
A Viral Suppressor of RNA Silencing May Be Targeting a Plant Defence Pathway Involving Fibrillarin


Abstract
:1. Introduction
2. Results
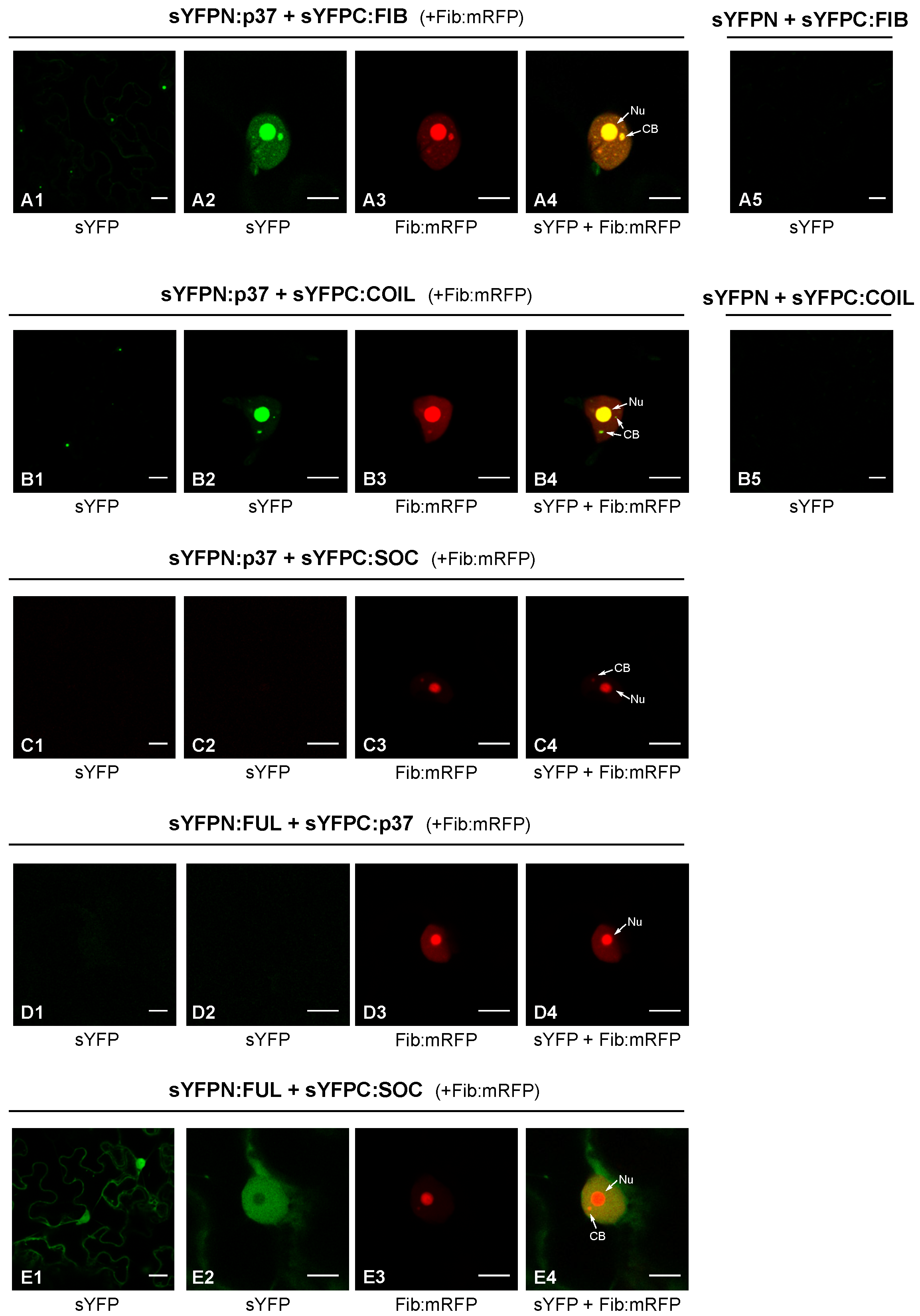
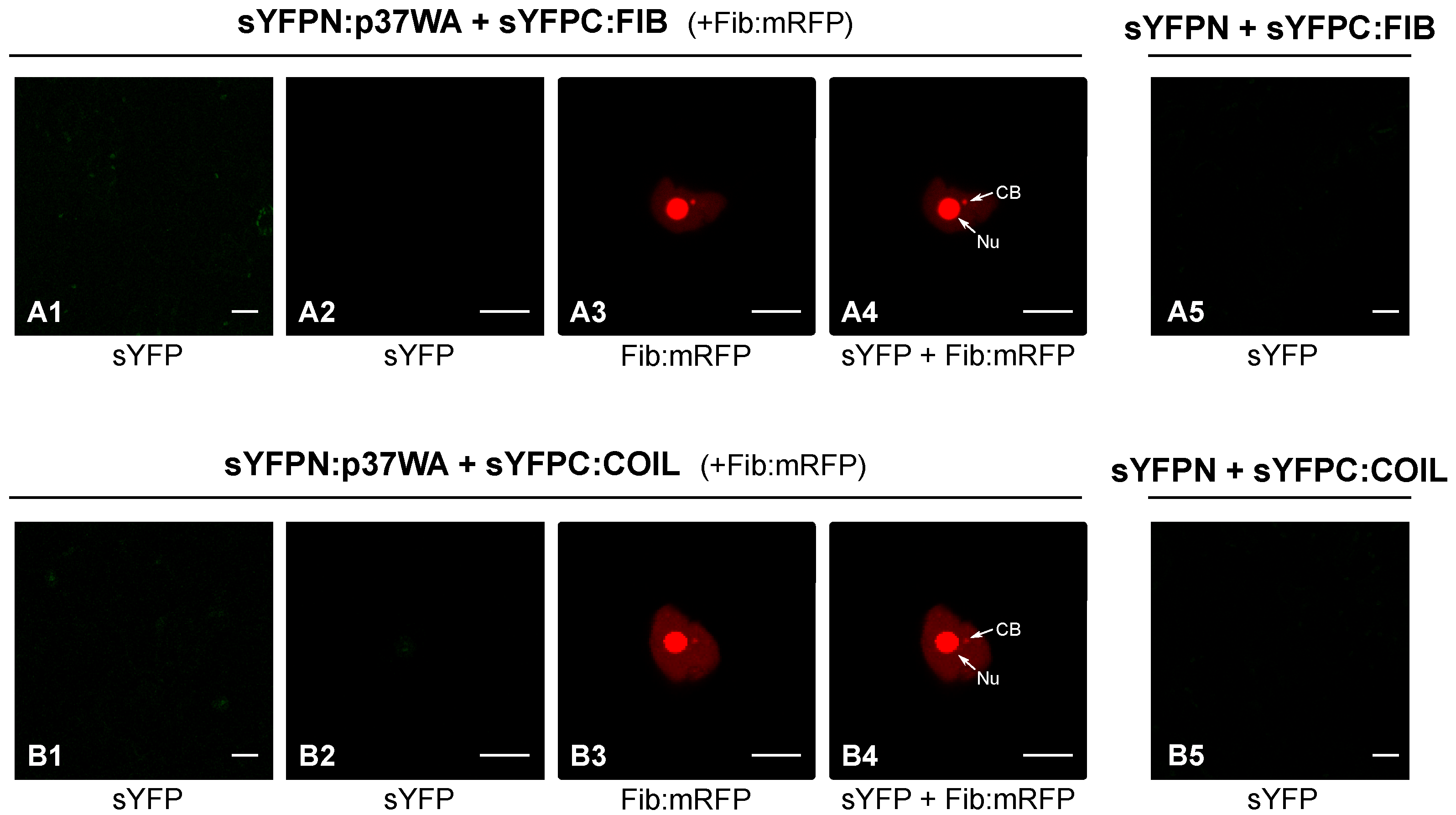
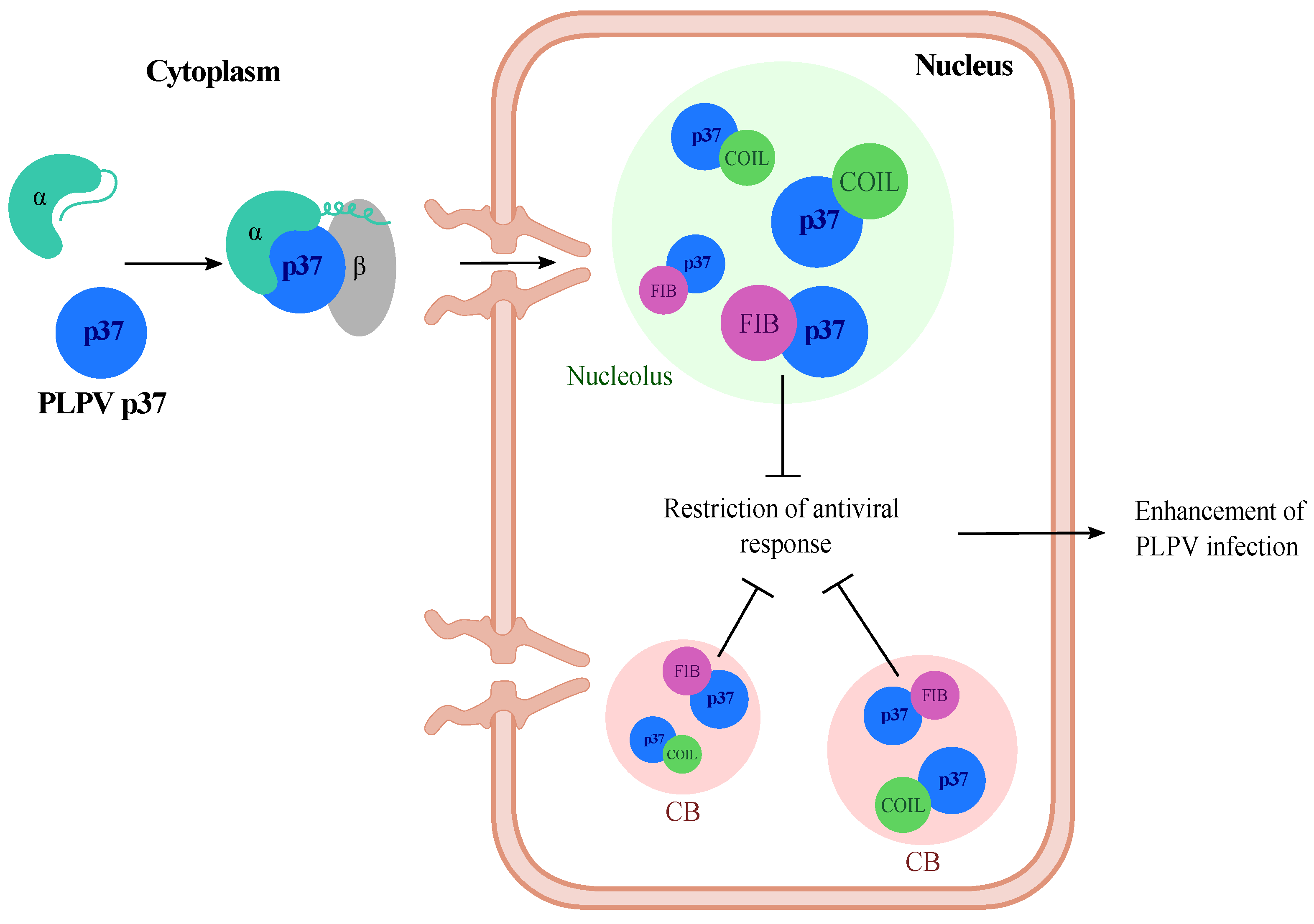
2.1. P37 Interacts In Vivo with Both Fibrillarin and Coilin
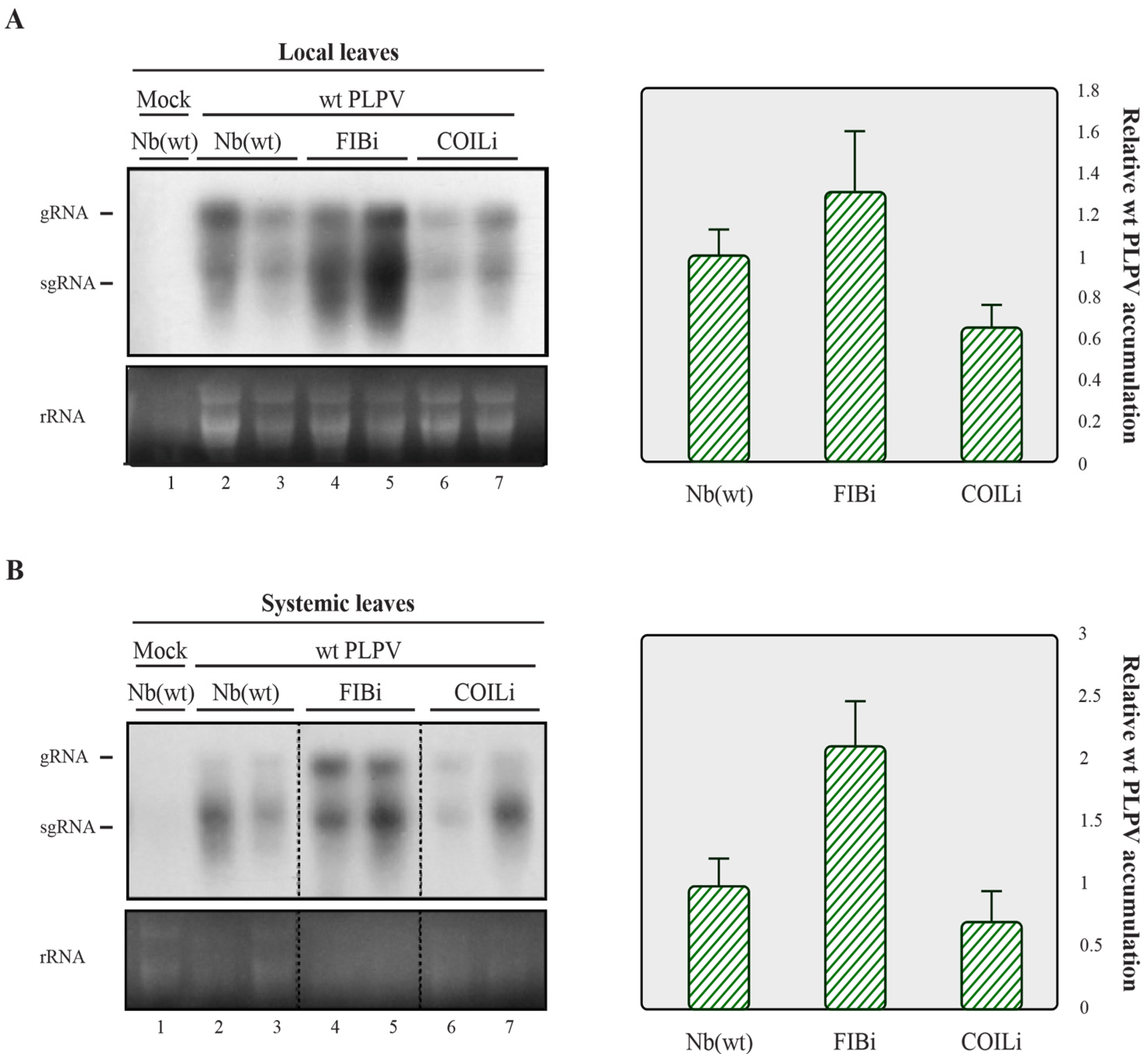
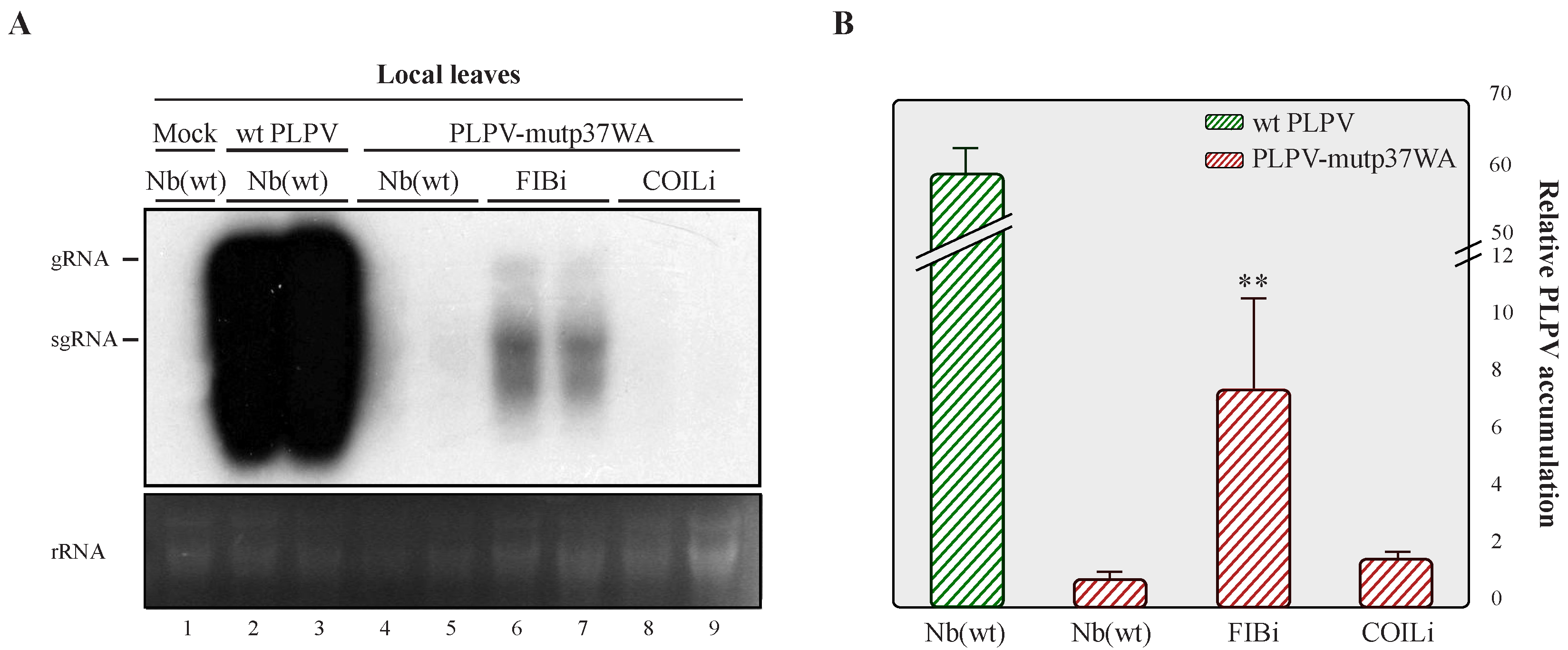
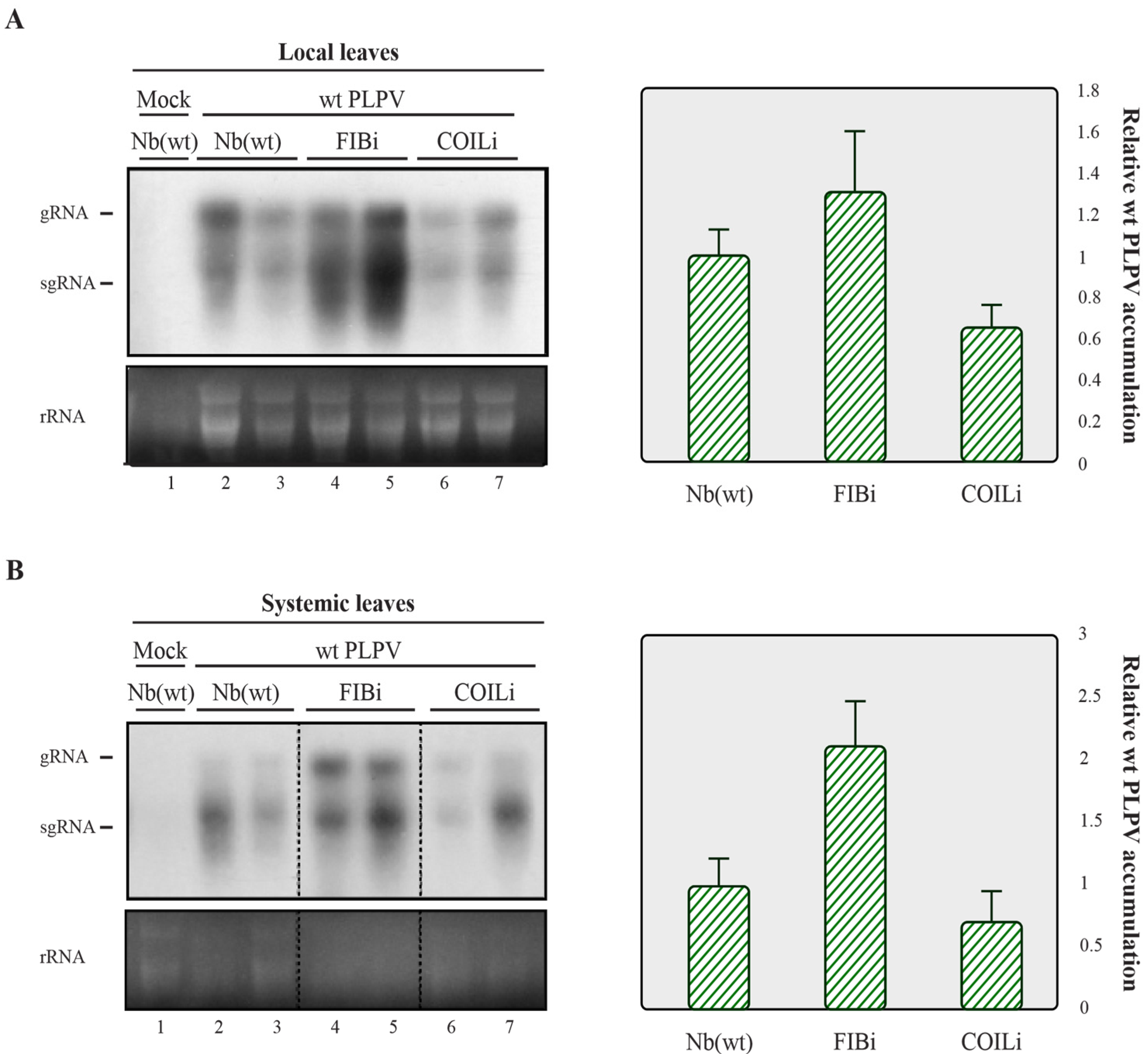
2.2. Downregulation of Fibrillarin but Not Coilin Results in Heightened Accumulation of an VSR Defective-PLPV
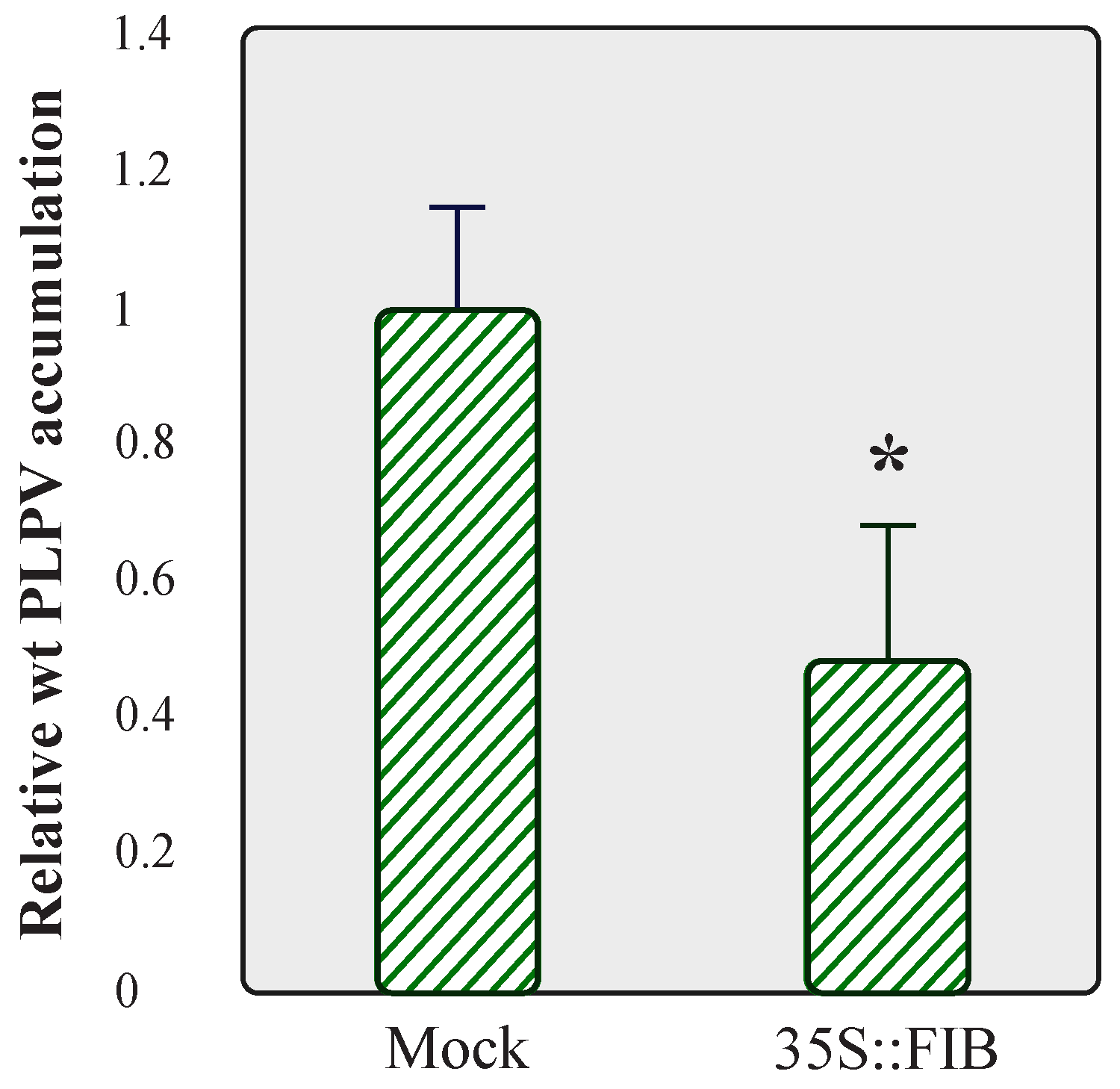
2.3. Overexpression of Fibrillarin Has a Negative Effect on PLPV Accumulation
2.4. Fibrillarin Expression Levels Are Not Affected by PLPV Infection
3. Discussion
4. Materials and Methods
4.1. DNA Constructs
4.2. Agrobacterium Tumefaciens Transformation and Agroinfiltration of N. benthamiana Plants
4.3. Confocal Microscopy
4.4. Co-Immunoprecipitation Assays
4.5. RNA Extraction and Northern Blot Hybridization
4.6. RT-qPCR
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramage, H.; Cherry, S. Virus-Host Interactions: From Unbiased Genetic Screens to Function. Annu. Rev. Virol. 2015, 2, 497–524. [Google Scholar] [CrossRef] [PubMed]
- García, J.A.; Pallás, V. Viral Factors Involved in Plant Pathogenesis. Curr. Opin. Virol. 2015, 11, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, A.; García, J.A.; Simón-Mateo, C.; Hernández, C. Plant Virus RNA Replication. In eLS; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2016. [Google Scholar] [CrossRef] [Green Version]
- Walker, E.J.; Ghildyal, R. Editorial: Viral Interactions with the Nucleus. Front. Microbiol. 2017, 8, 951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iarovaia, O.V.; Minina, E.P.; Sheval, E.V.; Onichtchouk, D.; Dokudovskaya, S.; Razin, S.V.; Vassetzky, Y.S. Nucleolus: A Central Hub for Nuclear Functions. Trends Cell Biol. 2019, 29, 647–659. [Google Scholar] [CrossRef]
- Trinkle-Mulcahy, L.; Sleeman, J.E. The Cajal Body and the Nucleolus: “In a Relationship” or “It’s Complicated”? RNA Biol. 2016, 14, 739–751. [Google Scholar] [CrossRef] [Green Version]
- Love, A.J.; Yu, C.; Petukhova, N.V.; Kalinina, N.O.; Chen, J.; Taliansky, M.E. Cajal Bodies and Their Role in Plant Stress and Disease Responses. RNA Biol. 2016, 14, 779–790. [Google Scholar] [CrossRef] [Green Version]
- Kalinina, N.O.; Makarova, S.; Makhotenko, A.; Love, A.J.; Taliansky, M. The Multiple Functions of the Nucleolus in Plant Development, Disease and Stress Responses. Front. Plant Sci. 2018, 9, 132. [Google Scholar] [CrossRef]
- Kim, S.H.; Macfarlane, S.; Kalinina, N.O.; Rakitina, D.V.; Ryabov, E.V.; Gillespie, T.; Haupt, S.; Brown, J.W.S.; Taliansky, M. Interaction of a Plant Virus-Encoded Protein with the Major Nucleolar Protein Fibrillarin Is Required for Systemic Virus Infection. Proc. Natl. Acad. Sci. USA 2007, 104, 11115–11120. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.-H.; Hsu, F.-C.; Lee, S.-C.; Lo, Y.-S.; Wang, J.-D.; Shaw, J.; Taliansky, M.; Chang, B.-Y.; Hsu, Y.-H.; Lin, N.-S. The Nucleolar Fibrillarin Protein Is Required for Helper Virus-Independent Long-Distance Trafficking of a Subviral Satellite RNA in Plants. Plant Cell 2016, 28, 2586–2602. [Google Scholar] [CrossRef]
- Shaw, J.; Love, A.J.; Makarova, S.S.; Kalinina, N.O.; Harrison, B.D.; Taliansky, M.E. Coilin, the Signature Protein of Cajal Bodies, Differentially Modulates the Interactions of Plants with Viruses in Widely Different Taxa. Nucleus 2014, 5, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Du, Z.; Lin, C.; Mao, Q.; Wu, K.; Wu, J.; Wei, T.; Wu, Z.; Xie, L. Rice Stripe Tenuivirus P2 May Recruit or Manipulate Nucleolar Functions through an Interaction with Fibrillarin to Promote Virus Systemic Movement. Mol. Plant Pathol. 2015, 16, 921–930. [Google Scholar] [CrossRef] [PubMed]
- Scheets, K.; Jordan, R.; White, K.A.; Hernández, C. Pelarspovirus, a Proposed New Genus in the Family Tombusviridae. Arch. Virol. 2015, 160, 2385–2393. [Google Scholar] [CrossRef] [Green Version]
- Castaño, A.; Ruiz, L.; Hernández, C. Insights into the Translational Regulation of Biologically Active Open Reading Frames of Pelargonium Line Pattern Virus. Virology 2009, 386, 417–426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castaño, A.; Hernández, C. Complete Nucleotide Sequence and Genome Organization of Pelargonium Line Pattern Virus and Its Relationship with the Family Tombusviridae. Arch. Virol. 2005, 150, 949–965. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cañamás, M.; Hernández, C. Key Importance of Small RNA Binding for the Activity of a Glycine-Tryptophan (GW) Motif-Containing Viral Suppressor of RNA Silencing. J. Biol. Chem. 2015, 290, 3106–3120. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Li, Y. Dissection of RNAi-Based Antiviral Immunity in Plants. Curr. Opin. Virol. 2018, 32, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Kontra, L.; Burgyán, J. Viral Silencing Suppressors: Tools Forged to Fine-Tune Host-Pathogen Coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Cañamás, M.; Hernández, C. New Insights into the Nucleolar Localization of a Plant RNA Virus-Encoded Protein That Acts in Both RNA Packaging and RNA Silencing Suppression: Involvement of Importins Alpha and Relevance for Viral Infection. Mol. Plant Microbe Interact. 2018, 31, 1134–1144. [Google Scholar] [CrossRef] [Green Version]
- Balanzà, V.; Martínez-Fernández, I.; Ferrándiz, C. Sequential Action of FRUITFULL as a Modulator of the Activity of the Floral Regulators SVP and SOC1. J. Exp. Bot. 2014, 65, 1193–1203. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.; Yu, C.; Makhotenko, A.V.; Makarova, S.S.; Love, A.J.; Kalinina, N.O.; MacFarlane, S.; Chen, J.; Taliansky, M.E. Interaction of a Plant Virus Protein with the Signature Cajal Body Protein Coilin Facilitates Salicylic Acid-Mediated Plant Defence Responses. New Phytol. 2019, 224, 439–453. [Google Scholar] [CrossRef]
- Pérez-Cañamás, M.; Hevia, E.; Katsarou, K.; Hernández, C. Genetic Evidence for the Involvement of Dicer-like 2 and 4 as Well as Argonaute 2 in the Nicotiana benthamiana Response against Pelargonium Line Pattern Virus. J. Gen. Virol. 2021, 102, 001656. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ruiz, H.; Takeda, A.; Chapman, E.J.; Sullivan, C.M.; Fahlgren, N.; Brempelis, K.J.; Carrington, J.C. Arabidopsis RNA-Dependent RNA Polymerases and Dicer-like Proteins in Antiviral Defense and Small Interfering RNA Biogenesis during Turnip Mosaic Virus Infection. Plant Cell 2010, 22, 481–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ludman, M.; Burgyán, J.; Fátyol, K. Crispr/Cas9 Mediated Inactivation of Argonaute 2 Reveals Its Differential Involvement in Antiviral Responses. Sci. Rep. 2017, 7, 1010. [Google Scholar] [CrossRef]
- Sawyer, I.A.; Sturgill, D.; Dundr, M. Membraneless Nuclear Organelles and the Search for Phases within Phases. Wiley Interdiscip. Rev. RNA 2019, 10, e1514. [Google Scholar] [CrossRef]
- Wu, J.; Yang, Z.; Wang, Y.; Zheng, L.; Ye, R.; Ji, Y.; Zhao, S.; Ji, S.; Liu, R.; Xu, L.; et al. Viral-Inducible Argonaute18 Confers Broad-Spectrum Virus Resistance in Rice by Sequestering a Host MicroRNA. Elife 2015, 4, e05733. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wurm, T.; Britton, P.; Brooks, G.; Hiscox, J.A. Interaction of the Coronavirus Nucleoprotein with Nucleolar Antigens and the Host Cell. J. Virol. 2002, 76, 5233–5250. [Google Scholar] [CrossRef] [Green Version]
- Yoo, D.; Wootton, S.K.; Li, G.; Song, C.; Rowland, R.R. Colocalization and Interaction of the Porcine Arterivirus Nucleocapsid Protein with the Small Nucleolar RNA-Associated Protein Fibrillarin. J. Virol. 2003, 77, 12173–12183. [Google Scholar] [CrossRef] [Green Version]
- Ponti, D.; Troiano, M.; Bellenchi, G.C.; Battaglia, P.A.; Gigliani, F. The HIV Tat Protein Affects Processing of Ribosomal RNA Precursor. BMC Cell Biol. 2008, 9, 32. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Corona, U.; Sobol, M.; Rodriguez-Zapata, L.C.; Hozak, P.; Castano, E. Fibrillarin from Archaea to Human. Biol. Cell 2015, 107, 159–174. [Google Scholar] [CrossRef]
- Tiku, V.; Kew, C.; Mehrotra, P.; Ganesan, R.; Robinson, N.; Antebi, A. Nucleolar Fibrillarin Is an Evolutionarily Conserved Regulator of Bacterial Pathogen Resistance. Nat. Commun. 2018, 9, 3607. [Google Scholar] [CrossRef]
- Rajamäki, M.-L.; Valkonen, J.P.T. Control of Nuclear and Nucleolar Localization of Nuclear Inclusion Protein a of Picorna-like Potato Virus A in Nicotiana Species. Plant Cell 2009, 21, 2485–2502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ruiz, H. Susceptibility Genes to Plant Viruses. Viruses 2018, 10, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, A.; Castellano, M.M. Coimmunoprecipitation of Interacting Proteins in Plants. Methods Mol. Biol. 2018, 1794, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Verwoerd, T.C.; Dekker, B.M.M.; Hoekema, A. A Small-Scale Procedure for the Rapid Isolation of Plant RNAs. Nucleic Acids Res. 1989, 17, 2362. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer | Position a | Genbank Acc. No. b | Sequence c | Constructs / RT-qPCR | |
|---|---|---|---|---|---|---|
| FIB | CH674 CH675 | 1–22 (S) 924–945 (AS) | AM269909.1 | 5′-GTGGATCCATGGTTGCACCAACTAGAGGTC-3′ 5′-GGGAGCTCTAGGCAGCAGCCTTCTGCTTC-3′ | (BamHI) (SacI) | sYFPN:FIB sYFPC:FIB |
| COIL | CH712 CH713 | 142–161 (S) 2559–2583 (AS) | MK903618.1 | 5′-CAGGATCCATGGAGGGCGTTAGGCTTC -3′ 5′-CCGGTACCTCAAATTTTGTTCTGGGATCTTAG-3′ | (BamHI) (KpnI) | sYFPN:COIL sYFPC:COIL |
| PLPV p27 | CH718 CH719 | 86–105 (S) 167–185 (AS) | EU835946.1 | 5′-CGCTCCTCGGTCCTAACTTG -3′ 5′-ATTTTGGCCAACCCATGGA -3′ | RT-qPCR | |
| FIB | CH937 CH938 | 299–318 (S) 468–487 (AS) | AM269909.1 | 5′-ATTTGGTGCCTGGTGAAGCT-3′ 5′-TTCCTGATGCAGCTCCAAGG -3′ | RT-qPCR | |
| PP2A | CH436 CH437 | 996–1016 (S) 1075–1095 (AS) | MF996339.1 | 5′-ACTTGGTGCCCTTTGTATGC-3′ 5′-TGGACCAAATTCTTCTGCAA-3′ | RT-qPCR | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Cañamás, M.; Taliansky, M.; Hernández, C. A Viral Suppressor of RNA Silencing May Be Targeting a Plant Defence Pathway Involving Fibrillarin. Plants 2022, 11, 1903. https://doi.org/10.3390/plants11151903
Pérez-Cañamás M, Taliansky M, Hernández C. A Viral Suppressor of RNA Silencing May Be Targeting a Plant Defence Pathway Involving Fibrillarin. Plants. 2022; 11(15):1903. https://doi.org/10.3390/plants11151903
Chicago/Turabian StylePérez-Cañamás, Miryam, Michael Taliansky, and Carmen Hernández. 2022. "A Viral Suppressor of RNA Silencing May Be Targeting a Plant Defence Pathway Involving Fibrillarin" Plants 11, no. 15: 1903. https://doi.org/10.3390/plants11151903
APA StylePérez-Cañamás, M., Taliansky, M., & Hernández, C. (2022). A Viral Suppressor of RNA Silencing May Be Targeting a Plant Defence Pathway Involving Fibrillarin. Plants, 11(15), 1903. https://doi.org/10.3390/plants11151903