
Short-Term CO2 Treatment of Harvested Grapes (Vitis vinifera L., cv. Trebbiano) before Partial Dehydration Affects Berry Secondary Metabolism and the Aromatic Profile of the Resulting Wine

 , ,
, , 
Abstract
:1. Introduction
2. Results
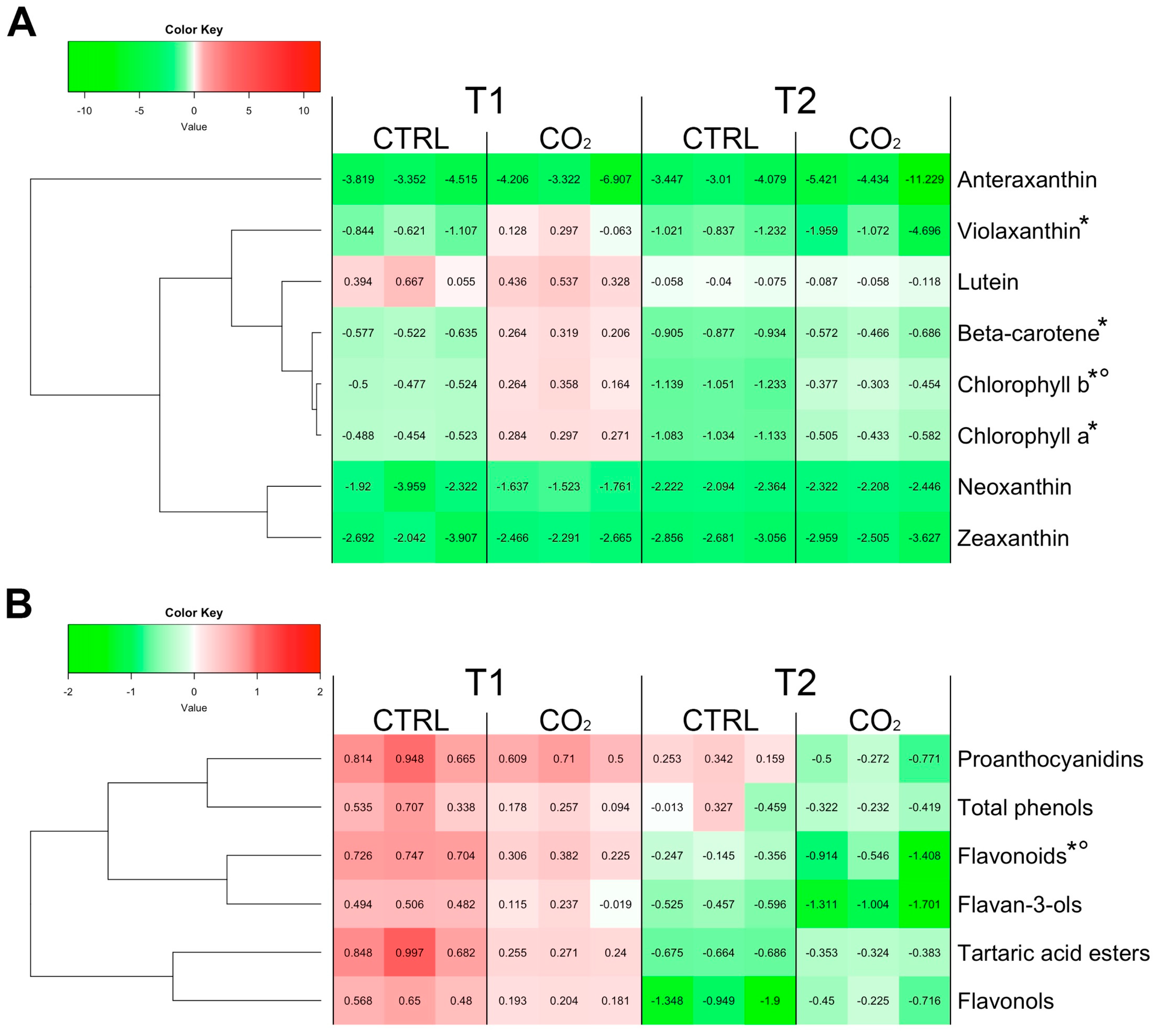
2.1. Lipophilic and Phenolic Compounds in Berry Skins
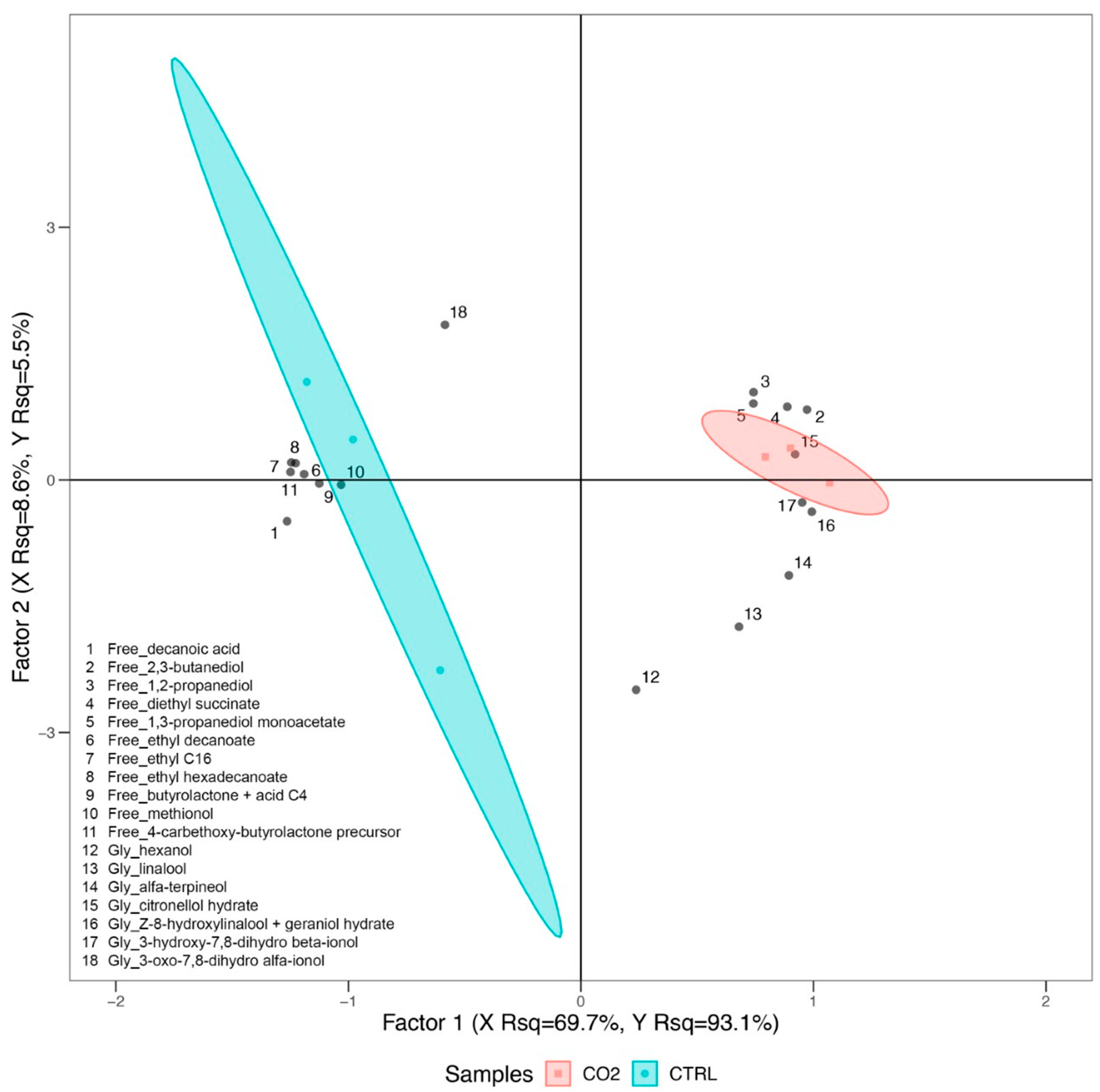
2.2. Free and Glycosylated Aroma Compounds in Trebbiano Wines
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental Plan
5.2. Berry Lipophilic Compounds Extraction and HPLC Detection and Quantification
5.3. Berry Polyphenol Extraction and Quantification
5.4. Aroma Compounds Extraction and Analysis of the Wines
5.4.1. Total Glycosylated Aroma Compounds Extraction
5.4.2. Enzymatic Hydrolysis of the Total Glycosylated Aroma Compounds
5.4.3. Total Free Aroma Compounds Extraction
5.4.4. GC-MS Analysis
5.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brizzolara, S.; Manganaris, G.A.; Fotopoulos, V.; Watkins, C.B.; Tonutti, P. Primary Metabolism in Fresh Fruits During Storage. Front. Plant Sci. 2020, 11, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanellis, A.; Tonutti, P.; Perata, P. Biochemical and molecular aspects of modified and controlled atmospheres. In Modified and Controlled Atmosphere for the Storage, Transportation, and Packaging of Horticultural Commodities; Yahia, E.M., Ed.; CRC Press: Boca Raton, FL, USA, 2009; pp. 553–567. [Google Scholar]
- Cukrov, D.; Zermiani, M.; Brizzolara, S.; Cestaro, A.; Licausi, F.; Luchinat, C.; Santucci, C.; Tenori, L.; van Veen, H.; Zuccolo, A.; et al. Extreme Hypoxic Conditions Induce Selective Molecular Responses and Metabolic Reset in Detached Apple Fruit. Front. Plant Sci. 2016, 7, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Q.G.; Gong, Z.Y.; Wang, M.M.; Li, X.; Grierson, D.; Yin, X.R.; Chen, K.S. A Transcription Factor Network Responsive to High CO2/Hypoxia Is Involved in Deastringency in Persimmon Fruit. J. Exp. Bot. 2018, 69, 2061–2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandra, D.; Lee, J.S.; Hong, Y.P.; Park, M.H.; Choi, A.J.; Kim, J.G. Short-Term Application of CO 2 Gas: Effects on Physicochemical, Microbial, and Sensory Qualities of “Charlotte” Strawberry during Storage. J. Food Saf. 2019, 39, e12597. [Google Scholar] [CrossRef]
- Ahumada, M.H.; Mitcham, E.J.; Moore, D.G. Postharvest Quality of “Thompson Seedless” Grapes after Insecticidal Controlled-Atmosphere Treatments. Hortscience 1996, 31, 833–836. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Ballesta, M.T.; Jiménez, J.B.; Romero, I.; Orea, J.M.; Maldonado, R.; Ureña, Á.G.; Escribano, M.I.; Merodio, C. Effect of High CO2 Pretreatment on Quality, Fungal Decay and Molecular Regulation of Stilbene Phytoalexin Biosynthesis in Stored Table Grapes. Postharvest Biol. Technol. 2006, 42, 209–216. [Google Scholar] [CrossRef]
- Sanchez-Ballesta, M.T.; Romero, I.; Jiménez, J.B.; Orea, J.M.; González-Ureña, Á.; Escribano, M.I.; Merodio, C. Involvement of the Phenylpropanoid Pathway in the Response of Table Grapes to Low Temperature and High CO2 Levels. Postharvest Biol. Technol. 2007, 46, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Becatti, E.; Chkaiban, L.; Tonutti, P.; Forcato, C.; Bonghi, C.; Ranieri, A.M. Short-Term Postharvest Carbon Dioxide Treatments Induce Selective Molecular and Metabolic Changes in Grape Berries. J. Agric. Food Chem. 2010, 58, 8012–8020. [Google Scholar] [CrossRef]
- Sanchez-Ballesta, M.T.; Alvarez, I.; Escribano, M.I.; Merodio, C.; Romero, I. Effect of High CO2 Levels and Low Temperature on Stilbene Biosynthesis Pathway Gene Expression and Stilbenes Production in White, Red and Black Table Grape Cultivars during Postharvest Storage. Plant Physiol. Biochem. 2020, 151, 334–341. [Google Scholar] [CrossRef]
- Besada, C.; Sanchez, G.; Salvador, A.; Granell, A. Volatile Compounds Associated to the Loss of Astringency in Persimmon Fruit Revealed by Untargeted GC-MS Analysis. Metabolomics 2013, 9, 157–172. [Google Scholar] [CrossRef]
- Amorim, C.; Alves Filho, E.G.; Rodrigues, T.H.S.; Bender, R.J.; Canuto, K.M.; Garruti, D.S.; Antoniolli, L.R. Volatile Compounds Associated to the Loss of Astringency in ‘Rama Forte’ Persimmon Fruit. Food Res. Int. 2020, 136, 109570. [Google Scholar] [CrossRef]
- Mencarelli, F.; Tonutti, P. Sweet, Reinforced and Fortified Wines: Grape Biochemistry, Technology and Vinification; John Wiley & Sons: Hoboken, NJ, USA; Chichester, West Sussex, UK, 2013; p. 357. [Google Scholar]
- Zamboni, A.; Minoia, L.; Ferrarini, A.; Tornielli, G.B.; Zago, E.; Delledonne, M.; Pezzotti, M. Molecular Analysis of Post-Harvest Withering in Grape by AFLP Transcriptional Profiling. J. Exp. Bot. 2008, 59, 4145–4159. [Google Scholar] [CrossRef] [Green Version]
- Rizzini, F.M.; Bonghi, C.; Tonutti, P. Postharvest Water Loss Induces Marked Changes in Transcript Profiling in Skins of Wine Grape Berries. Postharvest Biol. Technol. 2009, 52, 247–253. [Google Scholar] [CrossRef]
- Costantini, V.; Bellincontro, A.; de Santis, D.; Botondi, R.; Mencarelli, F. Metabolic Changes of Malvasia Grapes for Wine Production during Postharvest Drying. J. Agric. Food Chem. 2006, 54, 3334–3340. [Google Scholar] [CrossRef]
- Chkaiban, L.; Botondi, R.; Bellincontro, A.; de Santis, D.; Kefalas, P.; Mencarelli, F. Influence of Postharvest Water Stress on Lipoxygenase Andalcohol Dehydrogenase Activities, and on the Composition of Some Volatile Compounds of Gewürztraminer Grapes Dehydrated under Controlled and Uncontrolled Thermohygrometric Conditions. Aust. J. Grape Wine Res. 2007, 13, 142–149. [Google Scholar] [CrossRef]
- Bellincontro, A.; de Santis, D.; Botondi, R.; Villa, I.; Mencarelli, F. Different Postharvest Dehydration Rates Affect Quality Characteristics and Volatile Compounds of Malvasia, Trebbiano and Sangiovese Grapes for Wine Production. J. Sci. Food Agric. 2004, 84, 1791–1800. [Google Scholar] [CrossRef]
- Moreno, J.J.; Cerpa-Calderón, F.; Cohen, S.D.; Fang, Y.; Qian, M.; Kennedy, J.A. Effect of postharvest dehydration on the composition of pinot noir grapes (Vitis vinifera L.) and wine. Food Chem. 2008, 109, 755–762. [Google Scholar] [CrossRef]
- Mencarelli, F.; Bellincontro, A. Recent Advances in Postharvest Technology of the Wine Grape to Improve the Wine Aroma. J. Sci. Food Agric. 2020, 100, 5046–5055. [Google Scholar] [CrossRef]
- Vazquez-Hernandez, M.; Romero, I.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Deciphering the Role of CBF/DREB Transcription Factors and Dehydrins in Maintaining the Quality of Table Grapes Cv. Autumn Royal Treated with High CO2levels and Stored at 0 °C. Front. Plant Sci. 2017, 8, 1591. [Google Scholar] [CrossRef]
- Romero, I.; Domínguez, I.; Doménech-Carbó, A.; Gavara, R.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Effect of High Levels of CO2on the Electrochemical Behavior and the Enzymatic and Non-Enzymatic Antioxidant Systems in Black and White Table Grapes Stored at 0 °C. J. Sci. Food Agric. 2019, 99, 6859–6867. [Google Scholar] [CrossRef]
- Rosales, R.; Romero, I.; Fernandez-Caballero, C.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. Low temperature and short-term high-CO2 treatment in postharvest storage of table grapes at two maturity stages: Effects on transcriptome profiling. Front. Plant Sci. 2016, 7, 1020. [Google Scholar] [CrossRef] [Green Version]
- Blanch, M.; Sanchez-Ballesta, M.T.; Escribano, M.I.; Merodio, C. Water Distribution and Ionic Balance in Response to High CO2 Treatments in Strawberries (Fragaria vesca L. Cv. Mara de Bois). Postharvest Biol. Technol. 2012, 73, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Rocchi, L.; Rustioni, L.; Failla, O. Chlorophyll and Carotenoid Quantifications in White Grape (Vitis vinifera L.) Skins by Reflectance Spectroscopy. Vitis-J. Grapevine Res. 2016, 55, 11–16. [Google Scholar] [CrossRef]
- Huang, X.; Zhang, X.; Jiang, X.; Huang, S.; Pang, X.; Qu, H.; Zhang, Z. Quality Retention and Selective Gene Expression of Chinese Flowering Cabbage as Affected by Atmosphere Gas Composition. J. Food Processing Preserv. 2020, 44, e14548. [Google Scholar] [CrossRef]
- Gonçalves, B.; Falco, V.; Moutinho-Pereira, J.; Bacelar, E.; Peixoto, F.; Correia, C. Effects of Elevated CO2 on Grapevine (Vitis vinifera L.): Volatile Composition, Phenolic Content, and in Vitro Antioxidant Activity of Red Wine. J. Agric. Food Chem. 2009, 57, 265–273. [Google Scholar] [CrossRef] [PubMed]
- González-Lázaro, M.; Martínez-Lapuente, L.; Guadalupe, Z.; Ayestaran, B.; Bueno-Herrera, M.; López de la Cuesta, P.; Pérez-Magariño, S. Evaluation of Grape Ripeness, Carbonic Maceration and Pectolytic Enzymes to Improve the Chemical and Sensory Quality of Red Sparkling Wines. J. Sci. Food Agric. 2020, 100, 2618–2629. [Google Scholar] [CrossRef] [PubMed]
- Ayestarán, B.; Martínez-Lapuente, L.; Guadalupe, Z.; Canals, C.; Adell, E.; Vilanova, M. Effect of the Winemaking Process on the Volatile Composition and Aromatic Profile of Tempranillo Blanco Wines. Food Chem. 2019, 276, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Dourtoglou, V.G.; Yannovits, N.G.; Tychopoulos, V.G.; Vamvakias, M.M. Effect of Storage under CO2 Atmosphere on the Volatile, Amino Acid, and Pigment Constituents in Red Grape (Vitis vinifera L. Var. Agiorgitiko). J. Agric. Food Chem. 1994, 42, 338–344. [Google Scholar] [CrossRef]
- Yang, D.Y.; Kakuda, Y.; Subden, R.E. Higher Alcohols, Diacetyl, Acetoin and 2,3-Butanediol Biosynthesis in Grapes Undergoing Carbonic Maceration. Food Res. Int. 2006, 39, 112–116. [Google Scholar] [CrossRef]
- Castagna, A.; Nali, C.; Ciompi, S.; Lorenzini, G.; Soldatini, G.F.; Ranieri, A.; Ranieri, A. Ozone Exposure Affects Photosynthesis of Pumpkin (Cucurbita Pepo) Plants. New Phytol. 2001, 152, 223–229. [Google Scholar] [CrossRef]
- Borbalan, A.M.A.; Zorro, L.; Guillen, D.A.; Barroso, C.G. Study of the polyphenol content of red and white grape varieties by liquid chromatography–mass spectrometry and its relationship to antioxidant power. J. Chromatogr. 2003, 1012, 31–38. [Google Scholar] [CrossRef]
- Kim, D.O.; Chun, O.K.; Kim, Y.J.; Moon, H.Y.; Lee, C.Y. Quantification of Polyphenolics and Their Antioxidant Capacity in Fresh Plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef]
- Romani, A.; Mancini, P.; Tatti, S.; Vincieri, F.F. Polyphenols and polysaccharides in Tuscan grapes and wines. Ital. J. Food Sci. 1996, 1, 13–24. [Google Scholar]
- Nagel, C.W.; Glories, Y. Use of a modified dimethylaminocinnamaldehyde reagent for analysis of flavanols. Am. J. Enol. Vitic. 1991, 42, 364–366. [Google Scholar]
- Aubert, C.; Ambid, C.; Baumes, R.; Günata, Z. Investigation of Bound Aroma Constituents of Yellow-Fleshed Nectarines (Prunus Persica L. Cv. Springbright). Changes in Bound Aroma Profile during Maturation. J. Agric. Food Chem. 2003, 51, 6280–6286. [Google Scholar] [CrossRef]
- Selli, S.; Canbas, A.; Cabaroglu, T.; Erten, H.; Lepoutre, J.P.; Gunata, Z. Effect of Skin Contact on the Free and Bound Aroma Compounds of the White Wine of Vitis Vinifera L. Cv Narince. Food Control 2006, 17, 75–82. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Days | 0 | 15 | 22 | 32 | 36 | 70 | 73 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Treated | Control | Treated | Control | Treated | Control | Treated | Control | Treated | Control | Treated | ||
| % | 0 | 21.14 de | 16.40 e | 22.11 de | 25.52 de | 23.11 de | 28.26 cde | 25.87 de | 30.76 cde | 34.73 bc | 44.38 abc | 45.78 ab | 47.08 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santin, M.; Brizzolara, S.; Castagna, A.; Ranieri, A.; Tonutti, P. Short-Term CO2 Treatment of Harvested Grapes (Vitis vinifera L., cv. Trebbiano) before Partial Dehydration Affects Berry Secondary Metabolism and the Aromatic Profile of the Resulting Wine. Plants 2022, 11, 1973. https://doi.org/10.3390/plants11151973
Santin M, Brizzolara S, Castagna A, Ranieri A, Tonutti P. Short-Term CO2 Treatment of Harvested Grapes (Vitis vinifera L., cv. Trebbiano) before Partial Dehydration Affects Berry Secondary Metabolism and the Aromatic Profile of the Resulting Wine. Plants. 2022; 11(15):1973. https://doi.org/10.3390/plants11151973
Chicago/Turabian StyleSantin, Marco, Stefano Brizzolara, Antonella Castagna, Annamaria Ranieri, and Pietro Tonutti. 2022. "Short-Term CO2 Treatment of Harvested Grapes (Vitis vinifera L., cv. Trebbiano) before Partial Dehydration Affects Berry Secondary Metabolism and the Aromatic Profile of the Resulting Wine" Plants 11, no. 15: 1973. https://doi.org/10.3390/plants11151973
APA StyleSantin, M., Brizzolara, S., Castagna, A., Ranieri, A., & Tonutti, P. (2022). Short-Term CO2 Treatment of Harvested Grapes (Vitis vinifera L., cv. Trebbiano) before Partial Dehydration Affects Berry Secondary Metabolism and the Aromatic Profile of the Resulting Wine. Plants, 11(15), 1973. https://doi.org/10.3390/plants11151973