
Metabolites Produced by Fungi against Fungal Phytopathogens: Review, Implementation and Perspectives



Abstract
1. Introduction: Fungal Plant Pathogens and Control Alternatives
2. Fungal Endophytes: A Promising Tool in Plant Production and Protection
3. Secondary Metabolites from Fungal Organisms
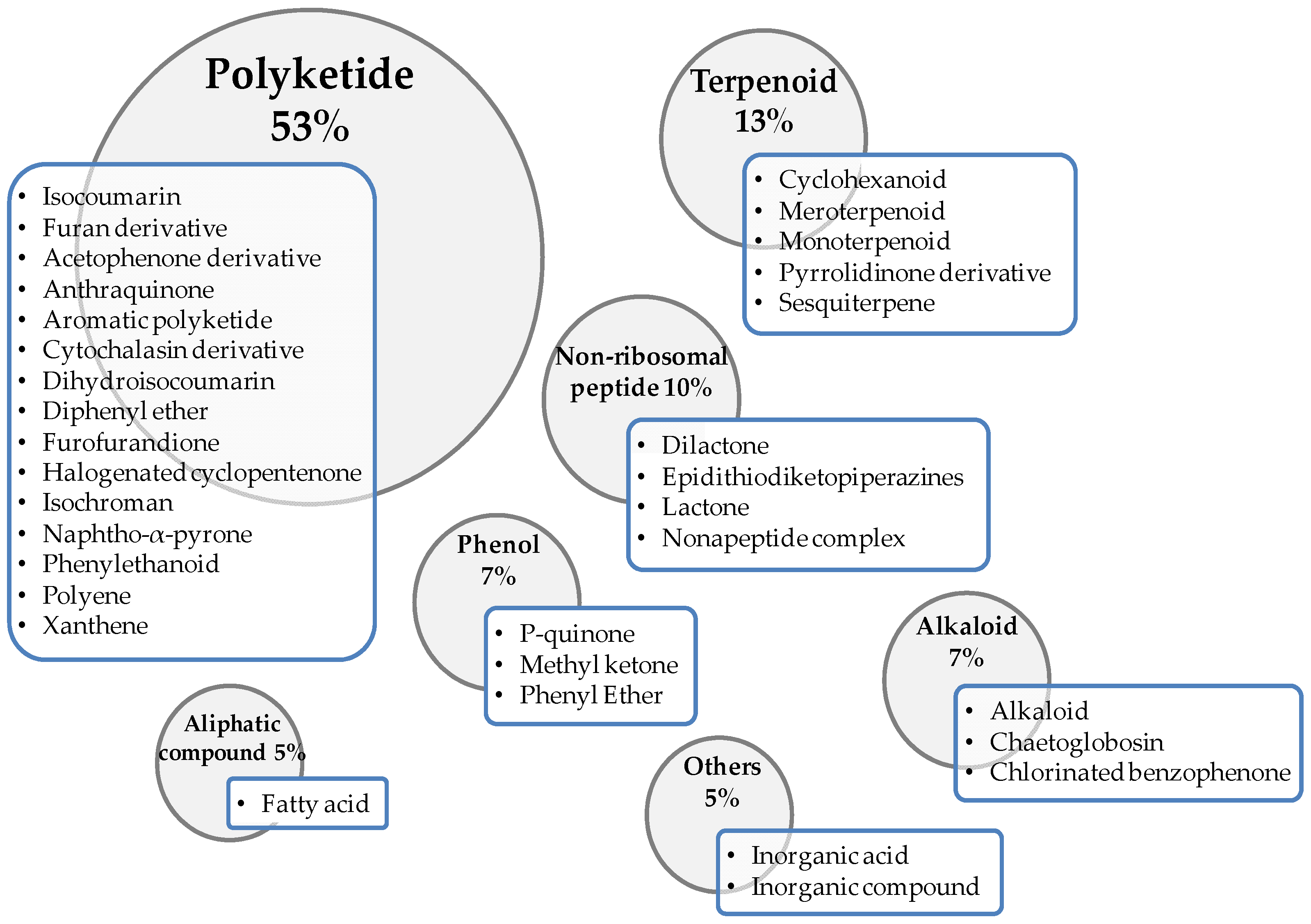
4. Secondary Metabolites of Fungi with Antifungal Properties

{kind=link}
| Metabolite(s) | Producer Fungi | Fungal Pathogen(s) | Action 1 | Conditions 2 | Efficacy 3 | Ref. |
|---|---|---|---|---|---|---|
| (12R)-12-Hydroxymonocerin | Exserohilum sp. | Fusarium oxysporum | A | IV | 3% | [104] |
| 3-(5-Oxo-2,5-dihydrofuran-3-yl) propanoic acid | Aspergillus tubingensis | Fusarium graminearum | A | IV | 2-fold | [105] |
| 4-Methylmellein, 4-hydroxymellein, 6-hydroxymellein, tyrosol | Penicillium sp. | Fusarium oxysporum | A | IV | 35% | [106] |
| 4-Prenyloxyclavatol | Nigrospora sphaerica | Colletotrichum gloeosporioides | A | IV | 63% | [107] |
| 5-Methylmellein | Biscogniauxia mediterranea | Phomopsis obscurans Phomopsis viticola | A | IV | 5% | [108] |
| 5-(Undeca-3′,5′,7′-trien-1′-yl) furan-2-ol | Emericella sp. | Verticillium dahliae | A | IV | 49% | [109] |
| 5-(Undeca-3′,5′,7′-trien-1′-yl) furan-2-carbonate | 12% | |||||
| Bicolorin D | Saccharicola bicolor | Sclerotinia sclerotiorum | A | IV | 13% | [110] |
| IP-A | 57% | |||||
| Brefeldin A | Cladosporium sp. | Aspergillus niger | A | IV | 8-fold | [111] |
| Cercosporamide | Cadophora orchidicola | Fusarium oxysporum Pestalotia diospyri Botrytis cinerea Sclerotium rolfsii Penicillium digitatum | A | IV | - | [112] |
| Cuminic acid | Aspergillus spp. | Phytophthora spp. | A | IV | - | [113] |
| Epirodin | Epicoccum nigrum | Brotrytis cinerea | A | IV, IP-A | - | [114] |
| Ergot alkaloids | Claviceps purpurea | - | A | - | - | [96] |
| Exserolide C | Exserohilum sp. | Fusarium oxysporum | A | IV | 3% | [104] |
| Ferric chloride, potassium hydroxide, vanillin–sulfuric acid | Xylaria allantoidea | - | A | IV | - | [103] |
| Geoxantethers A and B | Fungus from Massarinaceae | Microbotryum violaceum | A | IV | 16% | [115] |
| Griseofulvin | Penicillium sp. | Botrytis cinerea | A | IV | - | [95] |
| Guignardianone C | Phyllosticta sp. | Botrytis cinerea | A | IV | 52% | [96] |
| Hexadecanoic acid, 2,3-bis ((trimethylsilyl) oxy) propyl ester | Trichoderma harzanium | Sclerotinia sclerotiorum | A | IV | 25% | [116] |
| Leucinostatins A and B | Purpureocillium lilacinum | Phytophthora sp. | A | IV | - | [117] |
| Macrosporin | Phoma sp. | Fusarium graminearum | A | IV | 10% | [118] |
| Methyl dichloroasterrate | Aspergillus capensis | Botrytis cinerea Monilinia fructicola Sclerotinia sclerotiorum Sclerotinia trifoliorum | A, ISR | IV | 17% | [119] |
| Monocerin | Drechslera sp. | Botrytis cinerea Sclerotinia sclerotiorum | A | IV, IP-A | - | [120] |
| Nigrosporamide A | Nigrospora sphaerica | Colletotrichum gloeosporioides | A | IV | 10.83-fold | [107] |
| Not identified | Metarhizium anisopliae | Fusarium graminearum Fusarium oxysporum | A | IV | - | [121] |
| Not identified | Metarhizium anisopliae | Fusarium graminearum | A | IP-F | - | [53] |
| Palmitic acid, stearic acid, octadecenoic acid | Fusarium oxysporum | - | A, ISR, PGP | IV | - | [82] |
| Penicillither | Aspergillus capensis | Botrytis cinerea Monilinia fructicola Sclerotinia sclerotiorum Sclerotinia trifoliorum | A | IV | 5% | [119] |
| Penochalasin K | Penicillium chrysogenum, | Colletotrichum gloeosporioides | A | IV | 10-fold | [122] |
| Rhizoctonia solani | 2.66-fold | |||||
| Pestalachlorides A, B and C | Pestalotiopsis adusta | Fusarium culmorum Gibberella zeae Verticillium aiboatrum | A | IV | - | [123] |
| Pretrichodermamide A | Trichoderma harzianum Epiccocum nigrum | Ustilago maydis | A | IV | 2-fold | [124] |
| Pseudoanguillosporin A | Pseudoanguillospora sp. | Mycrobotryum violaceum | A | IV | 40% | [125] |
| Pyrenophorol | Lophodermium nitens | Cronartium ribicola | A | IP-G | - | [126] |
| Rosellichalasin | Aspergillus capensis | Botrytis cinerea Monilinia fructicola Sclerotinia sclerotiorum Sclerotinia trifoliorum | A | IV | 32% | [119] |
| Speciosin U | Saccharicola sp. | Cladosporium cladosporioides | A | IV | - | [127] |
| Sporothriolide | Nodulisporium sp. | Rhizoctonia solani Sclerotinia sclerotiorum | A | IV | 60% | [128] |
| IP-G | 79% | |||||
| Trichodermin | Trichoderma brevicompactum | Rhizoctonia solani | A | IV | 1.44-fold | [129] |
| Fusarium solani | 1.21-fold | |||||
| Versicolorin B | Aspergillus versicolor | Colletotrichum musae | A | IV | - | [130] |
| Viriditoxin | Byssochlamys spectabilis | Fusarium spp. | A | IV; IP-G | - | [20] |
5. Current and Future Perspectives for a Large-Scale Application
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moore, D.; Robson, G.D.; Trinci, A.P.J. 21st Century Guidebook to Fungi, 2nd ed.; Cambridge University Press: Cambridge, UK, 2020; pp. 1–610. [Google Scholar]
- Loo, J.A. Ecological impacts of non-native invertebrates and fungi on terrestrial ecosystems. Biol. Invasions 2009, 11, 81–96. [Google Scholar] [CrossRef]
- Mostafalou, S.; Abdollahi, M. Pesticides and human chronic diseases: Evidences, mechanisms, and perspectives. Toxicol. Appl. Pharmacol. 2013, 268, 157–177. [Google Scholar] [CrossRef] [PubMed]
- Pearson, B.L.; Simon, J.M.; McCoy, E.S.; Salazar, G.; Fragola, G.; Zylka, M.J. Identification of chemicals that mimic transcriptional changes associated with autism, brain aging and neurodegeneration. Nat. Commun. 2016, 7, 11173. [Google Scholar] [CrossRef] [PubMed]
- Howell, C.C.; Semple, K.T.; Bending, G.D. Isolation and characterisation of azoxystrobin degrading bacteria from soil. Chemosphere 2014, 95, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Leitão, S.; Cerejeira, M.J.; Van den Brink, P.J.; Sousa, J.P. Effects of azoxystrobin, chlorothalonil, and ethoprophos on the reproduction of three terrestrial invertebrates using a natural Mediterranean soil. Appl. Soil Ecol. 2014, 76, 124–131. [Google Scholar] [CrossRef]
- Warming, T.P.; Mulderij, G.; Christoffersen, K.S. Clonal variation in physiological responses of Daphnia magna to the strobilurin fungicide azoxystrobin. Environ. Toxicol. Chem. 2009, 28, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, E.T.; Lopes, I.; Pardal, M.A. Occurrence, fate and effects of azoxystrobin in aquatic ecosystems: A review. Environ. Int. 2013, 53, 18–28. [Google Scholar] [CrossRef]
- Tomlin, C.D.S. The Pesticide Manual; British Crop Protection Council: Farnham, UK, 2000. [Google Scholar]
- Leroch, M.; Kretschmer, M.; Hahn, M. Fungicide resistance phenotypes of Botrytis cinerea isolates from commercial vineyards in South West Germany. J. Phytopathol. 2011, 159, 63–65. [Google Scholar] [CrossRef]
- El-Baky, N.A.; Amara, A.A.A.F. Recent approaches towards control of fungal diseases in plants: An updated review. J. Fungi 2021, 7, 900. [Google Scholar] [CrossRef]
- Leplat, J.; Friberg, H.; Abid, M.; Steinberg, C. Survival of Fusarium graminearum, the causal agent of Fusarium head blight. A review. Agron. Sustain. Dev. 2013, 33, 97–111. [Google Scholar] [CrossRef]
- Feichtenberger, E. Citrus phytophthora diseases: Management challenges and successes. J. Citrus Pathol. 2015, 2, 27203. [Google Scholar] [CrossRef]
- Rahman, M.; Islam, T.; Jett, L.; Kotcon, J. Biocontrol agent, biofumigation, and grafting with resistant rootstock suppress soil-borne disease and improve yield of tomato in West Virginia. Crop Prot. 2021, 145, 105630. [Google Scholar] [CrossRef]
- Romeralo, C.; Santamaría, O.; Pando, V.; Diez, J.J. Fungal endophytes reduce necrosis length produced by Gremmeniella abietina in Pinus halepensis seedlings. Biol. Control 2015, 80, 30–39. [Google Scholar] [CrossRef]
- Rebek, E.J.; Frank, S.D.; Royer, T.A.; Bográn, C.E. Alternatives to chemical control of insect pests. In Insecticides—Basic and Other Applications; Solonesky, S., Larramendy, M.L., Eds.; InTech: London, UK, 2012; pp. 171–196. [Google Scholar] [CrossRef]
- Dong, C.; Wang, L.; Li, Q.; Shang, Q. Epiphytic and endophytic fungal communities of tomato plants. Hortic. Plant J. 2021, 7, 38–48. [Google Scholar] [CrossRef]
- Muller, T.; Ruppel, S. Progress in cultivation-independent phyllosphere microbiology. FEMS Microbiol. Ecol. 2014, 87, 2–17. [Google Scholar] [CrossRef]
- Mota, S.F.; Padua, P.F.; Ferreira, A.N.; Gomes, L.D.W.; Dias, M.A.; Souza, E.A.; Pereira, O.L.; Cardoso, P.G. Biological control of common bean diseases using endophytic Induratia spp. Biol. Control 2021, 159, 104629. [Google Scholar] [CrossRef]
- Rodrigo, S.; Santamaría, O.; Halecker, S.; Lledó, S.; Stadler, M. Antagonism between Byssochlamys spectabilis (anamorph Paecilomyces variotii) and plant pathogens: Involvement of the bioactive compounds produced by the endophyte. Ann. Appl. Biol. 2017, 171, 464–476. [Google Scholar] [CrossRef]
- Hardoim, P.R.; van Overbeek, L.S.; Berg, G.; Pirttilä, A.M.; Compant, S.; Campisano, A.; Döring, M.; Sessitsch, A. The hidden world within plants: Ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015, 79, 293–320. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Jin, Q.; Peng, H.; Zhu, T.; Ye, H. Effect of a fungus, Hypoxylon spp., on endophytes in the roots of Asparagus. FEMS Microbiol. Lett. 2019, 366, fnz207. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.E.; Maynard, Z.; Gilbert, G.S.; Coley, P.D.; Kursar, T.A. Are tropical fungal endophytes hyperdiverse? Ecol. Lett. 2000, 3, 267–274. [Google Scholar] [CrossRef]
- Saikkonen, K.; Ahlholm, J.; Helander, M.; Lehtimäki, S.; Niemeläinen, O. Endophytic fungi in wild and cultivated grasses in Finland. Ecography 2000, 23, 360–366. [Google Scholar] [CrossRef]
- Lledó, S.; Rodrigo, S.; Poblaciones, M.J.; Santamaria, O. Biomass yield, nutritive value and accumulation of minerals in Trifolium subterraneum L. as affected by fungal endophytes. Plant Soil 2016, 405, 197–210. [Google Scholar] [CrossRef]
- Whithe, J.F.; Kingsley, K.L.; Verma, S.K.; Kowalski, K.P. Rhizophagycycle: An oxidative process in plants for nutrient extraction from symbiotic microbes. Microorganisms 2018, 6, 95. [Google Scholar] [CrossRef] [PubMed]
- García-Latorre, C.; Rodrigo, S.; Santamaría, O. Endophytes as plant nutrient uptake-promoter in plants. In Endophytes: Mineral Nutrient Management, Volume 3. Sustainable Development and Biodiversity; Maheshwari, D.K., Dheeman, S., Eds.; Springer: Cham, Switzerland, 2021; Volume 26, pp. 247–265. [Google Scholar] [CrossRef]
- García-Latorre, C.; Rodrigo, S.; Santamaria, O. Effect of fungal endophytes on plant growth and nutrient uptake in Trifolium subterraneum and Poa pratensis as affected by plant host specificity. Mycol. Progress 2021, 20, 1217–1231. [Google Scholar] [CrossRef]
- Vázquez-de-Aldana, B.R.; Zabalgogeazcoa, I.; García-Ciudad, A.; García-Criado, B. An Epichloë endophyte affects the competitive ability of Festuca rubra against other grassland species. Plant Soil 2013, 362, 201–213. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Macias-Rodriguez, L.; Cortes-Penagos, C.; Lopez-Bucio, J. Trichoderma virens, a plant beneficial fungus, enhances biomass production and promotes lateral root growth through an auxin-dependent mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef]
- Spiering, M.J.; Greer, D.H.; Schmid, J. Effects of the fungal endophyte, Neotyphodium lolii, on net photosynthesis and growth rates of perennial ryegrass (Lolium perenne) are independent of in planta endophyte concentration. Ann. Bot. 2006, 98, 379–387. [Google Scholar] [CrossRef]
- Hamilton, C.E.; Bauerle, T.L. A new currency for mutualism? Fungal endophytes alter antioxidant activity in hosts responding to drought. Fungal Divers. 2012, 54, 39–49. [Google Scholar] [CrossRef]
- Ghabooli, M.; Khatabi, B.; Ahmadi, F.S.; Sepehri, M.; Mirzaei, M.; Amirkhani, A.; Jorrín-Novo, J.V.; Salekdeh, G.H. Proteomics study reveals the molecular mechanisms underlying water stress tolerance induced by Piriformospora indica in barley. J. Proteomics 2013, 94, 289–301. [Google Scholar] [CrossRef]
- Zhou, X.R.; Dai, L.; Xu, G.F.; Shen, H. A strain of Phoma species improves drought tolerance of Pinus tabulaeformis. Sci. Rep. 2021, 11, 7637. [Google Scholar] [CrossRef]
- Ismail, I.; Hamayun, M.; Hussain, A.; Iqbal, A.; Khan, S.A.; Khan, M.A.; Lee, I.J. An endophytic fungus Gliocladium cibotii regulates metabolic and antioxidant system of glycine max and helianthus annuus under heat stress. Pol. J. Environ. Stud. 2021, 30, 1631–1640. [Google Scholar] [CrossRef]
- Zamani, N.; Sabzalian, M.R.; Khoshgoftarmanesh, A.; Afyuni, M. Neotyphodium endophyte changes phytoextraction of zinc in Festuca arundinacea and Lolium perenne. Int. J. Phytoremediat. 2015, 17, 456–463. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Schillaci, M.; Walker, R.; Smith, P.M.C.; Watt, M.; Roessner, U. Alleviation of salinity stress in plants by endophytic plant-fungal symbiosis: Current knowledge, perspectives and future directions. Plant Soil 2021, 461, 219–244. [Google Scholar] [CrossRef]
- Moghaddam, M.S.H.; Safaie, N.; Soltani, J.; Hagh-Doust, N. Desert-adapted fungal endophytes induce salinity and drought stress resistance in model crops. Plant Physiol. Biochem. 2021, 160, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Ważny, R.; Rozpądek, P.; Domka, A.; Jędrzejczyk, R.J.; Nosek, M.; Hubalewska-Mazgaj, M.; Lichtscheidl, I.; Kidd, P.; Turnau, K. The effect of endophytic fungi on growth and nickel accumulation in Noccaea hyperaccumulators. Sci. Total Environ. 2021, 768, 144666. [Google Scholar] [CrossRef]
- Clay, K.; Schardl, C. Evolutionary origins and ecological consequences of endophyte symbiosis with grasses. Am. Nat. 2002, 160, S99–S127. [Google Scholar] [CrossRef] [PubMed]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Verma, V.C.; Wheeler, T.A.; Woodward, J.E.; Starr, J.L.; Sword, G.A. Tapping into the cotton fungal phytobiome for novel nematode biological control tools. Phytobiomes J. 2020, 4, 19–26. [Google Scholar] [CrossRef]
- Kumar, K.K.; Dara, S.K. Fungal and bacterial endophytes as microbial control agents for plant-parasitic nematodes. Int. J. Environ. Res. Public. Health 2021, 18, 4269. [Google Scholar] [CrossRef]
- Hartley, S.; Gange, A. Impacts of plant symbiotic fungi on insect herbivores: Mutualism in a multitrophic context. Annu. Rev. Entomol. 2009, 54, 323–342. [Google Scholar] [CrossRef]
- García Parisi, P.A.; Grimoldi, A.A.; Omacini, M. Endophytic fungi of grasses protect other plants from aphid herbivory. Fungal Ecol. 2014, 9, 61–64. [Google Scholar] [CrossRef]
- Ramakuwela, T.; Hatting, J.; Bock, C.; Vega, F.E.; Wells, L.; Mbata, G.N.; Shapiro-Ilan, D. Establishment of Beauveria bassiana as a fungal endophyte in pecan (Carya illinoinensis) seedlings and its virulence against pecan insect pests. Biol. Control 2020, 140, 104102. [Google Scholar] [CrossRef]
- Santamaría, O.; González, M.A.; Pajares, J.A.; Diez, J.J. Effect of fungicides, endophytes and fungal filtrates on in vitro growth of Spanish isolates of Gremmeniella abietina. For. Pathol. 2007, 37, 251–262. [Google Scholar] [CrossRef]
- Farhat, H.; Urooj, F.; Tariq, A.; Sultana, V.; Ansari, M.; Ahmad, V.U.; Ehteshamul-Haque, S. Evaluation of antimicrobial potential of endophytic fungi associated with healthy plants and characterization of compounds produced by endophytic Cephalosporium and Fusarium solani. Biocatal. Agric. Biotechnol. 2019, 18, 101043. [Google Scholar] [CrossRef]
- Rojas, E.C.; Jensen, B.; Jørgensen, H.J.L.; Latz, M.A.C.; Esteban, P.; Ding, Y.; Collinge, D.B. Selection of fungal endophytes with biocontrol potential against Fusarium head blight in wheat. Biol. Control 2020, 144, 104222. [Google Scholar] [CrossRef]
- Silva-Valderrama, I.; Toapanta, D.; de los Angeles Miccono, M.; Lolas, M.; Díaz, G.A.; Cantu, D.; Castro, A. Biocontrol potential of grapevine endophytic and rhizospheric fungi against trunk pathogens. Front. Microbiol. 2021, 11, 3311. [Google Scholar] [CrossRef]
- Santamaria, O.; Smith, D.R.; Stanosz, G.R. Interaction between Diplodia pinea or Diplodia scrobiculata and fungal endophytes isolated from pine shoots. Can. J. For. Res. 2012, 42, 1819–1826. [Google Scholar] [CrossRef]
- Halecker, S.; Wennrich, J.P.; Rodrigo, S.; Andrée, N.; Rabsch, L.; Baschien, C.; Steinert, M.; Stadler, M.; Surup, F.; Schulz, B. Fungal endophytes for biocontrol of ash dieback: The antagonistic potential of Hypoxylon rubiginosum. Fungal Ecol. 2020, 45, 100918. [Google Scholar] [CrossRef]
- Hao, Q.; Albaghdady, D.M.D.; Xiao, Y.; Xiao, X.; Mo, C.; Tian, T.; Wang, G. Endophytic Metarhizium anisopliae is a potential biocontrol agent against wheat Fusarium head blight caused by Fusarium graminearum. J. Plant Pathol. 2021, 103, 875–885. [Google Scholar] [CrossRef]
- Busby, P.E.; Ridout, M.; Newcombe, G. Fungal endophytes: Modifiers of plant disease. Plant Mol. Biol. 2016, 90, 645–655. [Google Scholar] [CrossRef]
- Zivanovic, A.; Rodgers, L. The role of fungal endophytes in plant pathogen resistance. Bios 2018, 89, 192–197. [Google Scholar] [CrossRef]
- Pavithra, G.; Sumant, B.; Meenakshi, R.; Seweta, S. Role of endophytic microbes against plant pathogens: A review. Asian J. Plant Sci. 2020, 19, 54–62. [Google Scholar] [CrossRef]
- Gao, F.-K.; Dai, C.-C.; Liu, X.-Z. Mechanisms of fungal endophytes in plant protection against pathogens. Afr. J. Microbiol. Res. 2010, 4, 1346–1351. [Google Scholar] [CrossRef]
- Zabalgogeazcoa, I. Fungal endophytes and their interaction with plant pathogens. Span. J. Agric. Res. 2008, 6, 138–146. [Google Scholar] [CrossRef]
- Elad, Y. Mechanisms involved in the biological control of Botrytis cinerea incited diseases. Eur. J. Plant Pathol. 1996, 102, 719–732. [Google Scholar] [CrossRef]
- Bailey, B.; Bae, H.; Strem, M.; Crozier, J.; Thomas, S.; Samuels, G.; Vinyard, B.; Holmes, K. Antibiosis, mycoparasitism, and colonization success for endophytic Trichoderma isolates with biological control potential in Theobroma cacao. Biol. Control. 2008, 46, 24–35. [Google Scholar] [CrossRef]
- Zeilinger, S.; Galhaup, C.; Payer, K.; Woo, S.L.; Mach, R.L.; Fekete, C.; Lorito, M.; Kubicek, C.P. Chitinase gene expression during mycoparasitic interaction of Trichoderma harzianum with its host. Fungal Genet. Biol. 1999, 26, 131–140. [Google Scholar] [CrossRef]
- Harman, G.; Howell, C.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef]
- Molitor, A.; Zajic, D.; Voll, L.M.; Pons-Kühnemann, J.; Samans, B.; Kogel, K.-H.; Waller, F. Barley leaf transcriptome and metabolite analysis reveals new aspects of compatibility and Piriformospora indicamediated systemic induced resistance to powdery mildew. Mol. Plant Microbe Interact. 2011, 24, 1427–1439. [Google Scholar] [CrossRef]
- Nassimi, Z.; Taheri, P. Endophytic fungus Piriformospora indica induced systemic resistance against rice sheath blight via affecting hydrogen peroxide and antioxidants. Biocontrol Sci. Technol. 2017, 3157, 256–267. [Google Scholar] [CrossRef]
- Shoresh, M.; Harman, G.E.; Mastouri, F. Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol. 2010, 48, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; He, Q.; Chen, C.; Zhang, C. Differential communications between fungi and host plants revealed by secretome analysis of phylogenetically related endophytic and pathogenic fungi. PLoS ONE 2016, 11, e0163368. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.K.M.; Tellier, A. Plant-parasite coevolution: Bridging the gap between genetics and ecology. Annu. Rev. Phytopathol. 2011, 49, 345–367. [Google Scholar] [CrossRef] [PubMed]
- Lubna; Asaf, S.; Hamayun, M.; Gul, H.; Lee, I.J.; Hussain, A. Aspergillus niger CSR3 regulates plant endogenous hormones and secondary metabolites by producing gibberellins and indoleacetic acid. J. Plant Interact. 2018, 13, 100–111. [Google Scholar] [CrossRef]
- Clarke, B.B.; White, J.F.; Hurley, H.; Torres, M.S.; Sun, S.; Huff, D.R. Endophyte-mediated suppression of dollar spot disease in fine fescues. Plant Dis. 2006, 90, 994–998. [Google Scholar] [CrossRef]
- Istifadah, N.; McGee, P.A. Endophytic Chaetomium globosum reduces development of tan spot in wheat caused by Pyrenophora tritici-repentis. Australas. Plant Pathol. 2006, 35, 411–418. [Google Scholar] [CrossRef]
- Tan, R.X.; Zou, W.X. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef]
- Schulz, B.; Boyle, C.; Draeger, S.; Römmert, A.K.; Krohn, K. Endophytic fungi: A source of novel biologically active secondary metabolites. Mycol. Res. 2002, 106, 996–1004. [Google Scholar] [CrossRef]
- Mousa, W.K.; Raizada, M.N. The diversity of anti-microbial secondary metabolites produced by fungal endophytes: An interdisciplinary perspective. Front. Microbiol. 2013, 4, 65. [Google Scholar] [CrossRef]
- Sahoo, S.; Subban, K.; Chelliah, J. Diversity of marine macro-algicolous endophytic fungi and cytototix potential of Biscogniauxia petrensis metabolites against cancer cell lines. Front. Microbiol. 2021, 12, 650177. [Google Scholar] [CrossRef]
- Adhila Haji Basheerudeen, M.; Mushtaq, S.A.; Soundhararajan, R.; Kumar Nachimuthu, S.; Srinivasan, H. Marine endophytic fungi mediated Silver nanoparticles and their application in plant growth promotion in Vigna radiata L. Int. J. Nano Dimens. 2021, 12, 1–10. [Google Scholar]
- Taritla, S.; Jumari, M.; Kamat, S.; Bhat, S.G.; Jayabaskaran, C. Optimization of physicochemical parameters for production of cytotoxic secondary metabolites and apoptosis induction activities in the culture extract of a marine algal-derived endophytic fungus Aspergillus sp. Front. Pharmacol. 2021, 12, 542891. [Google Scholar] [CrossRef]
- Quader, M.M.; Hamed, A.A.; Soldatou, S.; Abdelraof, M.; Elawady, M.E.; Hassane, A.S.I.; Belbahri, L.; Ebel, R.; Rateb, M.E. Antimicrobial and antibiofilm activities of the fungal metaboiltes isolated form the marine endophytes Epicoccum nigrum M13 and Alternaria alternata 13A. Mar. Drugs 2021, 19, 232. [Google Scholar] [CrossRef] [PubMed]
- Savidov, N.; Gloriozov, T.A.; Poroikov, V.V.; Dembitsky, V.M. Highly oxygenated isoprenoid lipids derived from fungi and fungal endophytes: Origin and biological activities. Steroids 2018, 140, 114–124. [Google Scholar] [CrossRef]
- Kharwar, R.N.; Mishra, A.; Gond, S.K. Anticancer compounds derived from fungal endophytes: Their importance and future challenges. Nat. Prod. Rep. 2011, 28, 1208–1228. [Google Scholar] [CrossRef]
- Chandra, S. Endophytic fungi: Novel sources of anticancer lead molecules. Appl. Microbiol. Biotechnol. 2012, 95, 47–59. [Google Scholar] [CrossRef]
- Mazumder, K.; Ruma, Y.N.; Akter, R.; Aktar, A.; Hossain, M.M.; Shahiina, Z.; Mazumdar, S.; Kerr, P.G. Identification of bioactive metabolites and evaluation of in vitro anti-inflammatory and in vivo antinociceptive and antiarthritic activities of endophyte fungi isolated from Elaeocarpus floribundus blume. J. Ethnopharmacol. 2021, 273, 113975. [Google Scholar] [CrossRef]
- Kiliç, G.; Tosun, G.; Bozdeveci, A.; Erik, I.; Öztürk, E.; Reis, R.; Sipahi, H.; Cora, M.; Karaoğlu, S.A.; Yaylı, N. Antimicrobial, cytotoxic, antiviral effects, and spectroscopic characterization of metabolites produced by Fusarium oxysporum YP9B. Rec. Nat. Prod. 2021, 15, 547–567. [Google Scholar] [CrossRef]
- Toghueo, R.M.K.; Zabalgogeazcoa, I.; Vázquez de Aldana, B.R.; Boyom, F.F. Enzymatic activity of endophytic fungi from the medicinal plants Terminalia catappa, Terminalia mantaly and Cananga odorata. S. Afr. J. Bot. 2017, 109, 146–153. [Google Scholar] [CrossRef]
- Vukelic, I.D.; Prokic, L.T.; Racic, G.M.; Pešic, M.B.; Bojovic, M.M.; Sierka, E.M.; Kalaji, H.M.; Pankovic, D.M. Effects of Trichoderma harzianum on photosynthetic characteristics and fruit quality of tomato plants. Int. J. Mol. Sci. 2021, 22, 6961. [Google Scholar] [CrossRef] [PubMed]
- Palanichamy, P.; Krishnamoorthy, G.; Kannan, S.; Marudhamuthu, M. Bioactive potential of secondary metabolites derived from medicinal plant endophytes. Egypt. J. Basic Appl. Sci. 2018, 5, 303–312. [Google Scholar] [CrossRef]
- Palupi, K.D.; Ilyas, M.; Agusta, A. Endophytic fungi inhabiting Physalis angulata L. plant: Diversity, antioxidant, and antibacterial activities of their ethyl acetate extracts. J. Basic Clin. Physiol. Pharmacol. 2021, 32, 823–829. [Google Scholar] [CrossRef]
- Kumaresan, S.; Karthi, V.; Senthilkumar, V.; Balakumar, B.S.; Stephen, A. Biochemical constituents and antioxidant potential of endophytic fungi isolated from the leaves of Azadirachta indica A. Juss (neem) from Chennai, India. J. Acad. Ind. Res. 2015, 3, 355–361. [Google Scholar]
- Bauerfeind, J.C. Carotenoids as Colorants and Vitamin A Precursors; Academic Press: New York, NY, USA, 1981. [Google Scholar]
- Marie, C.F.; Regine, M.Q.; Stephane, W.; Emilie, T.; Joseph, R.; Brigitte, W.R.; Edmond, R.; Marie, P.V. Immune status is more affected by age than by carotenoid depletionrepletion in healthy human subjects. Br. J. Nutr. 2012, 108, 2054–2065. [Google Scholar] [CrossRef]
- Hwang, J.S.; You, Y.H.; Bae, J.J.; Khan, S.A.; Kim, J.G.; Choo, Y.S. Effects of endophytic fungal secondary metabolites on growth and physiological response of Carex kobomugi. J. Coast. Res. 2011, 27, 544–548. [Google Scholar] [CrossRef]
- Chen, Q.; Li, Z.L.; Qu, Z.M.; Zhou, H.Y.; Qi, Y.J.; Liu, Z.G.; Zhang, M. Maize yield and root morphological characteristics affected by controlled release diammonium phosphate and Paecilomyces variotii extracts. Field Crops Res. 2020, 255, 107862. [Google Scholar] [CrossRef]
- Wang, X.; Yao, Y.; Chen, B.; Zhang, M.; Liu, Z.; Wang, Q.; Ma, J. Paecilomyces variotii extracts and controlled-release urea synergistically increased nitrogen use efficiency and rice yield. ACS Omega 2020, 22, 13303–13311. [Google Scholar] [CrossRef] [PubMed]
- Zheng, R.; Li, S.; Zhang, X.; Zhao, C. Biological activities of some new secondary metabolites isolated from endophytic fungi: A review study. Int. J. Mol. Sci. 2021, 22, 959. [Google Scholar] [CrossRef]
- Xu, K.; Li, X.-Q.; Zhao, D.-L.; Zhang, P. Antifungal secondary metabolites produced by the fungal endophytes: Chemical diversity and potential use in the development of biopesticides. Front. Microbiol. 2021, 12, 689527. [Google Scholar] [CrossRef]
- Huber, F.M.; Gottlieb, D. The mechanism of action of griseofulvin. Can. J. Microbiol. 1968, 14, 111–118. [Google Scholar] [CrossRef]
- Dembitsky, V.M. Naturally occurring bioactive Cyclobutane-containing (CBC) alkaloids in fungi, fungal endophytes, and plants. Phytomedicine 2014, 21, 1559–1581. [Google Scholar] [CrossRef]
- Sergeiko, A.; Poroikov, V.V.; Hanuš, L.O.; Dembitsky, V.M. Cyclobutane-containing alkaloids: Origin, synthesis, and biological activities. Open Med. Chem. J. 2008, 2, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Bladt, T.T.; Frisvad, J.C.; Knudsen, P.B.; Larsen, T.O. Anticancer and antifungal compounds from Aspergillus, Penicillium and other filamentous fungi. Molecules 2013, 18, 11338–11376. [Google Scholar] [CrossRef] [PubMed]
- Kuklev, D.V.; Domb, A.J.; Dembitsky, V.M. Bioactive acetylenic metabolites. Phytomedicine 2013, 20, 1145–1159. [Google Scholar] [CrossRef]
- Seigler, D.S. Acetylenic compounds. In Plant Secondary Metabolism; Seigler, D.S., Ed.; Springer: Boston, MA, USA, 1998; pp. 42–50. [Google Scholar] [CrossRef]
- Azerang, P.; Sardari, S. Antifungal activity of enynediesters and acetylenic compounds obtained by synthesis and in silico prediction pattern. J. Mycol. Med. 2012, 22, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Tripathi, S.K.; Feng, Q.; Lorenz, M.C.; Wright, M.A.; Jacob, M.R.; Mask, M.M.; Baerson, S.R.; Li, X.C.; Clark, A.M.; et al. A potent plant-derived antifungal acetylenic acid mediates its activity by interfering with fatty acid homeostasis. Antimicrob. Agents Chemother. 2012, 56, 2894–2907. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, G.D.; Gomes, R.R.; Gonçalves, R.; Fornari, G.; Maia, B.H.L.N.S.; Schmidt-Dannert, C.; Gaascht, F.; Glienke, G.; Schneider, G.X.; Colombo, I.R.; et al. Molecular identification and antimicrobial activity of foliar endophytic fungi on the Brazilian pepper tree (Schinus terebinthifolius) reveal new species of Diaporthe. Curr. Microbiol. 2021, 78, 3218–3229. [Google Scholar] [CrossRef]
- Li, R.; Chen, R.; Niu, S.; Guo, L.; Yin, J.; Che, Y. Exserolides A–F, new isocoumarin derivatives from the plant endophytic fungus Exserohilum sp. Fitoterapia 2014, 96, 88–94. [Google Scholar] [CrossRef]
- Yang, X.F.; Wang, N.N.; Kang, Y.F.; Ma, Y.M. A new furan derivative from an endophytic Aspergillus tubingensis of Decaisnea insignis (Griff.) Hook.f. & Thomson. Nat. Prod. Res. 2019, 33, 2777–2783. [Google Scholar] [CrossRef]
- Elkhayat, E.S.; Goda, A.M. Antifungal and cytotoxic constituents from the endophytic fungus Penicillium sp. Bull. Fac. Pharmacy Cairo Univ. 2017, 55, 85–89. [Google Scholar] [CrossRef]
- Zhu, X.; Chen, J.; Zhu, S.; He, Y.; Ding, W.; Li, C. Two new compounds from Nigrospora sphaerica ZMT05, a fungus derivated from Oxya chinensis Thunber. Nat. Prod. Res. 2017, 32, 2375–2381. [Google Scholar] [CrossRef]
- Silva-Hughes, A.F.; Wedge, D.E.; Cantrell, C.L.; Carvalho, C.R.; Pan, Z.; Moraes, R.M.; Madoxx, V.L.; Rosa, L.H. Diversity and antifungal activity of the endophytic fungi associated with the native medicinal cactus Opuntia humifusa (Cactaceae) from the United States. Microbiol. Res. 2015, 175, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Pang, X.J.; Xu, L.L.; Zhao, T.; Long, X.Y.; Zhang, Q.Y.; Qin, H.L.; Yang, D.F.; Yang, X.L. Two new alkylated furan derivatives with antifungal and antibacterial activities from the plant endophytic fungus Emericella sp. XL029. Nat. Prod. Res. 2018, 32, 2625–2631. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Guo, D.L.; Liu, G.H.; Fu, X.; Gu, Y.G.; Ding, L.S.; Zhou, Y. Antifungal halogenated cyclopentenones from the endophytic fungus Saccharicola bicolor of Bergenia purpurascens by the one strain-many compounds strategy. J. Agric. Food Chem. 2020, 68, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.W.; Jiao, R.H.; Cheng, A.B.; Tan, S.H.; Song, Y.C. Antimicrobial potentials of endophytic fungi residing in Quercus variabilis and brefeldin A obtained from Cladosporium sp. World J. Microbiol. Biotechnol. 2007, 23, 79–83. [Google Scholar] [CrossRef]
- Wang, L.; Shen, J.; Xu, L.; Gao, J.; Zhang, C.; Wang, Y.; Chen, F. A metabolite of endophytic fungus Cadophora orchidicola from Kalimeris indica serves as a potential fungicide and TLR4 agonist. J. Appl. Microbiol. 2019, 126, 1383–1390. [Google Scholar] [CrossRef]
- Ye, Y.; Chen, Y.; Hou, Y.; Yu, H.; Zhu, L.; Sun, Y.; Zhou, M.; Chen, Y.; Dong, M. Two new benzoic acid derivatives from endophytic fungus Aspergillus versicolor. Nat. Prod. Res. 2020, Online ahead of print. [CrossRef]
- Alcock, A.; Elmer, P.; Marsden, R.; Parry, F. Inhibition of Botrytis cinerea by Epirodin: A secondary metabolite from New Zealand isolates of Epicoccum nigrum. J. Phytopathol. 2015, 163, 841–852. [Google Scholar] [CrossRef]
- Richardson, S.N.; Nsiama, T.K.; Walker, A.K.; McMullin, D.R.; Miller, J.D. Antimicrobial dihydrobenzofurans and xanthenes from a foliar endophyte of Pinus strobus. Phytochemistry 2015, 117, 436–443. [Google Scholar] [CrossRef]
- Chowdhary, K.; Sharma, S. Plant growth promotion and biocontrol potential of fungal endophytes in the inflorescence of Aloe vera L. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2020, 90, 1045–1055. [Google Scholar] [CrossRef]
- Wang, G.; Liu, Z.; Lin, R.; Li, E.; Mao, Z.; Ling, J.; Yang, Y.; Yin, W.B.; Xie, B. Biosynthesis of antibiotic leucinostatins in bio-control fungus Purpureocillium lilacinum and their inhibition on Phytophthora revealed by genome mining. PLoS Pathog. 2016, 12, e1005685. [Google Scholar] [CrossRef]
- Huang, S.; Xu, J.; Li, F.; Zhou, D.; Xu, L.; Li, C. Identification and antifungal activity of metabolites from the mangrove fungus Phoma sp. L28. Chem. Nat. Compd. 2017, 53, 237–240. [Google Scholar] [CrossRef]
- Qin, J.; Lyu, A.; Zhang, Q.H.; Yang, L.; Zhang, J.; Wu, M.D.; Li, G.Q. Strain identification and metabolites isolation of Aspergillus capensis CanS-34A from Brassica napus. Mol. Biol. Rep. 2019, 46, 3451–3460. [Google Scholar] [CrossRef]
- d’Errico, G.; Aloj, V.; Flematti, G.R.; Sivasithamparam, K.; Worth, C.M.; Lombardi, N.; Ritieni, A.; Marra, R.; Lorito, M.; Vinale, F. Metabolites of a Drechslera sp. endophyte with potential as biocontrol and bioremediation agent. Nat. Prod. Res. 2021, 35, 4508–4516. [Google Scholar] [CrossRef]
- Ravindran, K.; Chitra, S.; Wilson, A.; Sivaramakrishnan, S. Evaluation of antifungal activity of Metarhizium anisopliae against plant phytopathogenic fungi. In Microbial Diversity and Biotechnology in Food Security; Kharwar, R.N., Upadhyay, R.S., Dubey, N.K., Raghuwanshi, R., Eds.; Springer: New Delhi, India, 2014; pp. 251–255. [Google Scholar] [CrossRef]
- Zhu, X.; Zhou, D.; Liang, F.; Wu, Z.; Shi, Z.; Li, C. Penochalasin K, a new unusual chaetoglobosin from the mangrove endophytic fungus Penicillium chrysogenum V11 and its effective semisynthesis. Fitoterapia 2017, 123, 23–28. [Google Scholar] [CrossRef]
- Li, E.; Jiang, L.; Guo, L.; Zhang, H.; Che, Y. Pestalachlorides A–C, antifungal metabolites from the plant endophytic fungus Pestalotiopsis adusta. Bioorg. Med. Chem. 2008, 17, 7894–7899. [Google Scholar] [CrossRef] [PubMed]
- Harwoko, H.; Daletos, G.; Stuhldreier, F.; Lee, J.; Wesselborg, S.; Feldbrüugge, M.; Müller, W.E.G.; Kalscheuer, R.; Ancheeva, E.; Proksch, P. Dithiodiketopiperazine derivatives from endophytic fungi Trichoderma harzianum and Epicoccum nigrum. Nat. Prod. Res. 2019, 35, 257–265. [Google Scholar] [CrossRef]
- Kock, I.; Draeger, S.; Schulz, B.; Elsässer, B.; Kurtán, T.; Kenéz, Á.; Antus, S.; Pescitelli, G.; Salvadori, P.; Speakman, J.B.; et al. Pseudoanguillosporin A and B: Two new isochromans isolated from the endophytic fungus Pseudoanguillospora sp. Eur. J. Org. Chem. 2009, 9, 1427–1434. [Google Scholar] [CrossRef]
- McMullin, D.R.; Nguyen, H.D.T.; Daly, G.J.; Menard, B.S.; Miller, J.D. Detection of foliar endophytes and their metabolites in Picea and Pinus seedling needles. Fungal Ecol. 2018, 31, 1–8. [Google Scholar] [CrossRef]
- Chapla, V.M.; Honório, A.E.; Gubiani, J.R.; Vilela, A.F.L.; Young, M.C.M.; Cardoso, C.L.; Pavan, F.R.; Cicarelli, R.M.; Michel Pinheiro Ferreira, P.; Bolzani, V.; et al. Acetylcholinesterase inhibition and antifungal activity of cyclohexanoids from the endophytic fungus Saccharicola sp. Phytochem. Lett. 2020, 39, 116–123. [Google Scholar] [CrossRef]
- Cao, L.L.; Zhang, Y.Y.; Liu, Y.J.; Yang, T.T.; Zhang, J.L.; Zhang, Z.G.; Shen, L.; Liu, J.Y.; Ye, Y.H. Anti-phytopathogenic activity of sporothriolide, a metabolite from endophyte Nodulisporium sp. A21 in Ginkgo biloba. Pestic. Biochem. Physiol. 2016, 129, 7–13. [Google Scholar] [CrossRef]
- Shentu, X.; Zhan, X.; Ma, Z.; Yu, X.; Zhang, C. Antifungal activity of metabolites of the endophytic fungus Trichoderma brevicompactum from garlic. Braz. J. Microbiol. 2014, 45, 248–254. [Google Scholar] [CrossRef]
- Li, X.; Wu, Y.; Liu, Z. Antifungal activity of an endophytic fungus Aspergillus versicolor DYSJ3 from Aphanamixis grandifolia Blume against Colletotrichum musae. Mycobiology 2021, 49, 498–506. [Google Scholar] [CrossRef]
- Wang, J.; Galgoci, A.; Kodali, S.; Herath, K.B.; Jayasuriya, H.; Dorso, K.; Vicente, F.; González, A.; Cully, D.; Bramhill, D.; et al. Discovery of a small molecule that inhibits cell division by blocking FtsZ, a novel therapeutic target of antibiotics. J. Biol. Chem. 2003, 278, 44424–44428. [Google Scholar] [CrossRef]
- Walters, D.; Raynor, L.; Mitchell, A.; Walker, R.; Walker, K. Antifungal activities of four fatty acids against plant pathogenic fungi. Mycopathologia 2004, 157, 87–90. [Google Scholar] [CrossRef]
- Liu, S.; Ruan, W.; Li, J.; Xu, H.; Wang, J.; Gao, Y.; Wang, J. Biological control of phytopathogenic fungi by fatty acids. Mycopathologia 2008, 166, 93–102. [Google Scholar] [CrossRef]
- Rojas Torres, M. Biocontrol de moniliophthora roreri y phytophthora palmivora Patógeno de Cacao (theobroma cacao l.) Con Hongos Endífitos Procedentes del Alto Amazonas. Master’s Thesis, Universidad Nacional de San Martin, Tarapoto, Perú, 2013. [Google Scholar]
- Myrchiang, P.; Dkhar, M.S.; Devi, H.R. Studies on endophytic fungi associated with medicinally important aromatic plant Artemisia nilagirica (C.B. Clarke) Pamp. and their antagonistic activity against Phytophthora infestans. J. Adv. Lab. Res. Biol. 2014, 5, 112–119. Available online: https://e-journal.sospublication.co.in/index.php/jalrb/article/view/202 (accessed on 1 December 2021).
- Simamora, A.V.; Hahuly, M.V.; Henuk, J.B. Endophytic fungi as potential biocontrol agents of Phytophthora palmivora in the cocoa plant. Biodiversitas 2021, 22, 2601–2609. [Google Scholar] [CrossRef]
- Soković, M.D.; Glamoclija, J.M.; Ciric, A.D. Natural products from plants and fungi as fungicides. In Fungicides-Showcases of Integrated Plant Disease Management from Around the World; Nita, M., Ed.; IntechOpen: London, UK, 2013. [Google Scholar] [CrossRef]
- Card, S.D.; Bastías, D.A.; Caradus, J.R. Antagonism to Plant Pathogens by Epichloë Fungal Endophytes—A Review. Plants 2021, 10, 1997. [Google Scholar] [CrossRef] [PubMed]
- Wäli, P.R.; Helander, M.; Nissinen, O.; Saikkonen, K. Susceptibility of endophyte-infected grasses to winter pathogens (snow molds). Botany 2006, 84, 1043–1051. [Google Scholar] [CrossRef]
- Xu, X.; Jeger, M. More ecological research needed for effective biocontrol of plant pathogens. In How Research Can Stimulate the Development of Commercial Biological Control Against Plant Diseases; de Cal, A., Melgarejo, P., Magan, N., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 15–30. [Google Scholar] [CrossRef]
- Ahlholm, J.U.; Helander, M.; Lehtimäki, S.; Wäli, P.; Saikkonen, K. Vertically transmitted fungal endophytes: Different responses of host-parasite systems to environmental conditions. Oikos 2002, 99, 173–183. [Google Scholar] [CrossRef]
- Moricca, S.; Ragazzi, A. Fungal endophytes in Mediterranean oak forests: A lesson from Discula quercina. Phytopathology 2008, 98, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Schulz, B.; Boyle, C. The endophytic continuum. Mycol. Res. 2005, 109, 661–686. [Google Scholar] [CrossRef]
- Delaye, L.; García-Guzmán, G.; Heil, M. Endophytes versus biotrophic and necrotrophic pathogens—are fungal lifestyles evolutionarily stable traits? Fungal Divers. 2013, 60, 125–135. [Google Scholar] [CrossRef]
- Guo, Q.-Y.; Cheng, L.; Zhu, H.-X.; Li, W.; Wei, Y.-H.; Chen, H.-Y.; Guo, L.-Z.; Weng, H.; Wang, J. Herbicidal activity of Aureobasidium pullulans PA-2 on weeds and optimization of its solid-state fermentation conditions. J. Integr. Agric. 2020, 19, 173–182. [Google Scholar] [CrossRef]
- Caradus, J.R.; Card, S.D.; Finch, S.C.; Hume, D.E.; Johnson, L.J.; Mace, W.J.; Popay, A.J. Ergot alkaloids in New Zealand pastures and their impact. N. Z. J. Agric. Res. 2020, 63, 1–41. [Google Scholar] [CrossRef]
- González-Menéndez, V.; Crespo, G.; Toro, C.; Martín, J.; de Pedro, N.; Tormo, J.R.; Genilloud, O. Extending the metabolite diversity of the endophyte Dimorphosporicola tragani. Metabolites 2019, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.B.; Gao, Y.Q.; Nie, X.D.; Tuong, T.M.; Li, D.; Gao, J.M. Antifungal activity of griseofulvin derivatives against phytopathogenic fungi in vitro and in vivo and three-dimensional quantitative structure-activity relationship analysis. J. Agric. Food Chem. 2019, 67, 6125–6132. [Google Scholar] [CrossRef] [PubMed]
- Kartsev, V.; Geronikaki, A.; Petrou, A.; Lichitsky, B.; Kostic, M.; Smiljkovic, M.; Soković, M.; Sirakanyanet, S. Griseofulvin derivatives: Synthesis, molecular docking and biological evaluation. Curr. Top. Med. Chem. 2019, 19, 1145–1161. [Google Scholar] [CrossRef]
- Ganeshan, S.; Kim, S.H.; Vujanovic, V. Scaling-up production of plant endophytes in bioreactors: Concepts, challenges and perspectives. Bioresour. Bioprocess. 2021, 8, 63. [Google Scholar] [CrossRef]
- Ministry of Science and Innovation of Spain. Gubernamental Strategy for Research, Technology and Innovation 2021–2023. Secretaría General Técnica del Ministerio de Ciencia e Innovación, Gobierno de España. 2021. Available online: https://www.ciencia.gob.es/gesdamdoc-servlet/?uuid=e1f1deb1-7321-4dd9-b8ca-f97ece358d1c&workspace=dam&formato=pdf (accessed on 1 December 2021).
- Wang, Q.; Peng, C.; Shi, L.; Liu, Z.; Zhou, D.; Meng, H.; Zhao, H.; Li, F.; Zhang, M. A technical system for the large-scale application of metabolites from Paecilomyces variotii SJ1 in agriculture. Front. Bioeng. Biotechnol. 2021, 9, 671879. [Google Scholar] [CrossRef]

| Fungal Isolate | Applied Crop | Target Fungal Pathogen | Application Form |
|---|---|---|---|
| Ampelomyces quisqualis | Grapes, tomato, pepper, aubergine | Powdery mildew | Spores |
| Aureobasidium pullulans | Pome fruit | Erwinia amylovora | CFU |
| Candida oleophila | Apples/pears | Postharvest diseases | CFU |
| Clonostachys rosea | Fruiting and leaf vegetables, seedlings, ornamentals, pot plants, cut flowers, wheat, corn, onion, potato, leek, berries | Seed borne and soil borne fungi, such as Fusarium, Pythium, Rhizoctonia and Phytophtora. Foliar pathogens, e.g., Botrytis and Didymella | CFU |
| Coniothyrium minitans | Winter rape, lettuce, cucumber, beans, sunflower | Sclerotinia sclerotiorum, S. minor | Spores |
| Pythium oligandrum | Oilseed rape | Sclerotinia sclerotiorum, Leptoshaeria maculans | Oospores |
| Saccharomyces cerevisiae | Pome fruits (apple, pear, quince, medlar, nashi) | Monilinia, Botrytis, Alternaria | CFU |
| Trichoderma asperellum | Tomato, pepper, cucumber, courgette, carnation plants growing in the greenhouse | Soil pathogens: Pythium spp., Rhizoctonia spp., Phytophthora spp., Phoma spp., Verticillium spp. and Fusarium spp. Fusarium oxysporum | CFU |
| Trichoderma atroviride | Grapevine, tomato | Wood decay diseases. Pythium spp. Rhizoctonia spp. Fusarium spp. | CFU |
| Trichoderma gamsii | Tomato, pepper, cucumber, courgette | Phythophthora sp. Fusarium sp. Rhizoctonia solani Pythium sp. Sclerotinia sclerotiorum | CFU |
| Verticillium albo-atrum | Elm trees | Vascular fungus: Ophiostoma novo-ulmi | Spores |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigo, S.; García-Latorre, C.; Santamaria, O. Metabolites Produced by Fungi against Fungal Phytopathogens: Review, Implementation and Perspectives. Plants 2022, 11, 81. https://doi.org/10.3390/plants11010081
Rodrigo S, García-Latorre C, Santamaria O. Metabolites Produced by Fungi against Fungal Phytopathogens: Review, Implementation and Perspectives. Plants. 2022; 11(1):81. https://doi.org/10.3390/plants11010081
Chicago/Turabian StyleRodrigo, Sara, Carlos García-Latorre, and Oscar Santamaria. 2022. "Metabolites Produced by Fungi against Fungal Phytopathogens: Review, Implementation and Perspectives" Plants 11, no. 1: 81. https://doi.org/10.3390/plants11010081
APA StyleRodrigo, S., García-Latorre, C., & Santamaria, O. (2022). Metabolites Produced by Fungi against Fungal Phytopathogens: Review, Implementation and Perspectives. Plants, 11(1), 81. https://doi.org/10.3390/plants11010081