer prevention by carotenoids. Arch. Biochem. Biophys. 2009, 483, 165–168. [Google Scholar] [CrossRef]
- Giuliano, G.; Tavazza, R.; Diretto, G.; Beyer, P.; Taylor, M.A. Metabolic engineering of carotenoid biosynthesis in plants. Trends Biotech. 2008, 26, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Menkir, A.; Palacios-Rojas, N.; Alamu, O.; Dias Paes, M.C.; Dhliwayo, T.; Maziya-dixon, B.; Mengesha, W.; Ndhlela, T.; Oliveira-Guimaraes, P.E.; Pixley, K.; et al. Vitamin A-biofortified Maize: Exploiting Native Genetic Variation for Nutrient Enrichment. Science Brief: Biofortification No. 2 (February 2018). CIMMYT, IITA, EMBRAPA. Available online: https://www.harvestplus.org/wp-content/uploads/2018/02/Vitamin-A-Science-Brief.pdf (accessed on 5 July 2022).
- Zhu, C.; Farré, G.; Zanga, D.; Lloveras, J.; Michelena, A.; Ferrio, J.P.; Voltas, J.; Slafer, G.; Savin, R.; Albajes, R.; et al. High-carotenoid maize: Development of plant biotechnology prototypes for human and animal health and nutrition. Phytochem. Rev. 2018, 17, 195–209. [Google Scholar] [CrossRef]
- Digesù, A.M.; Platani, C.; Cattivelli, L.; Mangini, G.; Blanco, A. Genetic variability in yellow pigment components in cultivated and wild tetraploid wheats. J. Cereal Sci. 2009, 50, 210–218. [Google Scholar] [CrossRef]
- Subira, J.; Peña, R.J.; Álvaro, F.; Ammar, K.; Ramdani, A.; Royo, C. Breeding progress in the pasta-making quality of durum wheat cultivars released in Italy and Spain during the 20th Century. Crop Pasture Sci. 2014, 65, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Bassolino, L.; Petroni, K.; Polito, A.; Marinelli, A.; Azzini, E.; Ferrari, M.; Ficco, D.B.M.; Mazzucotelli, E.; Tondelli, A.; Fricano, A.; et al. Does plant breeding for antioxidant-rich foods have an impact on human health? Antioxidants 2022, 11, 794. [Google Scholar] [CrossRef] [PubMed]
- Parker, G.D.; Chalmers, K.J.; Rathjen, A.J.; Langridge, P. Mapping loci associated with flour colour in wheat (Triticum aestivum L.). Theor. Appl. Genet. 1998, 97, 238–245. [Google Scholar] [CrossRef]
- Elouafi, I.; Nachit, M.M.; Martin, L.M. Identification of a microsatellite on chromosome 7B showing a strong linkage with yellow pigment in durum wheat (Triticum turgidum L. var. durum). Hereditas 2001, 135, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Mares, D.J.; Campbell, A.W. Mapping components of flour colour in Australian wheat. Aust. J. Agric. Res. 2001, 52, 1297–1309. [Google Scholar] [CrossRef] [Green Version]
- Atienza, S.G.; Avila, C.M.; Martín, A. The development of a PCR-based marker for PSY1 from Hordeum chilense, a candidate gene for carotenoid content accumulation in tritordeum seeds. Aust. J. Agric. Res. 2007, 58, 767–773. [Google Scholar] [CrossRef]
- Pozniak, C.J.; Knox, R.E.; Clarke, F.R.; Clarke, J.M. Identification of QTL and association of a phytoene synthase gene with endosperm colour in durum wheat. Theor. Appl. Genet. 2007, 114, 525–537. [Google Scholar] [CrossRef]
- Zhang, W.; Chao, S.; Manthey, F.; Chicaiza, O.; Brevis, J.C.; Echenique, V.; Dubcovsky, J. QTL analysis of pasta quality using a composite microsatellite and SNP map of durum wheat. Theor. Appl. Genet. 2008, 117, 1361–1377. [Google Scholar] [CrossRef]
- Leenhardt, F.; Lyan, B.; Rock, E.; Boussard, A.; Potus, J.; Chanliaud, E.; Remesy, C. Genetic variability of carotenoid concentration, and lipoxygenase and peroxidase activities among cultivated wheat species and bread wheat varieties. Eur. J. Agron. 2006, 25, 170–176. [Google Scholar] [CrossRef]
- Howitt, C.A.; Cavanagh, C.R.; Bowerman, A.F.; Cazzonelli, C.; Rampling, L.; Mimica, J.L.; Pogson, B.J. Alternative splicing, activation of cryptic exons and amino acid substitutions in carotenoid biosynthetic genes are associated with lutein accumulation in wheat endosperm. Funct. Integr. Genom. 2009, 9, 363–376. [Google Scholar] [CrossRef]
- Tanksley, S.D.; McCouch, S.R. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science 1997, 277, 1063–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marone, D.; Russo, M.A.; Mores, A.; Ficco, D.B.M.; Laidò, G.; Mastrangelo, A.M.; Borrelli, G.M. Importance of landraces in cereal breeding for stress tolerance. Plants 2021, 10, 1267. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.L.; Li, M.; McQuinn, R.P.; Chan, K.X.; McFarlane, H.E.; Ermakova, M.; Furbank, R.T.; Mares, D.J.; Dong, C.; Chalmers, K.J.; et al. A GDSL esterase/lipase catalyzes the esterification of lutein in bread wheat. Plant Cell 2019, 31, 3092–3112. [Google Scholar] [CrossRef] [PubMed]
- Watkins, J.; Pogson, B.; Mather, D. XAT Catalyzes Carotenoid Esterification in Wheat. Available online: https://plantae.org/xat-catalyzes-carotenoid-esterification-in-wheat/ (accessed on 5 July 2022).
- Rodríguez-Suárez, C.; Requena-Ramírez, M.D.; Hornero-Méndez, D.; Atienza, S.G. The breeder’s tool-box for enhancing the content of esterified carotenoids in wheat: From extraction and profiling of carotenoids to marker-assisted selection of candidate genes. In Carotenoids: Carotenoid and Apocarotenoid Biosynthesis, Metabolic Engineering and Synthetic Biology; Wurtzel, E.T., Ed.; Elsevier Academic Press: Cambridge, MA, USA, 2022; ISBN 9780323913539. [Google Scholar]
- Ziegler, J.U.; Wahl, S.; Würschum, T.; Longin, C.F.H.; Carle, R.; Schweiggert, R.M. Lutein and lutein esters in whole grain flours made from 75 genotypes of 5 triticum species grown at multiple sites. J. Agric. Food Chem. 2015, 63, 5061–5071. [Google Scholar] [CrossRef] [PubMed]
- Ávila, C.M.; Requena-Ramírez, M.D.; Rodríguez-Suárez, C.; Flores, F.; Sillero, J.C.; Atienza, S.G. Genome-wide association analysis for stem cross section properties, height and heading date in a collection of spanish durum wheat landraces. Plants 2021, 10, 1123. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Hentschel, V.; Kranl, K.; Hollmann, J.; Lindhauer, M.G.; Bohm, V.; Bitsch, R. Spectrophotometric determination of yellow pigment content and evaluation of carotenoids by high-performance liquid chromatography in durum wheat grain. J. Agric. Food Chem. 2002, 50, 6663–6668. [Google Scholar] [CrossRef]
- Abdel-Aal, E.S.M. Identification and quantification of seed carotenoids in selected wheat species. J. Agric. Food Chem. 2007, 55, 787–794. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Maccaferri, M.; Harris, N.S.; Twardziok, S.O.; Pasam, R.K.; Gundlach, H.; Spannagl, M.; Ormanbekova, D.; Lux, T.; Prade, V.M.; Milner, S.G.; et al. Durum wheat genome highlights past domestication signatures and future improvement targets. Nat. Genet. 2019, 51, 885–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiedler, J.D.; Salsman, E.; Liu, Y.; de Jiménez, M.M.; Hegstad, J.B.; Chen, B.; Manthey, F.A.; Chao, S.; Xu, S.; Elias, E.M.; et al. Genome-wide association and prediction of grain and semolina quality traits in durum wheat breeding populations. Plant Genome 2017, 10, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colasuonno, P.; Lozito, M.L.; Marcotuli, I.; Nigro, D.; Giancaspro, A.; Mangini, G.; De Vita, P.; Mastrangelo, A.M.; Pecchioni, N.; Houston, K.; et al. The carotenoid biosynthetic and catabolic genes in wheat and their association with yellow pigments. BMC Genom. 2017, 18, 122. [Google Scholar] [CrossRef] [Green Version]
- Colasuonno, P.; Gadaleta, A.; Giancaspro, A.; Nigro, D.; Giove, S.; Incerti, O.; Mangini, G.; Signorile, A.; Simeone, R.; Blanco, A. Development of a high-density SNP-based linkage map and detection of yellow pigment content QTLs in durum wheat. Mol. Breed. 2014, 34, 1563–1578. [Google Scholar] [CrossRef]
- Roncallo, P.F.; Cervigni, G.L.; Jensen, C.; Miranda, R.; Carrera, A.D.; Helguera, M.; Echenique, V. QTL analysis of main and epistatic effects for flour color traits in durum wheat. Euphytica 2012, 185, 77–92. [Google Scholar] [CrossRef] [Green Version]
- Giraldo, P.; Royo, C.; González, M.; Carrillo, J.M.; Ruiz, M. Genetic diversity and association mapping for agromorphological and grain quality traits of a structured collection of durum wheat landraces including subsp. durum, turgidum and diccocon. PLoS ONE 2016, 11, e0166577. [Google Scholar] [CrossRef] [Green Version]
- Reimer, S.; Pozniak, C.J.; Clarke, F.R.; Clarke, J.M.; Somers, D.J.; Knox, R.E.; Singh, A.K. Association mapping of yellow pigment in an elite collection of durum wheat cultivars and breeding lines. Genome 2008, 51, 1016–1025. [Google Scholar] [CrossRef]
- N’Diaye, A.; Haile, J.K.; Nilsen, K.T.; Walkowiak, S.; Ruan, Y.; Singh, A.K.; Clarke, F.R.; Clarke, J.M.; Pozniak, C.J. Haplotype loci under selection in canadian durum wheat germplasm over 60 years of breeding: Association with grain yield, quality traits, protein loss, and plant height. Front. Plant Sci. 2018, 9, 1589. [Google Scholar] [CrossRef]
- Roselló, M.; Royo, C.; Álvaro, F.; Villegas, D.; Nazco, R.; Soriano, J.M. Pasta-making quality QTLome from mediterranean durum wheat landraces. Front. Plant Sci. 2018, 9, 1512. [Google Scholar] [CrossRef]
- Gallagher, C.E.; Matthews, P.D.; Li, F.; Wurtzel, E.T. Gene duplication in the carotenoid biosynthetic pathway preceded evolution of the grasses. Plant Physiol. 2004, 135, 1776–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.Q.; Vallabhaneni, R.; Yu, J.; Rocheford, T.; Wurtzel, E.T. The maize phytoene synthase gene family: Overlapping roles for carotenogenesis in endosperm, photomorphogenesis, and thermal stress tolerance. Plant Physiol. 2008, 147, 1334–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsch, R.; Wust, F.; Bar, C.; Al-Babili, S.; Beyer, P. A third phytoene synthase is devoted to abiotic stress-induced abscisic acid formation in rice and defines functional diversification of phytoene synthase genes. Plant Physiol. 2008, 147, 367–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Suárez, C.; Mellado-Ortega, E.; Hornero-Méndez, D.; Atienza, S.G. Increase in transcript accumulation of Psy1 and e-Lcy genes in grain development is associated with differences in seed carotenoid content between durum wheat and tritordeum. Plant Mol. Biol. 2014, 84, 659–673. [Google Scholar] [CrossRef] [Green Version]
- Cong, L.; Wang, C.; Li, Z.; Chen, L.; Yang, G.; Wang, Y.; He, G. cDNA cloning and expression analysis of wheat (Triticum aestivum L.) phytoene and ζ-carotene desaturase genes. Mol. Biol. Rep. 2010, 37, 3351–3361. [Google Scholar] [CrossRef]
- Zhang, C.; Dong, C.H.; He, X.; Zhang, L.; Xia, X.C.; He, Z.H. Allelic variants at the TaZds-D1 locus on wheat chromosome 2DL and their association with yellow pigment content. Crop Sci. 2011, 51, 1580–1590. [Google Scholar] [CrossRef]
- Dong, C.; Ma, Z.; Xia, X.; Zhang, L.; He, Z. Allelic variation at the TaZds-A1 locus on wheat chromosome 2A and development of a functional marker in common wheat. J. Integr. Agric. 2012, 11, 1067–1074. [Google Scholar] [CrossRef]
- Pasten, M.C.; Roncallo, P.F.; Camargo Acosta, E.Y.; Echenique, V.; Garbus, I. Association of novel characterized sequence variations in the ζ-carotene desaturase (Zds) gene with yellow color and yellow pigment content in durum wheat cultivars. J. Cereal Sci. 2021, 99, 103185. [Google Scholar] [CrossRef]
- Atienza, S.G.; Ramirez, C.M.; Hernandez, P.; Martin, A. Chromosomal location of genes for carotenoid pigments in Hordeum chilense. Plant Breed. 2004, 123, 303–304. [Google Scholar] [CrossRef]
- Rodríguez-Suárez, C.; Atienza, S.G.G. Hordeum chilense genome, a useful tool to investigate the endosperm yellow pigment content in the Triticeae. BMC Plant Biol. 2012, 12, 200. [Google Scholar] [CrossRef] [Green Version]
- Avila, C.M.; Mattera, M.G.; Rodríguez-Suárez, C.; Palomino, C.; Ramírez, M.C.; Martin, A.; Kilian, A.; Hornero-Méndez, D.; Atienza, S.G. Diversification of seed carotenoid content and profile in wild barley (Hordeum chilense Roem. et Schultz.) and Hordeum vulgare L.–H. chilense synteny as revealed by DArTSeq markers. Euphytica 2019, 215, 45. [Google Scholar] [CrossRef]
- Matthews, P.D.; Wurtzel, E.T. Metabolic engineering of carotenoid accumulation in Escherichia coli by modulation of the isoprenoid precursor pool with expression of deoxyxylulose phosphate synthase. Appl. Microbiol. Biotechnol. 2000, 53, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Sharkey, T.D. Methylerythritol 4-phosphate (MEP) pathway metabolic regulation. Nat. Prod. Rep. 2014, 31, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallabhaneni, R.; Wurtzel, E.T. Timing and biosynthetic potential for carotenoid accumulation in genetically diverse germplasm of maize. Plant Physiol. 2009, 150, 562–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leenhardt, F.; Lyan, B.; Rock, E.; Boussard, A.; Potus, J.; Chanliaud, E.; Remesy, C. Wheat lipoxygenase activity induces greater loss of carotenoids than vitamin E during breadmaking. J. Agric. Food Chem. 2006, 54, 1710–1715. [Google Scholar] [CrossRef] [PubMed]
- Fraignier, M.P.; Michaux-Ferrière, N.; Kobrehel, K. Distribution of peroxidases in durum wheat (Triticum durum). Cereal Chem. 2000, 77, 11–17. [Google Scholar] [CrossRef]
- Ruiz, M.; Giraldo, P.; Royo, C.; Carrillo, J.M. Creation and validation of the spanish durum wheat core collection. Crop Sci. 2013, 53, 2530–2537. [Google Scholar] [CrossRef] [Green Version]
- Aravind, J.; Mukesh Sankar, S.; Wankhede, D.P.; Kaur, V. AugmentedRCBD: Analysis of Augmented Randomised Complete Block Designs. R package Version 0.1. Available online: https://aravind-j.github.io/augmentedRCBD/ (accessed on 12 July 2022).
- Federer, W.T. Augmented designs. Hawaii. Plant. Rec. 1956, 55, 191–208. [Google Scholar] [CrossRef] [Green Version]
- Federer, W. Augmented designs with one-way elimination of heterogeneity. Biometrics 1961, 17, 447–473. [Google Scholar] [CrossRef]
- Piepho, H.P.; Möhring, J. Computing heritability and selection response from unbalanced plant breeding trials. Genetics 2007, 177, 1881–1888. [Google Scholar] [CrossRef] [Green Version]
- Mínguez-Mosquera, M.I.; Hornero-Méndez, D. Separation and quantification of the carotenoid pigments in red peppers (Capsicum annuum L.), paprika and oleoresin by reversed-phase HPLC. J. Agric. Food Chem. 1993, 41, 1616–1620. [Google Scholar] [CrossRef]
- Mellado-Ortega, E.; Hornero-Méndez, D. Carotenoid evolution during short-storage period of durum wheat (Triticum turgidum conv. durum) and tritordeum (x Tritordeum Ascherson et Graebner) whole-grain flours. Food Chem. 2016, 192, 714–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, s0022–s2836. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Taylor and Francis: Oxfordshire, UK, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Hamazaki, K.; Iwata, H. Rainbow: Haplotype-based genome-wide association study using a novel SNP-set method. PLoS Comput. Biol. 2020, 16, e1007663. [Google Scholar] [CrossRef] [Green Version]
- R_Studio_Team RStudio: Integrated Development for R. RStudio, PBC, Boston, MA. Available online: https://support.rstudio.com/hc/en-us/articles/206212048-Citing-RStudio (accessed on 5 July 2022).
- Turner, S. qqman: An R package for visualizing GWAS results using Q-Q and manhattan plots. J. Open Source Softw. 2018, 3, 731. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Trait 1 | Chromosome | Pos (Mbp) 2 | LOD | FDR 3 | R-square | Type 4 | Effect 5 | Svevo 6 |
|---|---|---|---|---|---|---|---|---|---|
| 1022270 | Zeax | 2B | 5.33 | 3.70 | 0.200 | 0.096 | SNP/T|C | 0.01 | C |
| 1699053 | Flut | 2B | 15.6 | 5.49 | 0.028 | 0.083 | PAV/T|G | 0.49 | T |
| 1699053 | Tlut | 2B | 15.6 | 5.04 | 0.073 | 0.076 | PAV/T|G | 0.47 | T |
| 1699053 | Tcar | 2B | 15.6 | 4.90 | 0.102 | 0.075 | PAV/T|G | 0.51 | T |
| 4412035 | Pββ | 2B | 699.6 | 3.93 | 0.155 | 0.111 | SNP/A|G | 3.09 | A |
| 4910734 | Zeax | 2B | 739.9 | 5.86 | 0.006 | 0.149 | SNP/T|C | 0.01 | T |
| 2327237 | Tlut | 2B | 765.6 | 4.05 | 0.195 | 0.065 | PAV/T|G | 0.47 | T |
| 2327237 | Tcar | 2B | 765.6 | 4.03 | 0.201 | 0.065 | PAV/T|G | 0.06 | T |
| 1126970 | Zeax | 3A | 575.0 | 6.09 | 0.006 | 0.144 | SNP/T|C | 0.02 | T |
| 1127042 | Pββ | 3A | 679.7 | 4.06 | 0.130 | 0.133 | SNP/T|C | 1.61 | C |
| 5563469 | Flut | 3B | 796.9 | 3.99 | 0.173 | 0.060 | PAV/T|G | 0.65 | T |
| 1215093 | Zeax | 4A | 648.0 | 5.42 | 0.011 | 0.152 | SNP/T|A | 0.01 | A |
| 2346846 | Zeax | 5A | 306.3 | 4.48 | 0.060 | 0.124 | SNP/C|T | 0.10 | C |
| 4006073 | Zeax | 5A | 567.0 | 4.04 | 0.110 | 0.102 | SNP/A|G | 0.003 | A |
| 3028544 | Zeax | 5A | 661.4 | 4.32 | 0.070 | 0.110 | SNP/T|C | 0.02 | T |
| 12772437 | Pββ | 5B | 287.2 | 4.73 | 0.051 | 0.137 | SNP/T|C | 3.56 | T |
| 1237690 | Pββ | 5B | 601.1 | 3.95 | 0.154 | 0.108 | SNP/T|C | 4.17 | C |
| 2255960 | Zeax | 5B | 685.8 | 3.79 | 0.160 | 0.091 | SNP/G|A | 0.01 | A |
| 4408288 | Pββ | 6B | 45.2 | 4.93 | 0.051 | 0.146 | SNP/C|G | 0.69 | C |
| 1094075 | Zeax | 7B | 37.4 | 4.59 | 0.050 | 0.078 | SNP/A|G | 0.01 | A |
| 26672068 | Flut | 7B | 692.4 | 4.10 | 0.155 | 0.055 | PAV/T|G | 0.54 | T |
| 4008170 | Flut | 7B | 697.8 | 3.93 | 0.179 | 0.040 | PAV/T|G | 0.49 | T |
| 4407472 | Flut | 7B | 697.8 | 4.74 | 0.070 | 0.044 | PAV/T|G | 0.54 | T |
| 4407472 | Tlut | 7B | 697.8 | 4.37 | 0.130 | 0.050 | PAV/T|G | 0.53 | T |
| 4407472 | Tcar | 7B | 697.8 | 4.44 | 0.145 | 0.042 | PAV/T|G | 0.56 | T |
| 4989844 | Flut | 7B | 698.6 | 4.52 | 0.095 | 0.042 | PAV/T|G | 0.67 | T |
| 4989844 | Tlut | 7B | 698.6 | 4.45 | 0.121 | 0.0430 | PAV/T|G | 0.67 | T |
| 4989844 | Tcar | 7B | 698.6 | 4.11 | 0.190 | 0.040 | PAV/T|G | 0.69 | T |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Requena-Ramírez, M.D.; Rodríguez-Suárez, C.; Flores, F.; Hornero-Méndez, D.; Atienza, S.G. Marker-Trait Associations for Total Carotenoid Content and Individual Carotenoids in Durum Wheat Identified by Genome-Wide Association Analysis. Plants 2022, 11, 2065. https://doi.org/10.3390/plants11152065
Requena-Ramírez MD, Rodríguez-Suárez C, Flores F, Hornero-Méndez D, Atienza SG. Marker-Trait Associations for Total Carotenoid Content and Individual Carotenoids in Durum Wheat Identified by Genome-Wide Association Analysis. Plants. 2022; 11(15):2065. https://doi.org/10.3390/plants11152065
Chicago/Turabian StyleRequena-Ramírez, María Dolores, Cristina Rodríguez-Suárez, Fernando Flores, Dámaso Hornero-Méndez, and Sergio G. Atienza. 2022. "Marker-Trait Associations for Total Carotenoid Content and Individual Carotenoids in Durum Wheat Identified by Genome-Wide Association Analysis" Plants 11, no. 15: 2065. https://doi.org/10.3390/plants11152065
APA StyleRequena-Ramírez, M. D., Rodríguez-Suárez, C., Flores, F., Hornero-Méndez, D., & Atienza, S. G. (2022). Marker-Trait Associations for Total Carotenoid Content and Individual Carotenoids in Durum Wheat Identified by Genome-Wide Association Analysis. Plants, 11(15), 2065. https://doi.org/10.3390/plants11152065