

Optimizing Safe Approaches to Manage Plant-Parasitic Nematodes


Abstract
:1. Introduction
2. Biological Control Agents
2.1. Their General Categorization and Effects
2.2. Fungal and Bacterial Biocontrol
2.3. Nematode-Suppressive Soils
2.4. Evaluating Factors Affecting Their Success
2.4.1. Biological and Ecological Factors
2.4.2. Agricultural Practices
3. Botanicals as Bionematicides
3.1. Antagonistic Cultivated Plants
3.2. Plant-Related Materials and Compounds
3.3. Safety, Reliability, and Economics of the Related Nematicidal Products
4. Exploiting Poor- or Non-Host Crops
5. Other Methods of PPN Management
5.1. Additional Soil Amendments and Treatments
5.2. Advanced Methods
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Ibrahim, H.M.M.; Ahmad, E.M.; Martínez-Medina, A.; Aly, M.A.M. Effective approaches to study the plant-root knot nematode interaction. Plant Physiol. Biochem. 2019, 141, 332–342. [Google Scholar] [CrossRef] [PubMed]
- Molinari, S.; Leonetti, P. Bio-control agents activate plant immune response and prime susceptible tomato against root-knot nematodes. PLoS ONE 2019, 14, e0213230. [Google Scholar] [CrossRef]
- Hallmann, J.; Subbotin, S.A. Methods for extraction, processing and detection of plant and soil nematodes. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; Sikora, R.A., Coyne, D., Hallmann, J., Timper, P., Eds.; CABI: Wallingford, UK, 2018; pp. 87–119. [Google Scholar]
- Abdul Malik, N.A.; Kumar, I.S.; Nadarajah, K. Elicitor and receptor molecules: Orchestrators of plant defense and immunity. Int. J. Mol. Sci. 2020, 21, 963. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elgawad, M.M.M. Plant-parasitic nematodes and their biocontrol agents: Current status and future vistas. In Management of Phytonematodes: Recent Advances and Future Challenges; Ansari, R.A., Rizvi, R., Mahmood, I., Eds.; Springer Nature: Singapore, 2020; pp. 171–204. [Google Scholar] [CrossRef]
- Stirling, G.R. Biological control of plant-parasitic nematodes. In Diseases of Nematodes; Poinar, G.O., Jansson, H.-B., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 103–150. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M.; Askary, T.H. Fungal and bacterial nematicides in integrated nematode management strategies. Egypt. J. Biol. Pest Cont. 2018, 28, 74. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: Mechanisms and applications. Scientifica 2012, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Topalović, O.; Heuer, H.; Reineke, A.; Zinkernagel, J.; Hallmann, J. Antagonistic role of the microbiome from a Meloidogyne hapla-suppressive soil against species of plant-parasitic nematodes with different life strategies. Nematology 2019, 22, 75–86. [Google Scholar] [CrossRef]
- Topalović, O.; Hussain, M.; Heuer, H. Plants and associated soil microbiota cooperatively suppress plant-parasitic nematodes. Front. Microbiol. 2020, 11, 313. [Google Scholar] [CrossRef]
- Schouten, A. Mechanisms involved in nematode control by endophytic fungi. Annu. Rev. Phytopathol. 2016, 54, 121–142. [Google Scholar] [CrossRef]
- Ghahremani, Z.; Escudero, N.; Saus, E.; Gabaldón, T.; Sorribas, F.J. Pochonia chlamydosporia induces plant-dependent systemic resistance to Meloidogyne incognita. Front. Plant Sci. 2019, 10, 945. [Google Scholar] [CrossRef]
- Silva, S.D.; Carneiro, R.M.D.G.; Faria, M.; Souza, D.A.; Monnerat, R.G.; Lopes, R.B. Evaluation of Pochonia chlamydosporia and Purpureocillium lilacinum for suppression of Meloidogyne enterolobii on tomato and banana. J. Nematol. 2017, 49, 77–85. [Google Scholar] [CrossRef]
- Schouteden, N.; Waele, D.D.; Panis, B.; Vos, C.M. Arbuscular mycorrhizal fungi for the biocontrol of plant-parasitic nematodes: A review of the mechanisms involved. Front. Microbiol. 2015, 6, 1280. [Google Scholar] [CrossRef] [PubMed]
- Wolfgang, A.; Taffner, J.; Guimarães, R.A.; Coyne, D.; Berg, G. Novel strategies for soil-borne diseases: Exploiting the microbiome and volatile-based mechanisms toward controlling Meloidogyne-based disease complexes. Front. Microbiol. 2019, 10, 1296. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.L.; Shao, Z.Z.; Cai, M.M.; Zheng, L.Y.; Li, G.Y.; Yu, Z.N.; Zhang, J.B. Cyclo (l-Pro-l-Leu) of Pseudomonas putida MCCC 1A00316 isolated from Antarctic soil: Identification and characterization of activity against Meloidogyne incognita. Molecules 2019, 24, 768. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Yu, C.; Shao, Z.; Cai, M.; Li, G.; Zheng, L.; Yu, Z.; Zhang, J. Identification and characterization of nematicidal volatile organic compounds from deep-sea Virgibacillus dokdonensis MCCC 1A00493. Molecules 2020, 25, 744. [Google Scholar] [CrossRef]
- Forghani, F.; Hajihassani, A. Recent advances in the development of environmentally benign treatments to control root-knot nematodes. Front. Plant Sci. 2020, 11, 1125. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, H.J.; Li, J.Y.; Wang, C.; Chen, S.L.; Yan, S.Z. Effects of the endophytic bacteria Bacillus cereus BCM2 on tomato root exudates and Meloidogyne incognita infection. Pl. Dis. 2019, 103, 1551–1558. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, K.Y. Antagonistic potential of Bacillus pumilus L1 against root-knot nematode, Meloidogyne arenaria. J. Phytopathol. 2016, 164, 29–39. [Google Scholar] [CrossRef]
- Westphal, A. Detection and description of soils with specific nematode suppressiveness. J. Nematol. 2005, 37, 121–130. [Google Scholar] [PubMed]
- Eberlein, C.; Heuer, H.; Westphal, A. Biological suppression of populations of Heterodera schachtii adapted to different host genotypes of sugar beet. Front. Plant Sci. 2020, 11, 812. [Google Scholar] [CrossRef]
- Botelho, A.O.; Campos, V.P.; da Silva, J.C.P.; Freire, E.S.; de Pinho, R.S.C.; Barros, A.F.; Oliveira, D.F. Physicochemical and biological properties of the coffee (Coffea arabica) rhizosphere suppress the root-knot nematode Meloidogyne exigua. Biocontrol Sci. Technol. 2019, 29, 1181–1196. [Google Scholar] [CrossRef]
- Stirling, G.R.; Rames, E.; Stirling, A.M.; Hamill, S. Factors associated with the suppressiveness of sugarcane soils to plant-parasitic nematodes. J. Nematol. 2011, 43, 135–148. [Google Scholar]
- Cámara-Almirón, J.; Molina-Santiago, C.; Pérez-García, A.; de Vicente, A.; Cazorla, F.M.; Romero, D. Understanding bacterial physiology for improving full fitness. In How Research Can Stimulate the Development of Commercial Biological Control Against Plant Diseases, Progress in Biological Control; De Cal, A., Magan, N., Melgarejo, P., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 47–60. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Comments on the use of biocontrol agents against plant-parasitic nematodes. Int. J. PharmTech. Res. 2016, 9, 352–359. [Google Scholar]
- Dutta, T.K.; Khan, M.R.; Phani, V. Plant-parasitic nematode management via biofumigation using brassica and non-brassica plants: Current status and future prospects. Curr. Plant Biol. 2019, 17, 17–32. [Google Scholar] [CrossRef]
- Poveda, J.; Abril-Urias, P.; Escobar, C. Biological control of plant-parasitic nematodes by filamentous fungi inducers of resistance: Trichoderma, mycorrhizal and endophytic fungi. Front. Microbiol. 2020, 11, 992. [Google Scholar] [CrossRef]
- Duncan, L.W.; Stuart, R.J.; El-Borai, F.E.; Campos-Herrera, R.; Pathak, E.; Giurcanu, M.; Graham, J.H. Modifying orchard planting sites conserves entomopathogenic nematodes, reduces weevil herbivory and increases citrus tree growth, survival and fruit yield. Biol. Cont. 2013, 64, 26–36. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Spence, K.O.; Nakatani, J.; Lewis, E.E. Effect of soil salinity on entomopathogenic nematode survival and behaviour. Nematology 2011, 3, 859–867. [Google Scholar] [CrossRef]
- Hussaini, S.S. Entomopathogenic nematodes: Ecology, diversity and geographical distribution. In Biocontrol Agents: Entomopathogenic and Slug Parasitic Nematodes; Abd-Elgawad, M.M.M., Askary, T.H., Coupland, J., Eds.; CAB International: Wallingford, UK, 2017; pp. 88–142. [Google Scholar]
- Campos-Herrera, R.; Pathak, E.; El-Borai, F.E.; Schumann, A.; Abd-Elgawad, M.M.M.; Duncan, L.W. New citriculture system suppresses native and augmented entomopathogenic nematodes. Biol. Cont. 2013, 66, 183–194. [Google Scholar] [CrossRef]
- Campos-Herrera, R.; Stuart, R.J.; Pathak, E.; EL-Borai, F.E.; Duncan, L.W. Temporal patterns of entomopathogenic nematodes in Florida citrus orchards: Evidence of natural regulation by microorganisms and nematode competitors. Soil Biol. Biochem. 2019, 128, 193–204. [Google Scholar] [CrossRef]
- Giblin-Davis, R.M.; Williams, D.S.; Bekal, S.; Dickson, D.W.; Becker, J.O.; Preston, J.F. ‘Candidatus Pasteuria usgae’ sp. nov., an obligate endoparasite of the phytoparasitic nematode, Belonolaimus longicaudatus. Int. J. Syst. Evol. Microbiol. 2003, 53, 197–200. [Google Scholar] [CrossRef]
- Timper, P. Conserving and enhancing biological control of nematodes. J. Nematol. 2014, 46, 75–89. [Google Scholar] [PubMed]
- Sikora, R.A.; Roberts, P.A. Management Practices: An overview of integrated nematode management technologies. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; Sikora, R.A., Coyne, D., Hallmann, J., Timper, P., Eds.; CABI: Wallingford, UK, 2018; pp. 795–838. [Google Scholar]
- Abd-Elgawad, M.M.M. Managing nematodes in Egyptian citrus orchards. Bull. NRC 2020, 44, 41. [Google Scholar] [CrossRef]
- Ma, Y.; Li, Y.; Lai, H.; Guo, Q.; Xue, Q. Effects of two strains of Streptomyces on root-zone microbes and nematodes for biocontrol of root-knot nematode disease in tomato. Appl. Soil. Ecol. 2017, 112, 34–41. [Google Scholar] [CrossRef]
- Gorny, A.M.; Hay, F.S.; Esker, P.; Pethybridge, S.J. Spatial and spatiotemporal analysis of Meloidogyne hapla and Pratylenchus penetrans populations in commercial potato fields in New York, USA. Nematology 2020, 22, 1–13. [Google Scholar] [CrossRef]
- Escudero, N.; Lopez-Moya, F.; Ghahremani, Z.; Zavala-Gonzalez, E.A.; Alaguero-Cordovilla, A.; Ros-Ibañez, C.; Lacasa, A.; Sorribas, F.J.; Lopez-Llorca, L.V. Chitosan increases tomato root colonization by Pochonia chlamydosporia and their combination reduces root-knot nematode damage. Front. Plant Sci. 2017, 8, 1415. [Google Scholar] [CrossRef] [PubMed]
- El-Nagdi, W.M.A.; Youssefi, M.M.A.; Abd-El-Khair, H.; Abd-Elgawad, M.M.M.; Dawood, M.G. Effectiveness of Bacillus subtilis, B. pumilus, Pseudomonas fluorescens on Meloidogyne incognita infecting cowpea. Pak. J. Nematol. 2019, 37, 35–43. [Google Scholar] [CrossRef]
- Dahlin, P.; Eder, R.; Consoli, E.; Krauss, J.; Kiewnick, S. Integrated control of Meloidogyne incognita in tomatoes using fluopyram and Purpureocillium lilacinum strain 251. Crop Prot. 2019, 124, 104874. [Google Scholar] [CrossRef]
- d’Errico, G.; Marra, R.; Crescenzi, A.; Davino, S.W.; Fanigliulo, A.; Woo, S.L.; Lorito, M. Integrated management strategies of Meloidogyne incognita and Pseudopyrenochaeta lycopersici on tomato using a Bacillus firmus-based product and two synthetic nematicides in two consecutive crop cycles in greenhouse. Crop Prot. 2019, 122, 159–164. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Status of entomopathogenic nematodes in integrated pest management strategies in Egypt. In Biocontrol Agents: Entomopathogenic and Slug Parasitic Nematodes; Abd-Elgawad, M.M.M., Askary, T.H., Coupland, J., Eds.; CABI: Wallingford, UK, 2017; pp. 473–501. [Google Scholar]
- Hussain, M.; Zouhar, M.; Rysanek, P. Suppression of Meloidogyne incognita by the entomopathogenic fungus Lecanicillium muscarium. Plant Dis. 2018, 102, 977–982. [Google Scholar] [CrossRef]
- Sharma, I.P.; Sharma, A.K. Physiological and biochemical changes in tomato cultivar PT-3 with dual inoculation of mycorrhiza and PGPR against root-knot nematode. Symbiosis 2017, 71, 175–183. [Google Scholar] [CrossRef]
- Anonymous. Adopted recommendations to combat agricultural pests. In APC BooK 2018; Commercial Al-Ahram Press: Qalioub, Egypt, 2018; (In Arabic) Agricultural Pesticide Committee, Ministry of Agriculture, Media Support Center Press. Available online: http://www.apc.gov.eg/Files/Releases/Recomm18/mobile/index.html#p=16 (accessed on 17 August 2021).
- Grubišić, D.; Uroić, G.; Ivošević, A.; Grdiša, M. Nematode control by the use of antagonistic plants. Agric. Consp. Sci. 2018, 83, 269–275. [Google Scholar]
- Hamaguchi, T.; Sato, K.; Vicente, C.S.L.; Hasegawa, K. Nematicidal actions of the marigold exudate α-terthienyl: Oxidative stress-inducing compound penetrates nematode hypodermis. Biol. Open. 2019, 8, bio038646. [Google Scholar] [CrossRef]
- Ntalli, N.; Caboni, P. A review of isothiocyanates biofumigation activity on plant parasitic nematodes. Phytochem. Rev. 2017, 16, 827–834. [Google Scholar] [CrossRef]
- Sikora, R.A.; Coyne, D.; Hallmann, J.; Timper, P. Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, 3rd ed.; CABI: Cambridge, UK, 2018. [Google Scholar]
- Chiamolera, F.M.; Dias-Arieira, C.R.; de Souto, E.R.; Biela, F.; da Cunha, T.P.L.; Santana, S.D.M.; Puerari, H.H. Susceptibility of winter crops to Pratylenchus brachyurus and effect on the nematode population in the maize crop. Nematropica 2012, 42, 267–275. [Google Scholar]
- Ntalli, N.; Ratajczak, M.; Oplos, C.; Menkissoglu-Spiroudi, U.; Adamski, Z. Acetic acid, 2-undecanone, and (E)-2-decenal ultrastructural malformations on Meloidogyne incognita. J. Nematol. 2016, 48, 248–260. [Google Scholar] [CrossRef]
- Seo, H.J.; Park, A.R.; Kim, S.; Yeon, J.; Yu, N.H.; Ha, S.; Chang, J.Y.; Palrk, H.W.; Kim, J.-C. Biological control of root-knot nematodes by organic acid-producing Lactobacillus brevis wikim0069 isolated from kimchi. Plant Pathol. J. 2019, 35, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Zasada, I.A.; Meyer, S.L.F.; Halbrendt, J.M.; Rice, C. Activity of hydroxamic acids from Secale cereale against the plant-parasitic nematodes Meloidogyne incognita and Xiphinema americanum. Phytopathology 2005, 95, 1116–1121. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, S.; Grassi de Freitas, L. Use of antagonistic plants and natural products. In Nematology-Advances and Perspectives; Nematode Management and Utilisation; Chen, Z.X., Chen, S.Y., Dickson, D.W., Eds.; CABI Publishing: Wallingford, UK, 2004; pp. 931–977. Volume II. [Google Scholar]
- Abd-Elgawad, M.M.M.; Omer, E.A. Effect of essential oils of some medicinal plants on phytonematodes. Anz. Schädlingskunde Pflanzenschutz. Umweltschutz/J. Pest Sci. 1995, 68, 82–84. [Google Scholar] [CrossRef]
- Barros, A.F.; Campos, V.P.; De Oliveira, D.F.; De Jesus Silva, F.; Jardim, I.N.; Costa, V.A.; Matrangolo, C.A.R.; Ribeiro, R.C.F.; Silval, G.H. Activities of essential oils from three Brazilian plants and benzaldehyde analogues against Meloidogyne incognita. Nematology 2019, 21, 1081–1089. [Google Scholar] [CrossRef]
- Kalaiselvi, D.; Mohankumar, A.; Shanmugam, G.; Thiruppathi, G.; Nivitha, S.; Sundararaj, P. Altitude-related changes in the phytochemical profile of essential oils extracted from Artemisia nilagirica and their nematicidal activity against Meloidogyne incognita. Ind. Crops Prod. 2019, 139, 111472. [Google Scholar] [CrossRef]
- Zaidat, S.A.E.; Mouhouche, F.; Babaali, D.; Abdessemed, N.; De Cara, M.; Hammache, M. Nematicidal activity of aqueous and organic extracts of local plants against Meloidogyne incognita (Kofoid and White) Chitwood in Algeria. Egypt. J. Biol. Pest Cont. 2020, 30, 46. [Google Scholar] [CrossRef]
- Montiel-Rozas, M.; Díez-Rojo, M.; Ros, M.; Pascual, J.A. effect of plant extracts and metam sodium on the soilborne fungal pathogens, Meloidogyne spp., and soil microbial community. Agronomy 2020, 10, 513. [Google Scholar] [CrossRef]
- Jindapunnapat, K.; Reetz, N.D.; MacDonald, M.H.; Bhagavathy, G.; Chinnasri, B.; Soonthornchareonnon, N.; Sasnarukkit, A.; Chauhan, K.R.; Chitwood, D.J.; Meyer, S.L. Activity of vetiver extracts and essential oil against Meloidogyne incognita. J. Nematol. 2018, 50, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Meyer, S.L.F.; MacDonald, M.H.; Zheng, L.; Jing, C.; Chitwood, D.J. Nematotoxicity of Paeonia spp. extracts and Camellia oleifera tea seed cake and extracts to Heterodera glycines and Meloidogyne incognita. Plant Dis. 2019, 103, 2191–2198. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elgawad, M.M.M.; Mohamed, M.M.M. Efficacy of selected biocontrol agents on Meloidogyne incognita on eggplant. Nematol. Medit. 2006, 34, 105–109. [Google Scholar]
- Ji, X.; Li, J.; Meng, Z.; Dong, S.; Zhang, S.; Qiao, K. Inhibitory effect of allicin against Meloidogyne incognita and Botrytis cinerea in tomato. Sci. Hortic. 2019, 253, 203–208. [Google Scholar] [CrossRef]
- Wilson, M.J.; Jackson, T.A. Progress in the commercialization of bionematicides. BioControl 2013, 58, 715–722. [Google Scholar] [CrossRef]
- Ramirez, R.A.; Henderson, D.R.; Riga, E.; Lacey, L.A.; Synder, W.E. Harmful effects of mustard green manure on entomopathogenic nematodes, Biol. Cont. 2009, 48, 147–154. [Google Scholar]
- Moreira, F.J.C.; Barbosa da Silva, M.C.; Araujo Rodrigues, A.; Neves Tavares, M.K. Alternative control of root-knot nematodes (Meloidogyne javanica and M. enterolobii) using antagonists. Int. J. Agron. Agric. Res. 2015, 7, 121–129. [Google Scholar]
- Ndeve, N.D.; Matthews, W.C.; Santos, J.R.P.; Huynh, B.L.; Roberts, P.A. Broad-based root-knot nematode resistance identified in cowpea gene-pool two. J. Nematol. 2018, 50. [Google Scholar] [CrossRef] [PubMed]
- Zwart, R.S.; Thudi, M.; Channale, S.; Manchikatla, P.K.; Varshney, R.K.; Thompson, J.P. Resistance to plant-parasitic nematodes in chickpea: Current status and future perspectives. Front. Plant Sci. 2019, 10, 966. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elgawad, M.M.M. A new rating scale for screening plant genotypes against root-knot and reniform nematodes. Anz. Schadling. Pflanzenschutz. Umweltschutz/J. Pest Sci. 1991, 64, 37–39. [Google Scholar] [CrossRef]
- Molinari, S. Resistance and virulence in plant-nematode interactions. In Nematodes; Boeri, F., Chung, J.A., Eds.; Nova Science Publisher Inc.: New York, NY, USA, 2012; pp. 59–82. [Google Scholar]
- Noling, J.W. Nematode Management in Tomatoes, Peppers, and Eggplant. 2019. Available online: https://edis.ifas.ufl.edu/publication/NG032 (accessed on 17 August 2021).
- Starr, J.L.; Cook, R.; Bridge, J. Plant Resistance to Parasitic Nematodes; CABI: Wallingford, UK, 2002. [Google Scholar]
- Starr, J.L.; Roberts, P.A. Resistance to plant-parasitic nematodes. In Nematology, Advances and Perspective; Chen, Z.X., Chen, S.Y., Dickson, D.W., Eds.; CABI: Wallingford, UK, 2004; Volume 2, pp. 879–907. [Google Scholar]
- Abd-Elgawad, M.M.M. Towards sound use of statistics in nematology. Bull. NRC. 2021, 45, 13. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M.; Molinari, S. Markers of plant resistance to nematodes: Classical and molecular strategies. Nematol. Medit. 2008, 36, 3–11. [Google Scholar]
- Lopez-Perez, J.A.; Roubtsova, T.; Ploeg, A. Effect of three plant residues and chicken manure used as biofumigants at three temperatures on Meloidogyne incognita infestation of tomato in greenhouse experiments. J. Nematol. 2005, 37, 489–494. [Google Scholar]
- Ntalli, N.; Adamski, Z.; Doula, M.; Monokrousos, N. Nematicidal amendments and soil remediation. Plants 2020, 9, 429. [Google Scholar] [CrossRef]
- Vu, T.; Hauschild, R.; Sikora, R.A. Fusarium oxysporum endophytes induced systemic resistance against Radopholus similis on banana. Nematology 2006, 8, 847–852. [Google Scholar] [CrossRef]
- Hahn, M.H.; May De Mio, L.L.; Kuhn, O.J.; Da Silva Silveira Duarte, H. Nematophagous mushrooms can be an alternative to control Meloidogyne javanica. Biol. Cont. 2019, 138, 104024. [Google Scholar] [CrossRef]
- Youssef, M.M.A.; El-Nagdi, W.M.A. Population density of Meloidogyne incognita and eggplant growth vigour affected by sucrose-activated bread yeast (Saccharomyces cerevisiae). Pak. J. Nematol. 2018, 36, 117–122. [Google Scholar]
- Ghareeb, R.Y.; Basyony, A.B.A.; Al-nazwani, M.S.; El-Saedy, M.A.M. Biocontrol of Meloidogyne incognita attacking potato plants using the extract of a new isolate of Chlorella vulgaris and molecular characterization. Pl. Cell Biotech. Molecul. Biol. 2021, 13–14, 47–62. [Google Scholar]
- Blyuss, K.B.; Fatehi, F.; Tsygankova, V.A.; Biliavska, L.O.; Iutynska, G.O.; Yemets, A.I.; Blume, Y.B. RNAi-based biocontrol of wheat nematodes using natural poly-component biostimulants. Front. Plant Sci. 2019, 10, 483. [Google Scholar] [CrossRef] [PubMed]
- Rouse-Miller, J.; Bartholomew, E.S.; St. Martin, C.C.G.; Vilpigue, P. Bioprospecting compost for long-term control of plant parasitic nematodes. In Management of Phytonematodes: Recent Advances and Future Challenges; Ansari, R., Rizvi, R., Mahmood, I., Eds.; Springer Nature: Singapore, 2020; pp. 35–50. [Google Scholar]
- Hallmann, J.; Meressa, B.H. Nematode parasites of vegetables. In Plant Parasitic Nematodes in Subtropical and Tropical Agriculture; Sikora, R.A., Coyne, D., Hallmann, J., Timper, P., Eds.; CABI: Wallingford, UK, 2018; pp. 346–410. [Google Scholar]
- Abd-Elgawad, M.M.M.; Elshahawy, I.E.; Abd-El-Kareem, F. Efficacy of soil solarization on black root rot disease and speculation on its leverage on nematodes and weeds of strawberry in Egypt. Bull. NRC. 2019, 43, 175. [Google Scholar] [CrossRef]
- Kokalis-Burelle, N.; Rosskopf, E.N.; Butler, D.M.; Fennimore, S.A.; Holzinger, J. Evaluation of steam and soil solarization for Meloidogyne arenaria control in Florida floriculture crops. J. Nematol. 2016, 48, 183–192. [Google Scholar] [CrossRef]
- Veronico, P.; Paciolla, C.; Sasanelli, N.; De Leonardis, S.; Melillo, M.T. Ozonated water reduces susceptibility in tomato plants to Meloidogyne incognita by the modulation of the antioxidant system. Mol. Plant Pathol. 2017, 18, 529–539. [Google Scholar] [CrossRef]
- Magan, N. Importance of ecological windows for efficacy of biocontrol agents. In How Research Can Stimulate the Development of Commercial Biological Control Against Plant Diseases, Progress in Biological Control; De Cal, A., Magan, N., Melgarejo, P., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 1–14. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M.; Askary, T.H. Factors affecting success of biological agents used in controlling plant-parasitic nematodes. Egypt. J. Biol. Pest Cont. 2020, 30, 17. [Google Scholar] [CrossRef]
- Barthès, B.G.; Brunet, D.; Rabary, B.; Ba, O.; Villenave, C. Near infrared reflectance spectroscopy (NIRS) could be used for characterization of soil nematode community. Soil Biol. Biochem. 2011, 43, 1649–1659. [Google Scholar] [CrossRef]
- Fan, Z.; Qin, Y.; Liu, S.; Xing, R.; Yu, H.; Chen, X.; Li, K.; Li, R.; Wang, X.; Li, P. The bioactivity of new chitin oligosaccharide dithiocarbamate derivatives evaluated against nematode disease (Meloidogyne incognita). Carbohydr. Polym. 2019, 224, 115155. [Google Scholar] [CrossRef]
- Abd-El-Kareem, F.; Elshahawy, I.E.; Abd-Elgawad, M.M.M. Management of strawberry leaf blight disease caused by Phomopsis obscurans using silicate salts under field conditions. Bull. NRC 2019, 43, 1. [Google Scholar] [CrossRef]
- Roldi, M.; Dias-Arieira, C.R.; Da Silva, S.A.; Dorigo, O.F.; Machado, A.C.Z. Control of Meloidogyne paranaensis in coffee plants mediated by silicon. Nematology 2017, 19, 245–250. [Google Scholar] [CrossRef]
- Brito, O.D.C.; Ferreira, J.C.A.; Hernandes, I.; Silva, E.J.; Dias-Arieira, C.R. Management of Meloidogyne javanica on tomato using agro-industrial wastes. Nematology 2020, 1–14. [Google Scholar] [CrossRef]
- Cheng, F.; Wang, J.; Song, Z.; Cheng, J.; Zhang, D.; Liu, Y. Nematicidal effects of 5-Aminolevulinic acid on plant-parasitic nematodes. J. Nematol. 2017, 49, 295–303. [Google Scholar] [CrossRef]
- DiLegge, M.J.; Manter, D.K.; Vivanco, J.M. A novel approach to determine generalist nematophagous microbes reveals Mortierella globalpina as a new biocontrol agent against Meloidogyne spp. nematodes. Sci. Rep. 2019, 9, 7521. [Google Scholar] [CrossRef]
- Safeena, M.I.S.; Zakeel, M.C.M. Nanobiotechnology-driven management of phytonematodes. In Management of phytonematodes: Recent Advances and Future Challenges; Ansari, R.A., Rizvi, R., Mahmood, I., Eds.; Springer Nature: Singapore, 2020; pp. 1–33. [Google Scholar]
- Abd-Elgawad, M.M.M. Optimizing sampling and extraction methods for plant-parasitic and entomopathogenic nematodes. Plants 2021, 10, 629. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elgawad, M.M.M. Optimizing biological control agents for controlling nematodes of tomato in Egypt. Egypt. J. Biol. Pest Cont. 2020, 58, 1–10. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Biological control agents in the integrated nematode management of pepper in Egypt. Egypt. J. Biol. Pest Cont. 2020, 30, 70. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Biological control agents in the integrated nematode management of potato in Egypt. J. Biol. Pest Cont. 2020, 30, 121. [Google Scholar] [CrossRef]
- Abd-Elgawad, M.M.M. Biological control of nematodes infecting eggplant in Egypt. Bull. NRC 2021, 45, 6. [Google Scholar] [CrossRef]
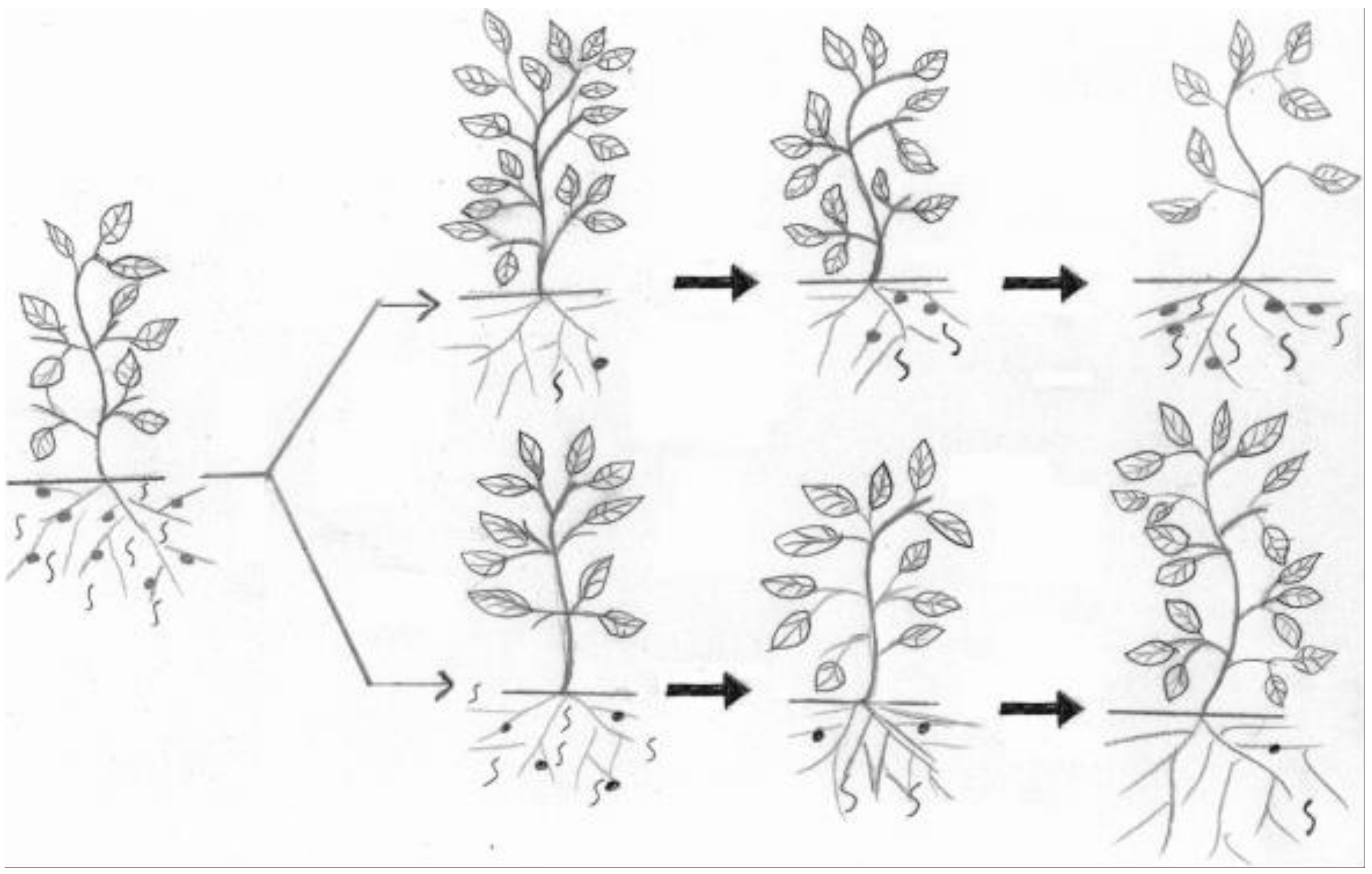

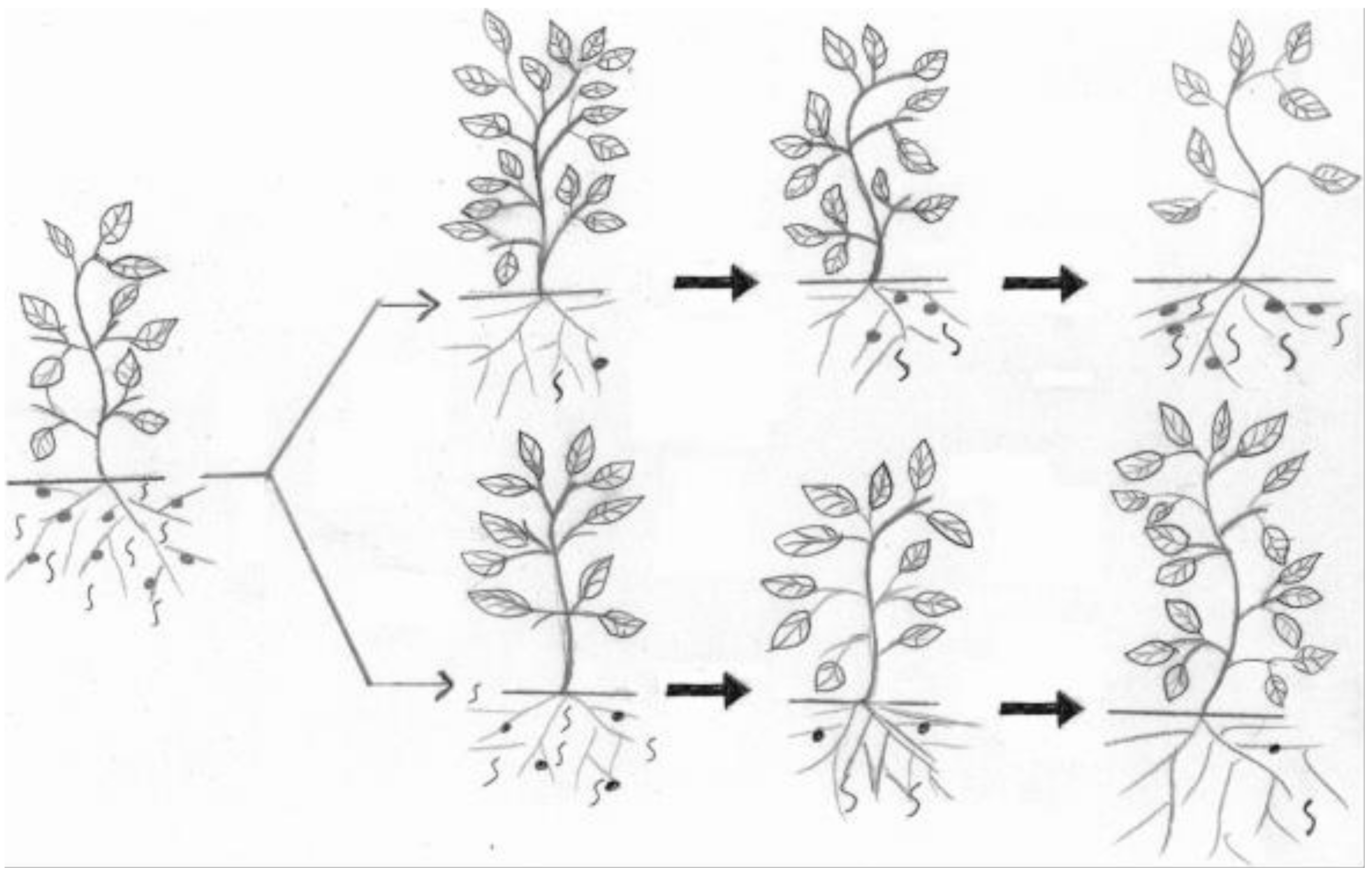{kind=link}
| Biological Control Agent | Nematode Species | Type of Study | Host Plant | Reference |
|---|---|---|---|---|
| Bacteria | ||||
| Bacillus firmus | Meloidogyne incognita | In vivo | tomato | [43] |
| Pasteuria penetranse | Meloidogyne exigua | In vivo | coffee | [23] |
| PGPR: Pseudomonas jessenii and P. synxantha | M. incognita | In vivo | tomato | [46] |
| Fungi | ||||
| (A) Filamentous: Trichoderma spp. | Rotylenchulus reniformis, M. javanica, M. incognita, Heterodera cajani | In vivo | tomato, brinjal, okra, soybean, sugarbeet, pigeonpea | [7] |
| AMF: Rhizophagus irregularis | M. incognita | In vivo | tomato | [46] |
| Endophyte: Fusarium oxysporum | Radopholus similis | In vivo | banana | [80] |
| (B) Mushrooms: Lentinula edodes, Macrocybe titans, Pleurotus eryngii | M. javanica | In vitro | tomato | [81] |
| (C) Yeasts: Saccharomyces cerevisiae | M. incognita | In vivo | eggplant | [82] |
| Co-application: Pochonia chlamydosporia and Chitosan | M. javanica | In vivo | tomato | [40] |
| Sequential application: Fluopyram and Purpureocillium lilacinum | M. incognita | In vivo | tomato | [42] |
| Dual-purpose: Heterorhabditis bacteriophora EGG | M. incognita | In vivo | watermelon | [44] |
| Algae: Chlorella vulgaris | M. incognita | In planta | potato | [83] |
| Nematode-suppressive soil | M. hapla, Pratylenchus neglectus | In vivo | tomato | [9] |
| Botanicals: Tagetes spp. | M. incognita, M. javanica, M. acrita | In vivo | tomato and eggplant | [48] |
| Soil amendments | M. incognita, Heterodera glycines | In vivo | tomato and soybean | [79] |
| RNA interference via stimulants of soil streptomycetes | Heterodera avenae | In planta | wheat | [84] |
| Active Ingredient | Product Name | Application Rate (Product Hectare −1) + | Price per Hectare |
|---|---|---|---|
| Abamectin (soluble concentrate at 20 g/L) generated from the fermentation process of Streptomyces avermitilis | Tervigo 2% SC | 5.95 L/Hectare | USD 319 |
| 109 CFU/mL of Serratia sp., Pseudomonas sp., Azotobacter sp., Bacillus circulans and B. thuringiensis | Micronema | 71.4 L/Hectare (thrice)/year | USD 40 |
| 108 units/mL Purpureocillium lilacinus | Bio-Nematon | 4.76 L/Hectare/year | USD 78 |
| 109 bacterium cells of Serratia marcescens/mL water | Nemaless | 23.8 L/Hectare (thrice)/year | USD 95 |
| Cadusafos (O-ethyl S,S-bis (1-methylpropyl) phosphorodithioate) | Rugby 10 G | 57.14 Kg/Hectare | USD 1028 |
| Oxamyl (methyl 2-(dimethylamino)-N-(methylcarbamoyloxy)-2 oxoethanimidothioate) | Vydate 24% SL | 9.52 L/Hectare (twice)/year | USD 445 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-Elgawad, M.M.M. Optimizing Safe Approaches to Manage Plant-Parasitic Nematodes. Plants 2021, 10, 1911. https://doi.org/10.3390/plants10091911
Abd-Elgawad MMM. Optimizing Safe Approaches to Manage Plant-Parasitic Nematodes. Plants. 2021; 10(9):1911. https://doi.org/10.3390/plants10091911
Chicago/Turabian StyleAbd-Elgawad, Mahfouz M. M. 2021. "Optimizing Safe Approaches to Manage Plant-Parasitic Nematodes" Plants 10, no. 9: 1911. https://doi.org/10.3390/plants10091911
APA StyleAbd-Elgawad, M. M. M. (2021). Optimizing Safe Approaches to Manage Plant-Parasitic Nematodes. Plants, 10(9), 1911. https://doi.org/10.3390/plants10091911