
Flooding and Soil Properties Control Plant Intra- and Interspecific Interactions in Salt Marshes

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Intra- and Interspecific Competition Affect Plant Growth Traits
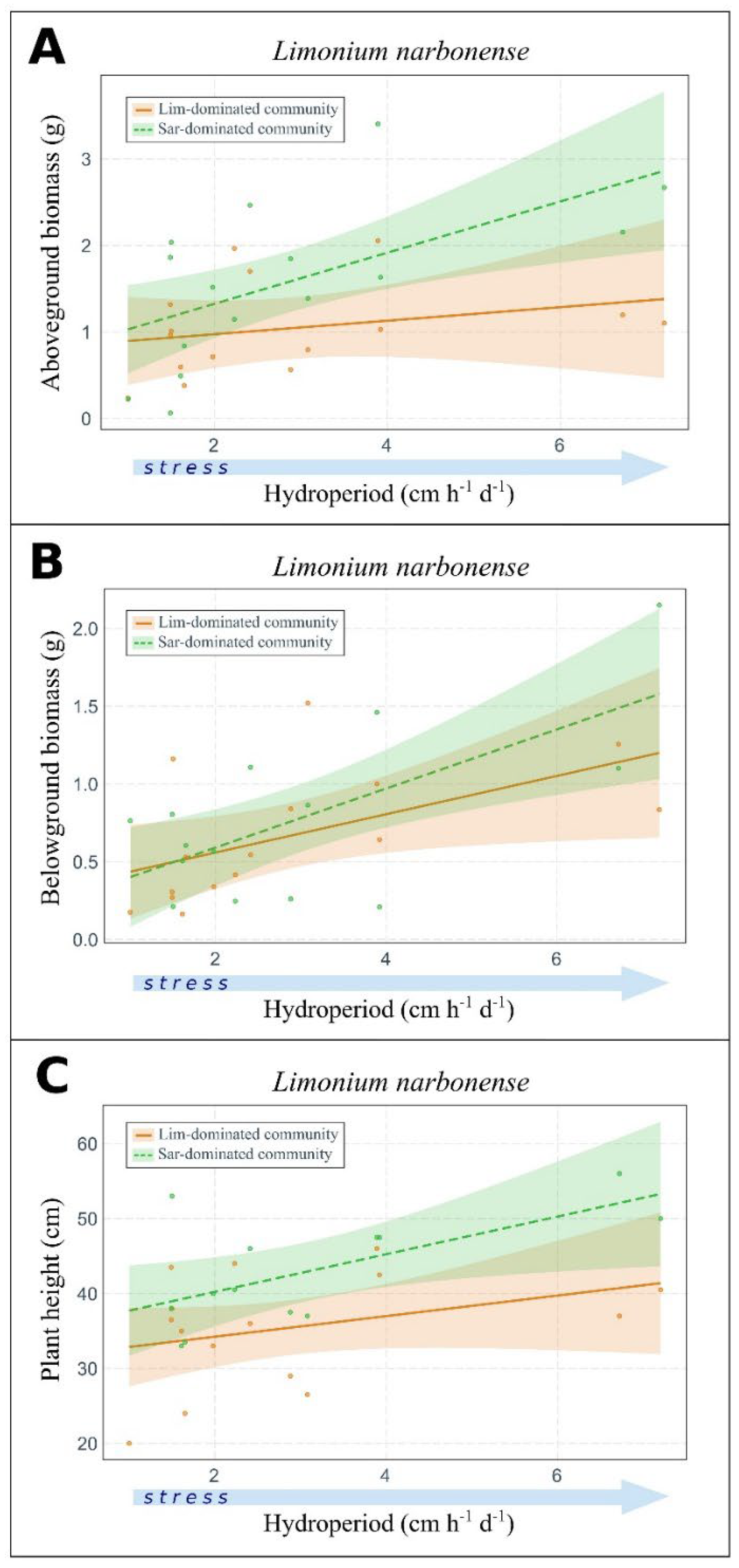
2.2. Plant Community Does Not Influence Flooding Stress Responses
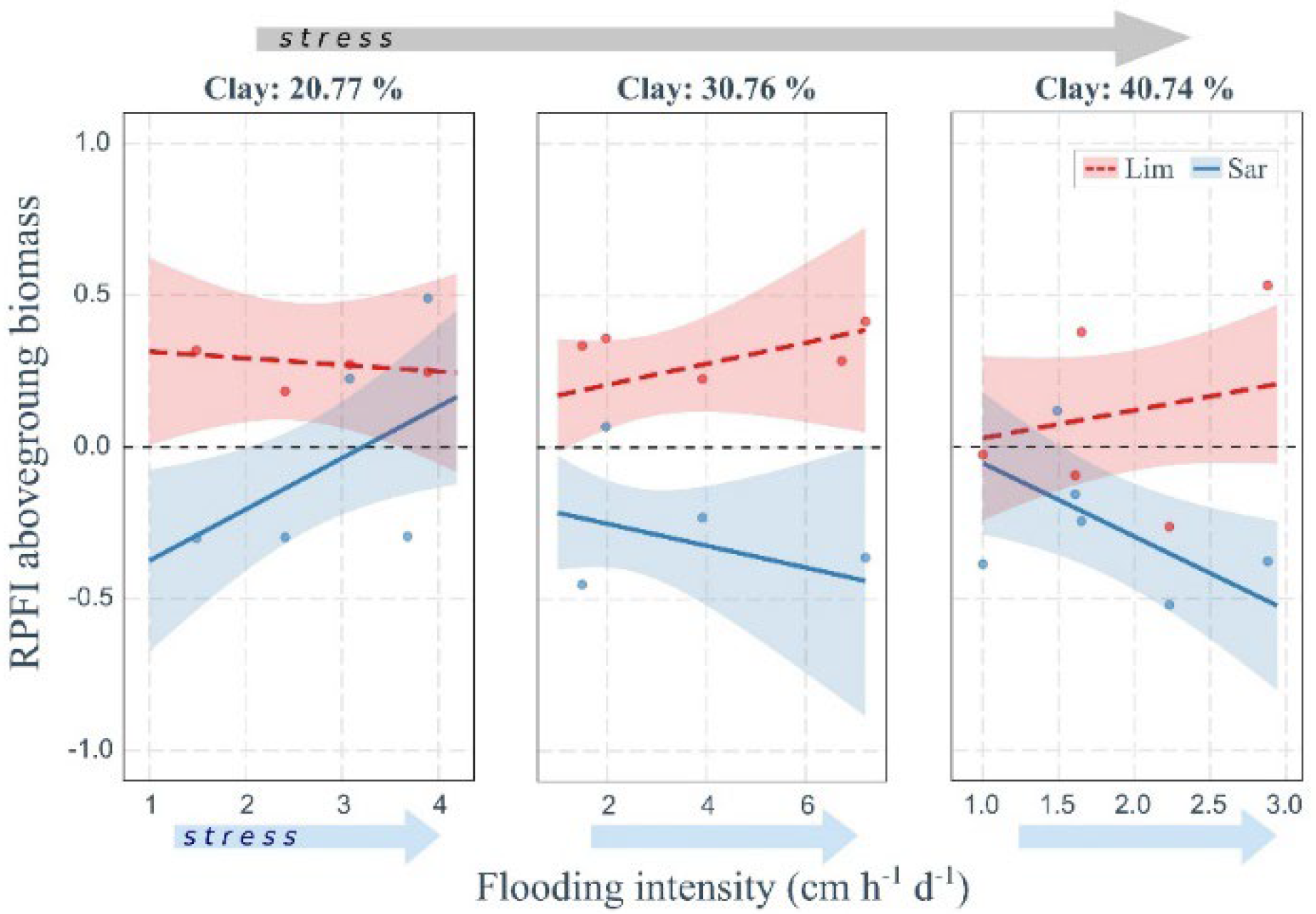
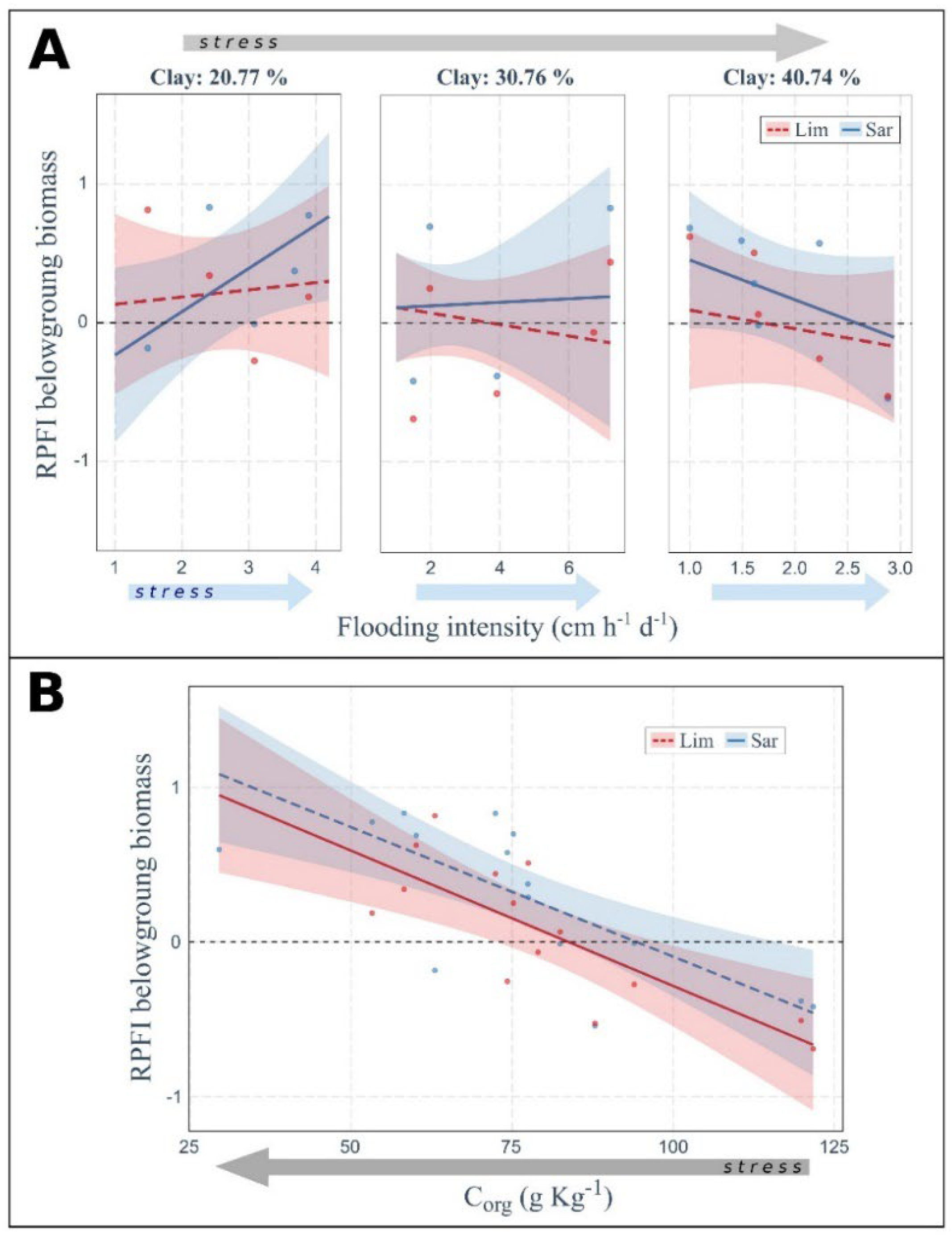
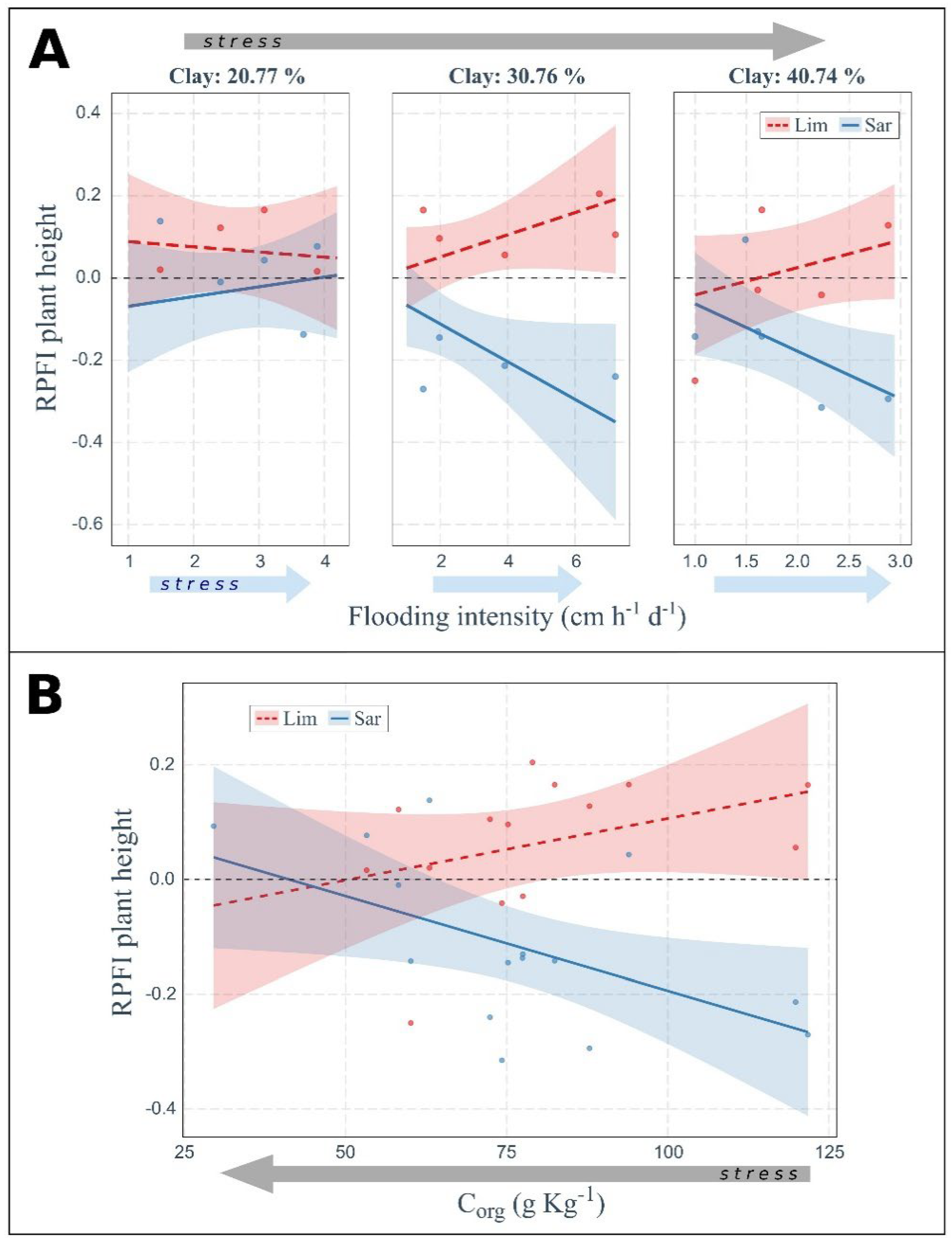
2.3. Soil Properties and Flooding Drive Plant Fitness Response
3. Discussion
3.1. Effect of Plant-Plant Interactions on Plant Traits
3.2. Effect of Flooding on Plant Traits
3.3. Effect of Flooding and Soil Texture on Plant Fitness
3.4. Effect of Flooding and Soil Organic C on Plant Fitness
4. Conclusions
5. Materials and Methods
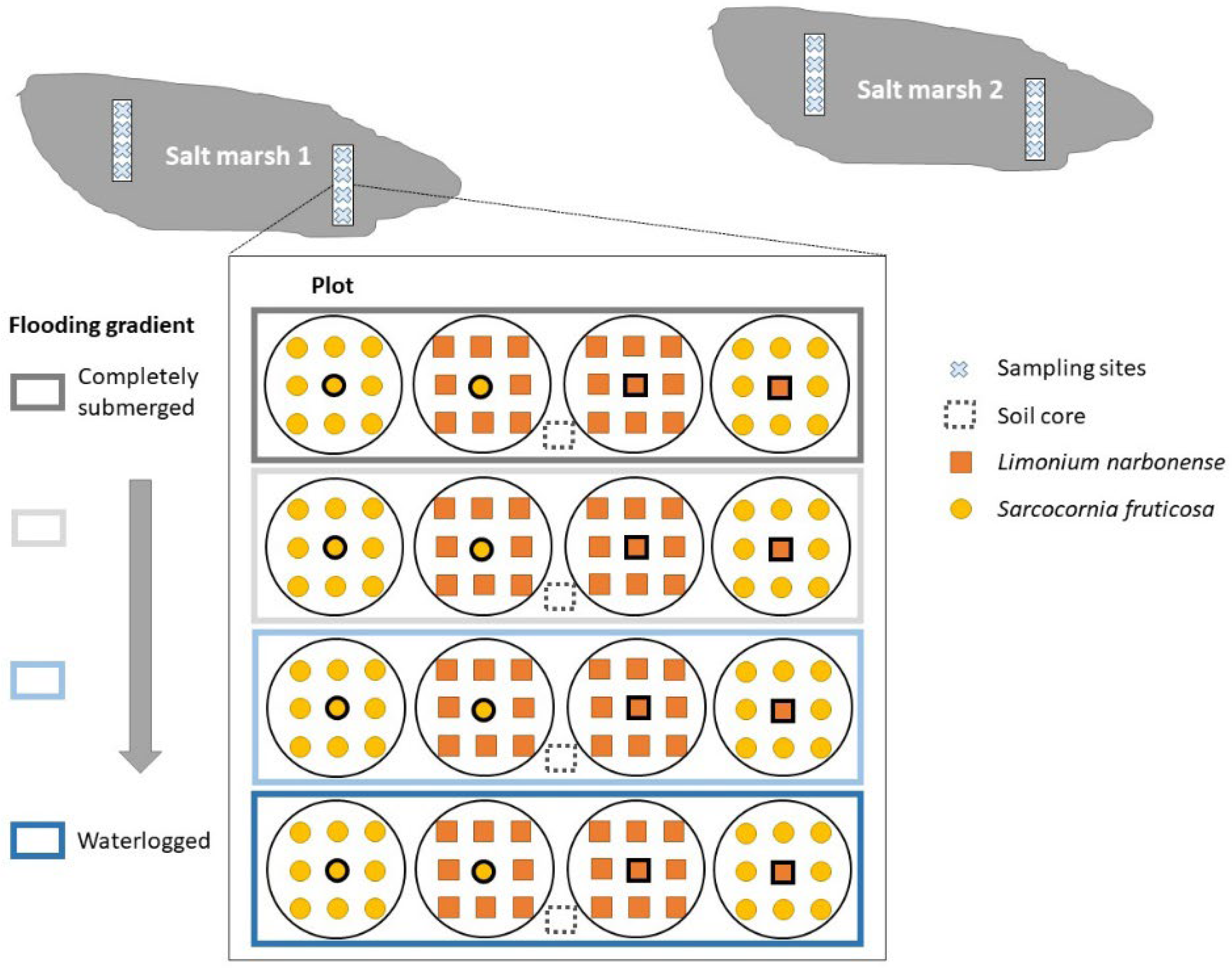
5.1. Study Area and Sampling Design
5.2. Flooding Stress
5.3. Plant Traits and Soil Analyses
5.4. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grime, J.P. Plant Strategies and Vegetation Processes, 1st ed.; Wiley: Chichester, UK, 1979; ISBN 978-0-471-99692-7. [Google Scholar]
- Pezeshki, S.R.; DeLaune, R.D. Soil oxidation-reduction in wetlands and its impact on plant functioning. Biology 2012, 1, 196–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davy, A.J.; Brown, M.J.H.; Mossman, H.L.; Grant, A. Colonization of a newly developing salt marsh: Disentangling independent effects of elevation and redox potential on halophytes. J. Ecol. 2011, 99, 1350–1357. [Google Scholar] [CrossRef]
- Ponnamperuma, F.N. Effects of Flooding on Soils. In Flooding and Plant Growth; Kozlowski, T., Ed.; Academic Press: New York, NY, USA, 1984; pp. 9–45. [Google Scholar]
- Colmer, T.D. Long-distance transport of gases in plants: A perspective on internal aeration and radial oxygen loss from roots. Plant Cell Environ. 2003, 26, 17–36. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, O.; Colmer, T.; Sand-Jensen, K. Underwater photosynthesis of submerged plants-recent advances and methods. Front. Plant Sci. 2013, 4, 140. [Google Scholar] [CrossRef] [Green Version]
- Winkel, A.; Colmer, T.D.; Pedersen, O. Leaf gas films of Spartina anglica enhance rhizome and root oxygen during tidal submergence. Plant Cell Environ. 2011, 34, 2083–2092. [Google Scholar] [CrossRef]
- Brooker, R.W.; Maestre, F.T.; Callaway, R.M.; Lortie, C.L.; Cavieres, L.A.; Kunstler, G.; Liancourt, P.; Tielbörger, K.; Travis, J.M.J.; Anthelme, F.; et al. Facilitation in plant communities: The past, the present, and the future. J. Ecol. 2008, 96, 18–34. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, E.; Boscutti, F.; De Nobili, M.; Casolo, V. Plant traits shape the effects of tidal flooding on soil and plant communities in saltmarshes. Plant Ecol 2018, 219, 823–835. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Brooker, R.W.; Scott, D.; Palmer, S.C.F.; Swaine, E. Transient facilitative effects of heather on scots pine along a grazing disturbance gradient in Scottish moorland. J. Ecol. 2006, 94, 637–645. [Google Scholar] [CrossRef]
- Bertness, M.D.; Gough, L.; Shumway, S.W. Salt tolerances and the distribution of fugitive salt marsh plants. Ecology 1992, 73, 1842–1851. [Google Scholar] [CrossRef]
- Irving, A.D.; Bertness, M.D. Trait-dependent modification of facilitation on cobble beaches. Ecology 2009, 90, 3042–3050. [Google Scholar] [CrossRef] [PubMed]
- Fogel, B.N.; Crain, C.M.; Bertness, M.D. Community level engineering effects of Triglochin maritima (Seaside arrowgrass) in a salt marsh in northern New England, USA. J. Ecol. 2004, 92, 589–597. [Google Scholar] [CrossRef]
- Levine, J.M.; Hacker, S.D.; Harley, C.D.G.; Bertness, M.D. nitrogen effects on an interaction chain in a salt marsh community. Oecologia 1998, 117, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Maestre, F.T.; Callaway, R.M.; Valladares, F.; Lortie, C.J. Refining the stress-gradient hypothesis for competition and facilitation in plant communities. J. Ecol. 2009, 97, 199–205. [Google Scholar] [CrossRef]
- Bonanomi, G.; Stinca, A.; Chirico, G.B.; Ciaschetti, G.; Saracino, A.; Incerti, G. Cushion plant morphology controls biogenic capability and facilitation effects of Silene acaulis along an elevation gradient. Funct. Ecol. 2016, 30, 1216–1226. [Google Scholar] [CrossRef] [Green Version]
- Huckle, J.M.; Marrs, R.H.; Potter, J.A. Interspecific and intraspecific interactions between salt marsh plants: Integrating the effects of environmental factors and density on plant performance. Oikos 2002, 96, 307–319. [Google Scholar] [CrossRef]
- García-Cervigón, A.I.; Gazol, A.; Sanz, V.; Camarero, J.J.; Olano, J.M. Intraspecific competition replaces interspecific facilitation as abiotic stress decreases: The shifting nature of plant–plant interactions. Perspect. Plant Ecol. Evol. Syst. 2013, 15, 226–236. [Google Scholar] [CrossRef]
- Callaway, R.M.; Brooker, R.W.; Choler, P.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Paolini, L.; Pugnaire, F.I.; Newingham, B.; Aschehoug, E.T.; et al. Positive interactions among alpine plants increase with stress. Nature 2002, 417, 844–848. [Google Scholar] [CrossRef]
- Daleo, P.; Iribarne, O. Beyond Competition: The stress-gradient hypothesis tested in plant-herbivore interactions. Ecology 2009, 90, 2368–2374. [Google Scholar] [CrossRef]
- Maestre, F.T.; Cortina, J. Do positive interactions increase with abiotic stress? A test from a semi-arid steppe. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2004, 271, S331–S333. [Google Scholar] [CrossRef] [Green Version]
- Barchuk, A.H.; Valiente-Banuet, A.; Díaz, M.P. Effect of shrubs and seasonal variability of rainfall on the establishment of Aspidosperma quebracho-blanco in two edaphically contrasting environments. Austral Ecol. 2005, 30, 695–705. [Google Scholar] [CrossRef]
- Zhang, L.; Shao, H. Direct plant-plant facilitation in coastal wetlands: A review. Estuar. Coast. Shelf Sci. 2013, 119, 1–6. [Google Scholar] [CrossRef]
- Malkinson, D.; Tielbörger, K. What does the stress-gradient hypothesis predict? resolving the discrepancies. Oikos 2010, 119, 1546–1552. [Google Scholar] [CrossRef]
- Patel, K.F.; Fansler, S.J.; Campbell, T.P.; Bond-Lamberty, B.; Smith, A.P.; RoyChowdhury, T.; McCue, L.A.; Varga, T.; Bailey, V.L. Soil texture and environmental conditions influence the biogeochemical responses of soils to drought and flooding. Commun. Earth Environ. 2021, 2, 127. [Google Scholar] [CrossRef]
- Vittori Antisari, L.; Ferronato, C.; Pellegrini, E.; Boscutti, F.; Casolo, V.; de Nobili, M.; Vianello, G. Soil properties and plant community relationship in a saltmarsh of the Grado and Marano Lagoon (northern Italy). J. Soils Sediments 2017, 17, 1862–1873. [Google Scholar] [CrossRef] [Green Version]
- Huckle, J.M.; Potter, J.A.; Marrs, R.H. Influence of environmental factors on the growth and interactions between salt marsh plants: Effects of salinity, sediment and waterlogging. J. Ecol. 2000, 88, 492–505. [Google Scholar] [CrossRef]
- Lima, T.R.A.; Martins, F.R.; Menezes, B.S.; Marquitti, F.M.D.; Sfair, J.C.; Silveira, A.P.; Araújo, F.S. The stress gradient hypothesis explains plant-plant interaction networks in edapho climatic gradients. Acta Oecol. 2022, 115, 103831. [Google Scholar] [CrossRef]
- Cronk, J.K.; Fennessy, M.S. Wetland Plants: Biology and Ecology; CRC Press: Boca Raton, FL, USA, 2001; ISBN 978-0-429-14901-6. [Google Scholar]
- Róis, A.S.; Rodríguez López, C.M.; Cortinhas, A.; Erben, M.; Espírito-Santo, D.; Wilkinson, M.J.; Caperta, A.D. Epigenetic rather than genetic factors may explain phenotypic divergence between coastal populations of diploid and tetraploid Limonium spp. (Plumbaginaceae) in Portugal. BMC Plant Biol. 2013, 13, 205. [Google Scholar] [CrossRef] [Green Version]
- Dietz, H.; Steinlein, T.; Ullmann, I. The role of growth form and correlated traits in competitive ranking of six perennial ruderal plant species grown in unbalanced mixtures. Acta Oecol. 1998, 19, 25–36. [Google Scholar] [CrossRef]
- Keddy, P.; Fraser, L.H.; Wisheu, I.C. A Comparative approach to examine competitive response of 48 wetland plant species. J. Veg. Sci. 1998, 9, 777–786. [Google Scholar] [CrossRef]
- Black, J.N. The significance of petiole length, leaf area, and light interception in competition between strains of subterrranean clover (Trifolium subterraneum L.) grown in swards. Aust. J. Agric. Res. 1960, 11, 277–291. [Google Scholar] [CrossRef]
- Wang, C.; Liu, J.; Xiao, H.; Zhou, J. Differences in leaf functional traits between Rhus typhina and native species. CLEAN-Soil Air Water 2016, 44, 1591c1597. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Voesenek, L.A.C.J. Flooding stress: Acclimations and genetic diversity. Annu. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Mauseth, J.D. Theoretical aspects of surface-to-volume ratios and water-storage capacities of succulent shoots. Am. J. Bot. 2000, 87, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Cieraad, E.; van Bodegom, P.M. Are ecophysiological adaptive traits decoupled from leaf economics traits in wetlands? Funct. Ecol. 2019, 33, 1202–1210. [Google Scholar] [CrossRef] [Green Version]
- Flowers, T.J.; Colmer, T.D. Salinity tolerance in halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Sun, X.-S.; Cui, Y.; Zhuo, N.; Wei, G.-W.; Luo, F.-L.; Zhang, M.-X. Interacting Flooding and Competition Negatively Affect Growth of Riparian Species Dominating a Reservoir Shoreline. Water 2021, 13, 1471. [Google Scholar] [CrossRef]
- Casanova, M.T.; Brock, M.A. How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol. 2000, 147, 237–250. [Google Scholar] [CrossRef]
- Reents, S.; Mueller, P.; Tang, H.; Jensen, K.; Nolte, S. Plant genotype determines biomass response to flooding frequency in tidal wetlands. Biogeosciences 2021, 18, 403–411. [Google Scholar] [CrossRef]
- Lan, Z.; Chen, Y.; Shen, R.; Cai, Y.; Luo, H.; Jin, B.; Chen, J. Effects of flooding duration on wetland plant biomass: The importance of soil nutrients and season. Freshw. Biol. 2021, 66, 211–222. [Google Scholar] [CrossRef]
- Silvestri, S.; Defina, A.; Marani, M. Tidal regime, salinity and salt marsh plant zonation. Estuar. Coast. Shelf Sci. 2005, 62, 119–130. [Google Scholar] [CrossRef]
- Pellegrini, E.; Forlani, G.; Boscutti, F.; Casolo, V. Evidence of non-structural carbohydrates-mediated response to flooding and salinity in Limonium narbonense and Salicornia fruticosa. Aquat. Bot. 2020, 166, 103265. [Google Scholar] [CrossRef]
- Lou, Y.; Pan, Y.; Gao, C.; Jiang, M.; Lu, X.; Xu, Y.J. Response of plant height, species richness and aboveground biomass to flooding gradient along vegetation zones in floodplain wetlands, northeast China. PLoS ONE 2016, 11, e0153972. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, E.; Konnerup, D.; Winkel, A.; Casolo, V.; Pedersen, O. Contrasting oxygen dynamics in Limonium narbonense and Sarcocornia fruticosa during partial and complete submergence. Funct. Plant Biol. 2017, 44, 867–876. [Google Scholar] [CrossRef]
- Herzog, M.; Pedersen, O. Partial versus complete submergence: Snorkelling aids root aeration in Rumex palustris but not in R. acetosa. Plant Cell Environ. 2014, 37, 2381–2390. [Google Scholar] [CrossRef]
- Colmer, T.D.; Flowers, T.J. Flooding tolerance in halophytes. New Phytol. 2008, 179, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, O.; Sauter, M.; Colmer, T.D.; Nakazono, M. Regulation of root adaptive anatomical and morphological traits during low soil oxygen. New Phytol. 2021, 229, 42–49. [Google Scholar] [CrossRef] [Green Version]
- Figueroa, M.E.; Castillo, J.M.; Redondo, S.; Luque, T.; Castellanos, E.M.; Nieva, F.J.; Luque, C.J.; Rubio-Casal, A.E.; Davy, A.J. Facilitated invasion by hybridization of Sarcocornia species in a salt-marsh succession. J. Ecol. 2003, 91, 616–626. [Google Scholar] [CrossRef]
- Zehnder, A.J.B.; Stumm, W. Geochemistry and biogeochemistry of anaerobic habitats. In Biology of Anaerobic Microorganisms; Wageningen University & Research: Wageningen, The Netherlands, 1988; pp. 1–38. [Google Scholar]
- Pellegrini, E.; Petranich, E.; Acquavita, A.; Canário, J.; Emili, A.; Covelli, S. Mercury uptake by halophytes in response to a long-term contamination in coastal wetland salt marshes (Northern Adriatic Sea). Environ. Geochem. Health 2017, 39, 1273–1289. [Google Scholar] [CrossRef]
- Boorman, L.A. Limonium vulgare Mill. and L. humile Mill. J. Ecol. 1967, 55, 221–232. [Google Scholar] [CrossRef]
- Callaway, R.M. Facilitative and interfering effects of Arthrocnemum subterminale on winter annuals. Ecology 1994, 75, 681–686. [Google Scholar] [CrossRef]
- Bradley, P.M.; Morris, J.T. Physical characteristics of salt marsh sediments: Ecological implications. Mar. Ecol. Prog. Ser. 1990, 61, 245–252. [Google Scholar] [CrossRef]
- Gul, B.; Khan, M.A. Effect of intraspecific competition and inundation regime on the growth of Arthrocnemum macrostachyum in a coastal swamp in Karachi, Pakistan. Pak. J. Bot. 1999, 31, 163–172. [Google Scholar]
- Callaway, R.M.; Walker, L.R. Competition and facilitation: A synthetic approach to interactions in plant communities. Ecology 1997, 78, 1958–1965. [Google Scholar] [CrossRef]
- Choler, P.; Michalet, R.; Callaway, R.M. Facilitation and competition on gradients in alpine plant communities. Ecology 2001, 82, 3295–3308. [Google Scholar] [CrossRef]
- Soliveres, S.; Eldridge, D.J.; Maestre, F.T.; Bowker, M.A.; Tighe, M.; Escudero, A. Microhabitat Amelioration and reduced competition among understorey plants as drivers of facilitation across environmental gradients: Towards a unifying framework. Perspect. Plant Ecol. Evol. Syst. 2011, 13, 247–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamers, L.; Govers, L.; Janssen, I.; Geurts, J.; Van der Welle, M.; Van Katwijk, M.; Van der Heide, T.; Roelofs, J.; Smolders, A. Sulfide as a soil phytotoxin—a review. Front. Plant Sci. 2013, 4, 268. [Google Scholar] [CrossRef] [Green Version]
- Kylafis, G.; Loreau, M. Niche construction in the light of niche theory. Ecol. Lett. 2011, 14, 82–90. [Google Scholar] [CrossRef]
- Vittori Antisari, L.; De Nobili, M.; Ferronato, C.; Natale, M.; Pellegrini, E.; Vianello, G. Hydromorphic to subaqueous soils transitions in the central Grado lagoon (northern Adriatic Sea, Italy). Estuar. Coast. Shelf Sci. 2016, 173, 39–48. [Google Scholar] [CrossRef]
- Pellegrini, E.; Contin, M.; Livia, V.A.; Chiara, F.; Nobili, M.D. Soil organic carbon and carbonates are binding phases for simultaneously extractable metals (SEM) in calcareous saltmarsh soils. Environ. Toxicol. Chem. 2019, 38, 2688–2697. [Google Scholar] [CrossRef]
- Carrasco-Barea, L.; Llorens, L.; Romaní, A.M.; Gispert, M.; Verdaguer, D. Litter decomposition of three halophytes in a Mediterranean salt marsh: Relevance of litter quality, microbial activity and microhabitat. Sci. Total Environ. 2022, 838, 155743. [Google Scholar] [CrossRef] [PubMed]
- Buth, G.J.C.; Voesenek, L.A.C.J. Decomposition of standing and fallen litter of halophytes in a Dutch salt marsh. In Vegetation between Land and Sea: Structure and Processes; Huiskes, A.H.L., Blom, C.W.P.M., Rozema, J., Eds.; Geobotany; Springer: Dordrecht, The Netherlands, 1987; pp. 146–162. ISBN 978-94-009-4065-9. [Google Scholar]
- Ret, M. Bilancio Idrologico e Circolazione Idrica della Laguna di Marano e Grado. Master’s Thesis, Faculty of Engineering, University of Udine, Udine, Italy, 2006. [Google Scholar]
- Armas, C.; Ordiales, R.; Pugnaire, F.I. Measuring plant interactions: A new comparative index. Ecology 2004, 85, 2682–2686. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J. Nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-98. Available online: http://cran.r-project.org/package=nlme (accessed on 1 November 2021).
- Russell, L. Emmeans: Estimated Marginal Means, aka Least-Squares Means, R Package Version 2018, 1. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 1 November 2021).
- R Core Team R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Minimum | Maximum | N | Mean ± SD | |
|---|---|---|---|---|
| Limonium narbonense | ||||
| Plant height (cm) | 12.0 | 56.0 | 30 | 38.0 ± 9.6 |
| Aboveground biomass (g) | 0.06 | 3.40 | 29 | 1.31 ± 0.80 |
| Belowground biomass (g) | 0.16 | 2.15 | 29 | 0.72 ± 0.48 |
| Sarcocornia fruticosa | ||||
| Plant height (cm) | 14.2 | 34.2 | 32 | 23.6 ± 5.8 |
| Aboveground biomass (g) | 0.45 | 4.63 | 32 | 2.21 ± 1.12 |
| Belowground biomass (g) | 0.01 | 0.40 | 32 | 0.13 ± 0.09 |
| Flooding intensity (cm h−1 d−1) | 1.0 | 7.2 | 16 | 2.9 ± 1.8 |
| Soil texture | ||||
| Clay (%) | 11 | 51 | 16 | 31.4 ± 10.2 |
| Silt (%) | 5 | 31 | 16 | 16.3 ± 8.4 |
| Sand (%) | 34 | 83 | 16 | 52.3 ± 14.4 |
| Soil organic C (%) | 3.0 | 12.2 | 16 | 7.7 ± 2.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pellegrini, E.; Incerti, G.; Pedersen, O.; Moro, N.; Foscari, A.; Casolo, V.; Contin, M.; Boscutti, F. Flooding and Soil Properties Control Plant Intra- and Interspecific Interactions in Salt Marshes. Plants 2022, 11, 1940. https://doi.org/10.3390/plants11151940
Pellegrini E, Incerti G, Pedersen O, Moro N, Foscari A, Casolo V, Contin M, Boscutti F. Flooding and Soil Properties Control Plant Intra- and Interspecific Interactions in Salt Marshes. Plants. 2022; 11(15):1940. https://doi.org/10.3390/plants11151940
Chicago/Turabian StylePellegrini, Elisa, Guido Incerti, Ole Pedersen, Natasha Moro, Alessandro Foscari, Valentino Casolo, Marco Contin, and Francesco Boscutti. 2022. "Flooding and Soil Properties Control Plant Intra- and Interspecific Interactions in Salt Marshes" Plants 11, no. 15: 1940. https://doi.org/10.3390/plants11151940
APA StylePellegrini, E., Incerti, G., Pedersen, O., Moro, N., Foscari, A., Casolo, V., Contin, M., & Boscutti, F. (2022). Flooding and Soil Properties Control Plant Intra- and Interspecific Interactions in Salt Marshes. Plants, 11(15), 1940. https://doi.org/10.3390/plants11151940