
Plant Growth Regulators Application Enhance Tolerance to Salinity and Benefit the Halophyte Plantago coronopus in Saline Agriculture



Abstract
1. Introduction
2. Results
2.1. Effect of Plant Growth Regulators (PGRs) Application on Growth of P. coronopus
2.2. Effect of PGRs Application on Sorbitol Content
2.3. Effect of PGRs Application on the Total Amount of Phenols and Flavonoids in Saline Conditions
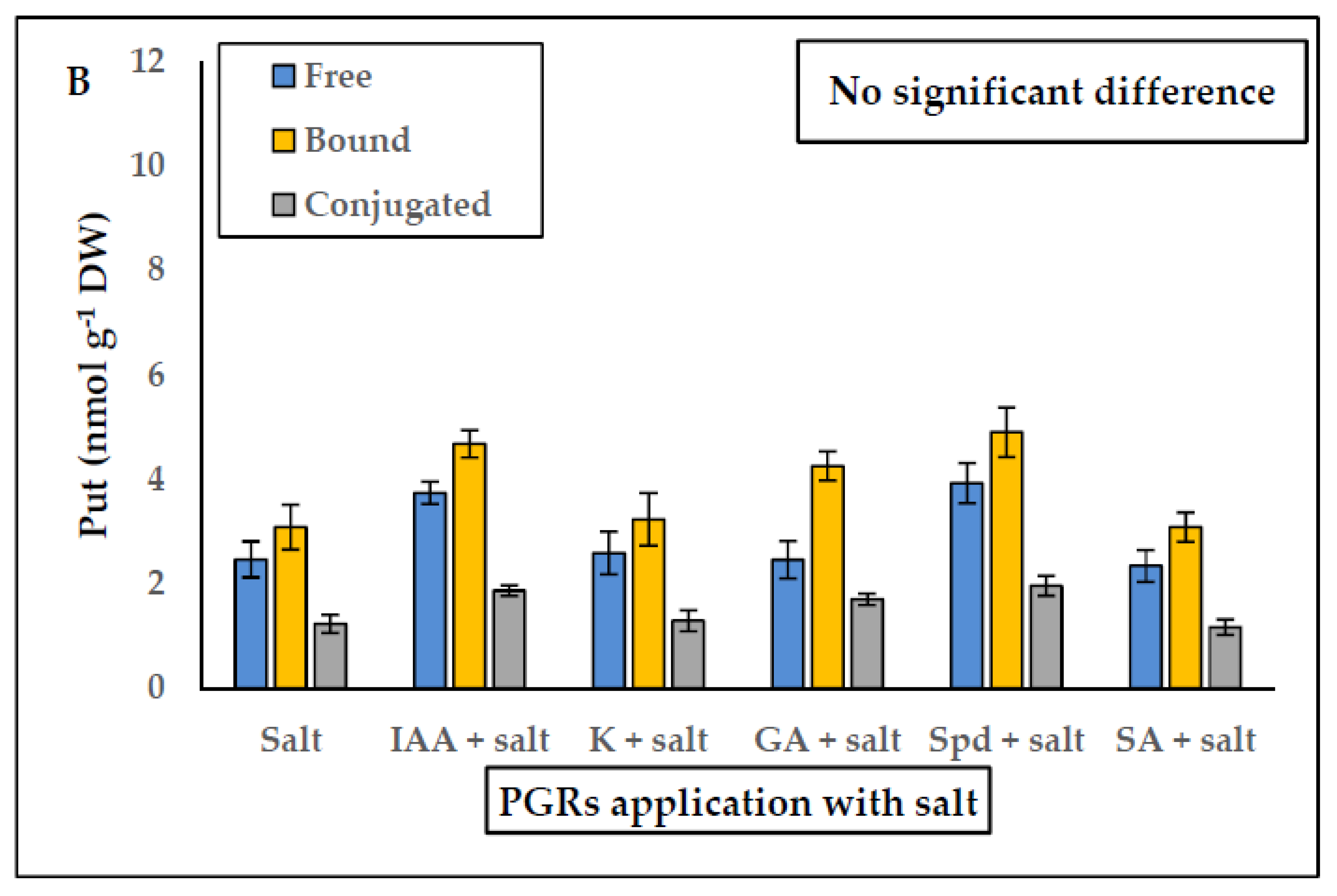
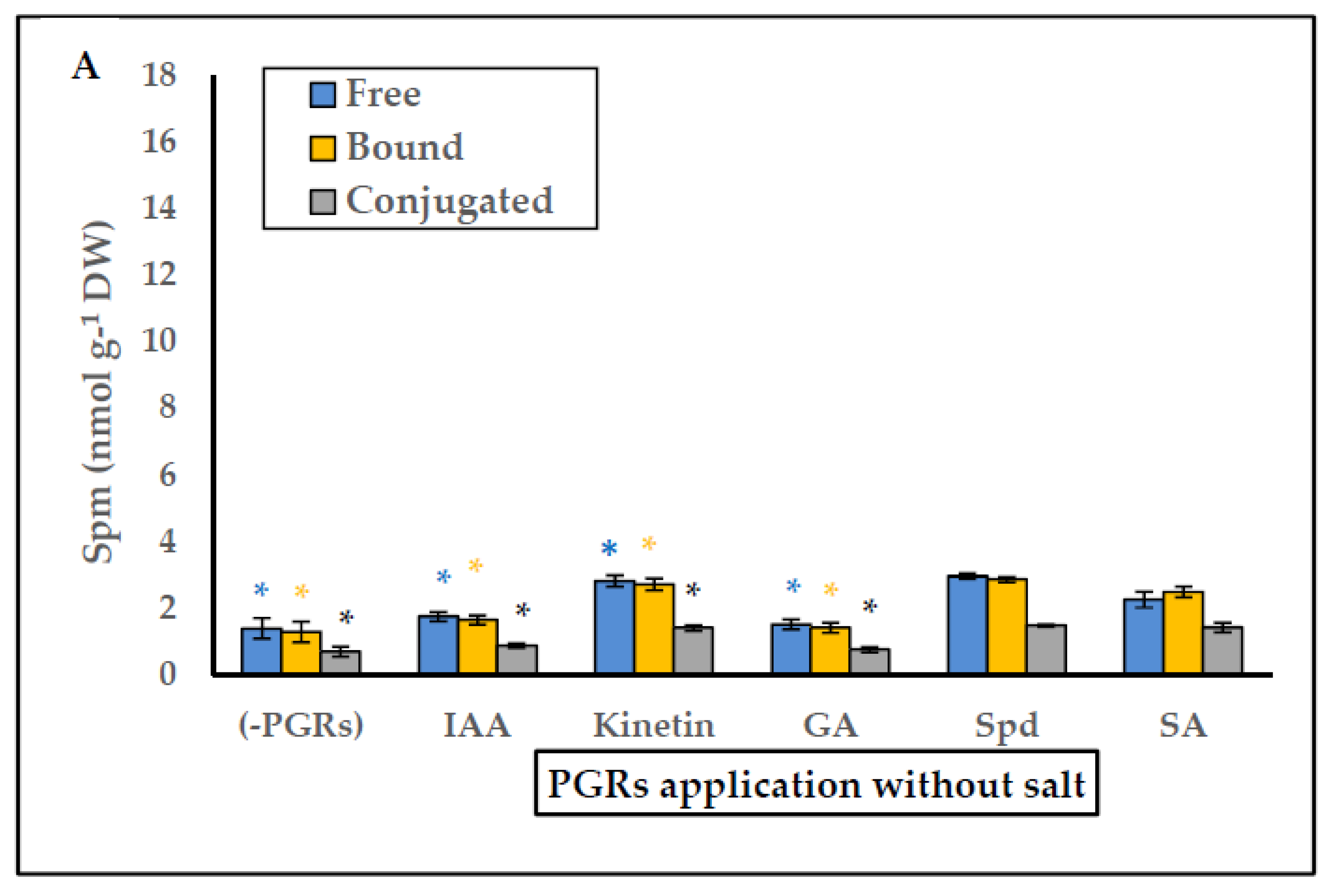
2.4. Effect of PGRs Application on Endogenous Free, Bound and Conjugated Polyamines and Ethylene
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Experimental Design and Treatments with PGRs and NaCl
- No PGRs
- IAA (1 µM)
- Kinetin (1 µM)
- GA3 (1 µM)
- Spd (0.5 mM)
- SA (0.5 mM)
- 7.
- NaCl (200 mM)
- 8.
- IAA (1 µM) + NaCl (200 mM)
- 9.
- Kinetin (1 µM) + NaCl (200 mM)
- 10.
- GA3 (1 µM) + NaCl (200 mM)
- 11.
- Spd (0.5 mM) + NaCl (200 mM)
- 12.
- SA (0.5 mM) + NaCl (200 mM)
4.3. Growth Parameters
4.4. Sorbitol Quantification
4.5. Determination of Total Phenols and Flavonoids
4.6. Analysis of Free, Bound and Conjugated Polyamines
4.7. Ethylene Production
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Thornton, P.K.; Ericksen, P.J.; Herrero, M.; Challinor, A.J. Climate variability and vulnerability to climate change: A review. Glob. Chang. Biol. 2014, 20, 3313–3328. [Google Scholar] [CrossRef]
- Leisner, C.P. Review: Climate change impacts on food security-focus on perennial cropping systems and nutritional value. Plant Sci. 2020, 293, 110412. [Google Scholar] [CrossRef]
- Shahzad, A.; Ullah, S.; Dar, A.A.; Sardar, M.F.; Mehmood, T.; Tufail, M.A.; Shakoor, A.; Haris, M. Nexus on climate change: Agriculture and possible solution to cope future climate change stress. Environ. Sci. Pollut. Res. Int. 2021, 28, 14211–14232. [Google Scholar] [CrossRef]
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. eHALOP a database of salt-tolerant plants: Helping put halophytes to work. Plant Cell Physiol. 2016, 57, e10. [Google Scholar] [CrossRef]
- Grigore, M.N. Definition and classification of halophytes as an ecological group of plants. In Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture, 1st ed.; Grigore, M.N., Ed.; Springer: Cham, Switzerland, 2021; pp. 1–48. [Google Scholar] [CrossRef]
- Gul, B.; Ansari, R.; Flowers, T.J.; Khan, M.A. Germination strategies of halophyte seeds under salinity. Environ. Exp. Bot. 2013, 92, 4–18. [Google Scholar] [CrossRef]
- Kumari, A.; Das, P.; Parida, A.K.; Agarwal, P.K. Proteomics, metabolomics, and ionomics perspectives of salinity tolerance in halophytes. Front. Plant Sci. 2015, 6, 537. [Google Scholar] [CrossRef]
- Bueno, M.; Cordovilla, M.P. Ecophysiology and uses of halophytes in diverse habitats. In Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture, 1st ed.; Grigore, M.N., Ed.; Springer: Cham, Switzerland, 2021; pp. 1–25. [Google Scholar] [CrossRef]
- Grigore, M.N.; Toma, C. Integrative anatomy of halophytes from Mediterranean climate. In Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture, 1st ed.; Grigore, M.N., Ed.; Springer: Cham, Switzerland, 2021; pp. 1–35. [Google Scholar] [CrossRef]
- Shaid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; García-Sánchez, F. Insights into physiological and biochemical impacts of salt stress on plant growth and development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Borsai, O.; Al Hassan, M.; Negrusier, C.; Raigón, M.D.; Boscaiu, M.; Sestraş, R.E.; Vicente, O. Responses to salt stress in Portulaca: Insight into its tolerance mechanism. Plants 2020, 9, 1660. [Google Scholar] [CrossRef]
- Panta, S.; Flowers, T.; Lane, P.; Doyle, R.; Haros, G.; Shabala, S. Halophyte agriculture: Success stories. Environ. Exp. Bot. 2014, 107, 71–83. [Google Scholar] [CrossRef]
- Atzori, G. The potential of edible halophytes as new crops in saline agriculture. The ice plant (Mesembryanthemum crystallinum L.) case study. In Future of Sustainable Agriculture in Saline Environments, 1st ed.; CRC Press: Boca Raton, NY, USA, 2021; pp. 443–460. [Google Scholar] [CrossRef]
- Nikalje, G.C.; Srivastava, A.K.; Pandey, G.K.; Suprasanna, P. Halophytes in biosaline agriculture: Mechanism, utilization, and value addition. Land Degrad. Dev. 2018, 29, 1081–1095. [Google Scholar] [CrossRef]
- Christiansen, A.H.C.; Lyra, D.A.; Jørgensen, H. Increasing the value of Salicornia bigelovii green biomass grown in a desert environment through biorefining. Ind. Crops Prod. 2021, 160, 113105. [Google Scholar] [CrossRef]
- Duarte, B.; Caҫador, I. Iberian halophytes as agroecological solutions for degraded lands and biosaline agriculture. Sustainability 2021, 13, 1005. [Google Scholar] [CrossRef]
- Shaygan, M.; Mulligan, D.; Baumgartl, T. The potential of three halophytes (Tecticornia pergranulata, Sclerolaena longicuspis, and Frankenia serpyllifolia) for the rehabilitation of brine-affected soils. Land Degrad. Dev. 2018, 29, 2002–2014. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Alam, M.M.; Bhowmik, P.C.; Hossain, M.A.; Rahman, M.M.; Vara Prasad, M.N.; Ozturk, M.; Fujita, M. Potential use of halophytes to remediate saline soils. BioMed Res. Int. 2014, 2014, 589341. [Google Scholar] [CrossRef] [PubMed]
- Jesus, J.M.; Danko, A.S.; Fiúza, A.; Borges, M.T. Phytoremediation of salt-affected soils: A review of processes, applicability, and the impact of climate change. Environ. Sci. Poll. Res. 2015, 22, 6511–6525. [Google Scholar] [CrossRef]
- Boestfleisch, C.; Wagenseil, N.B.; Buhmann, A.K.; Seal, C.E.; Wade, E.M.; Muscolo, A.; Papenbrock, J. Manipulating the antioxidant capacity of halophytes to increase their cultural and economic value through saline cultivation. AoB Plants 2014, 6, plu046. [Google Scholar] [CrossRef]
- Zheng, L.; Ma, H.; Jiao, Q.; Ma, C.; Wang, P. Phytohormones: Important participators in plant salt tolerance. Intl. J. Agric. Biol. 2020, 24, 319–332. [Google Scholar] [CrossRef]
- Ashfaq, M.; Khan, S. Role of phytohormones in improving the yield of oilseed crops. In Oils Seed Crops: Yield and Adaptation under Environmental Stress, 1st ed.; Ahmad, P., Ed.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2017; Volume 9, pp. 165–183. [Google Scholar] [CrossRef]
- Llanes, A.; Reginato, M.; Devinar, G.; Luna, V. What is known about phytohormones in halophytes? A review. Biologia 2018, 73, 727–742. [Google Scholar] [CrossRef]
- Szepesi, A. Halotropism: Phytohormonal aspects and potential applications. Front. Plant Sci. 2020, 11, 571025. [Google Scholar] [CrossRef] [PubMed]
- Berková, V.; Kameniarová, M.; Ondrisková, V.; Berka, M.; Menšiková, S.; Kopecká, R.; Luklová, M.; Novák, J.; Spichal, L.; Rashotte, A.M.; et al. Arabidopsis response to inhibitor of cytokinin degradation INCYDE: Modulations of cytokinin signaling and plant proteome. Plants 2020, 9, 1563. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015, 75, 391–404. [Google Scholar] [CrossRef]
- Marković, M.; Momcilov, T.; Uzelac, B.; Cingel, A.; Milošević, S.; Jevremović, S.; Subotić, A. Breaking the dormancy of snake’s head fritillary (Fritillary meleagris L.) in vitro bulbs-Part 1: Effect of GA3, GA inhibitors and temperature on fresh weight, sprouting and sugar content. Plants 2020, 9, 1449. [Google Scholar] [CrossRef]
- Liu, J.H.; Wang, W.; Wu, H.; Gong, X.; Moriguchi, T. Polyamines function in stress tolerance: From synthesis to regulation. Front. Plant Sci. 2015, 6, 827. [Google Scholar] [CrossRef] [PubMed]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small amines with large effects on plant abiotic stress tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef] [PubMed]
- Azad, N.; Rezayian, M.; Hassanpour, H.; Nikman, V.; Ebrahimzadeh, H. Physiological mechanisms of salicylic acid in Mentha pulegium L. under salinity and drought stress. Braz. J. Bot. 2021, 44, 359–369. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic acid in plant salinity stress signalling and tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Bueno, M.; Lendínez, M.L.; Calero, J.; Cordovilla, M.P. Salinity responses of three halophytes from inland saltmarshes of Jaén (southern Spain). Flora 2020, 266, 151589. [Google Scholar] [CrossRef]
- Consejería del Medio Ambiente. Flora Vascular de Andalucía Oriental; Junta de Andalucía: Sevilla, Spain, 2009. [Google Scholar]
- Al Hassan, M.; Pacurar, A.; López-Gresa, M.P.; Donat-Torres, M.P.; Llinares, J.V.; Boscaiu, M.; Vicente, O. Effects of salt stress on three ecologically distinct Plantago species. PLoS ONE 2016, 11, e0160236. [Google Scholar] [CrossRef]
- Koyro, H.W. Effect of salinity on growth, photosynthesis, water relations and solute compositions of the potential cash crop halophyte Plantago coronopus (L.). Environ. Exp. Bot. 2006, 56, 136–146. [Google Scholar] [CrossRef]
- Ltaeif, H.B.; Sakhraoui, A.; González-Orenga, S.; Faz, A.L.; Boscaiu, M.; Vicente, O.; Rouz, S. Responses to salinity in four Plantago species from Tunisia. Plants 2021, 10, 1392. [Google Scholar] [CrossRef] [PubMed]
- Jdey, A.; Falleh, H.; Ben Jannet, S.; Mkadmini Hammi, K.; Dauvergne, X.; Ksouri, R.; Magné, C. Phytochemical investigation and antioxidant, antibacterial and anti-tyrosinase performances for six medicinal halophytes. South Afr. J. Bot. 2017, 112, 508–514. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Karkanis, A.; Martins, N.; Ferreira, I.C. Edible halophytes of the Mediterranean basin: Potential candidates for novel food products. Trends Food Sci. Technol. 2018, 74, 69–84. [Google Scholar] [CrossRef]
- Ceccanti, C.; Brizzi, A.; Landi, M.; Incrocci, L.; Pardossi, A.; Guidi, L. Evaluation of major minerals and trace elements in wild and domesticated edible herbs traditionally used in the Mediterranean area. Biol. Trace Elem. Res. 2021, 199, 3553–3561. [Google Scholar] [CrossRef]
- Puccinelli, M.; Pezzarossa, B.; Pintimalli, L.; Malorgio, F. Selenium biofortification of three wild species, Rumex acetosa L., Plantago coronopus L., and Portulaca oleracea L., grown as microgreens. Agronomy 2021, 11, 1155. [Google Scholar] [CrossRef]
- Ceccanti, C.; Landi, M.; Incrocci, L.; Pardossi, A.; Venturi, F.; Taglieri, I.; Ferroni, G.; Guidi, L. Comparison of three domestications and wild-harvested plants for nutraceutical properties and sensory profiles in five wild edible herbs: Is domestication possible? Foods 2020, 9, 1065. [Google Scholar] [CrossRef]
- Amir, R.; Munir, F.; Khan, M.; Iqbal, T. Use of plant hormones for the improvement of plant growth and production under salt stress. In Salt Stress, Microbes, and Plant Interactions: Causes and Solution, 2nd ed.; Akhtar, M.S., Ed.; Springer Nature: Singapore, 2019; Volume 1, pp. 59–90. [Google Scholar] [CrossRef]
- Morsi, M.M.; Abdelmigid, H.M.; Aljoudi, N.G.S. Exogenous salicylic acid ameliorates the adverse effects of salt stress on antioxidant system in Rosmarinus officinalis L. Egypt. J. Bot. 2018, 58, 249–263. [Google Scholar] [CrossRef]
- Islam, M.J.; Ryu, B.R.; Azad, M.O.K.; Rahman, M.H.; Rana, M.S.; Lim, J.D.; Lim, Y.S. Exogenous putrescine enhances salt tolerance and ginsenosides content in Korena ginseng (Panax ginseng Meyer) sprouts. Plants 2021, 10, 1313. [Google Scholar] [CrossRef]
- Tounekti, T.; Hernández, I.; Müller, M.; Khemira, H.; Munné-Bosch, S. Kinetin applications alleviate salt stress and improve the antioxidant composition of leaf extracts in Salvia officinalis. Plant Physiol. Biochem. 2011, 49, 1165–1176. [Google Scholar] [CrossRef]
- Li, S.; Jin, H.; Zhang, Q. The effect of exogenous spermidine concentration on polyamine metabolism and salt tolerance in zoysiagrass (Zoysia japonica Steud) subjected to short-term salinity stress. Front. Plant Sci. 2016, 7, 1221. [Google Scholar] [CrossRef] [PubMed]
- Tiburcio, A.F.; Altabella, T.; Bitrián, M.; Alcázar, R. The roles of polyamines during the lifespan of plants: From development to stress. Planta 2014, 240, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kamiab, F.; Tavassolian, I.; Hosseinifarahi, M. Biologia Futura: The role of polyamine in plant science. Biol. Futur. 2020, 71, 183–194. [Google Scholar] [CrossRef]
- Filgueiras, C.C.; Martins, A.D.; Pereira, R.V.; Willett, D.S. The ecology of salicylic acid signaling: Primary, secondary and tertiary effects with application in agriculture. Int. J. Mol. Sci. 2019, 20, 5851. [Google Scholar] [CrossRef]
- Li, S.M.; Zheng, H.X.; Zhang, X.S.; Sui, N. Cytokinins as central regulators during plant growth and stress response. Plant Cell Rep. 2021, 40, 271–282. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine function in plants: Metabolism, regulation on development, and roles in abiotic stress responses. Front. Plant Sci. 2019, 9, 1945. [Google Scholar] [CrossRef]
- Pottosin, I.; Shabala, S. Polyamines control of cation transport across plant membranes: Implications for ion homeostasis and abiotic stress signaling. Front. Plant Sci. 2014, 5, 154. [Google Scholar] [CrossRef] [PubMed]
- Pál, M.; Szalai, G.; Janda, T. Speculation: Polyamines are important in abiotic stress signaling. Plant Sci. 2015, 237, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Ghabriche, R.; Ghnaya, T.; Mnasri, M.; Zaier, H.; Baioui, R.; Vromman, D.; Abdelly, C.; Lutts, S. Polyamine and tyramine involvement in NaCl-induced improvement of Cd resistance in the halophyte Inula crithmoides L. J. Plant Physiol. 2017, 216, 136–144. [Google Scholar] [CrossRef]
- Ruiz-Carrasco, K.; Antognoni, F.; Coulibaly, A.K.; Lizardi, S.; Covarrubias, A.; Martínez, E.A.; Molina-Montenegro, M.A.; Biondi, S.; Zurita-Silva, A. Variation in salinity tolerance of four lowland genotypes of quinoa (Chenopodium quinoa Willd.) as assessed by growth, physiological traits, and sodium transporter gene expression. Plant Physiol. Biochem. 2011, 49, 1333–1341. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Rapparini, F.; Bertazza, G.; Silva, H.; Torrigiani, P.; Biondi, S. Comparing salt-induced responses at the transcript level in a salares and coastal-lowlands landrace of quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2017, 139, 127–142. [Google Scholar] [CrossRef]
- Zarranz Elso, M.; García-Jiménez, P.; Robaina, R.R. Endogenous polyamine content and photosynthetic performance under hypo-osmotic conditions reveal Cymodocea nodosa as an obligate halophyte. Aquat. Biol. 2012, 17, 7–17. [Google Scholar] [CrossRef][Green Version]
- Gharbi, E.; Martínez, J.P.; Benahmed, H.; Fauconnier, M.L.; Lutts, S.; Quinet, M. Salicylic acid differently impacts ethylene and polyamine synthesis in the glycophyte Solanum lycopersicum and the wild-related halophyte Solanum chilense exposed to mild salt stress. Physiol. Plant 2016, 158, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Zapata, P.J.; Serrano, M.; García-Legaz, M.F.; Pretel, M.T.; Botella, M.A. Short term effect of salt shock on ethylene and polyamines depends on plant salt sensitivity. Front. Plant Sci. 2017, 8, 855. [Google Scholar] [CrossRef] [PubMed]
- Minocha, R.; Majumdar, R.; Minocha, S.C. Polyamines and abiotic stress in plants: A complex relationship. Front. Plant Sci. 2014, 5, 175. [Google Scholar] [CrossRef] [PubMed]
- Pardo-Domènech, L.L.; Tifrea, A.; Grigore, M.N.; Boscaiu, M.; Vicente, O. Proline and glycine betaine accumulation in two succulent halophytes under natural and experimental conditions. Plant Biosyst. 2016, 150, 904–915. [Google Scholar] [CrossRef]
- Pottosin, I.; Velarde-Buendía, A.M.; Bose, J.; Fuglsang, A.T.; Shabala, S. Polyamines cause plasma membrane depolarization, activate Ca2+, and modulate H+-ATPase pump activity in pea roots. J. Exp. Bot. 2014, 65, 2463–2472. [Google Scholar] [CrossRef] [PubMed]
- Pottosin, I.; Olivas-Aguirre, M.; Dobrovinskaya, O.; Zepeda-Jazo, I.; Shabala, S. Modulation of ion transport across plant membranes by polyamines: Understanding specific modes of action under stress. Front. Plant Sci. 2021, 11, 616077. [Google Scholar] [CrossRef]
- Kaur, G.; Asthir, B. Proline: A key player in plant abiotic stress tolerance. Biol. Plant. 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Ozgur, R.; Uzilday, B.; Sekmen, A.H.; Turkan, I. Reactive oxygen species regulation and antioxidant defence in halophytes. Funct. Plant Biol. 2013, 40, 832–847. [Google Scholar] [CrossRef]
- Slama, I.; Abdelly, C.; Bouchereau, A.; Flowers, T.; Savouré, A. Diversity, distribution, and roles of osmoprotective compounds accumulated in halophytes under abiotic stress. Ann. Bot. 2015, 115, 433–447. [Google Scholar] [CrossRef]
- Anwar, R.; Mattoo, A.K.; Avtar, K.H. Polyamine interactions with plant hormones: Crosstalk at several levels. In Polyamines: A Universal Molecular Nexus for Growth, Survival, and Specialized Metabolism, 1st ed.; Kusano, T., Suzuki, H., Eds.; Springer: Tokyo, Japan, 2015; pp. 267–302. [Google Scholar] [CrossRef]
- Ksouri, R.; Megdiche, W.; Falleh, H.; Trabelsi, N.; Boulaaba, M.; Smaoui, A.; Abdelly, C. Influence of biological, environmental, and technical factors on phenolic content and antioxidant activities of Tunisian halophytes. Comptes Rendus Biol. 2008, 331, 865–873. [Google Scholar] [CrossRef]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I.; Abdelly, C. Medicinal halophytes: Potent source of health promoting biomolecules withy medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.G.; Custódio, L.; Rodrigues, M.J.; Neng, N.R.; Nogueira, J.M.F.; Carlier, J.; Costa, M.C.; Varela, J.; Barreira, L. Profiling of antioxidant potential and phytoconstituents of Plantago coronopus. Braz. J. Biol. 2017, 77, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Morales, P.; Carvalho, A.M.; Ferreira, I.C.F.R. Antioxidant potential of wild plant foods. In Mediterranean Wild Edible Plants: Ethnobotany and Food Composition Tables; Springer: New York, NY, USA, 2016; pp. 209–232. ISBN 978-1-49-393329-7. [Google Scholar]
- Bueno, M.; Cordovilla, M.P. Polyamines in halophytes. Front. Plant Sci. 2019, 10, 439. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, N.E.; Tsiavos, T.; Kotzabasis, K. Chemical bonding of chlorophylls and plant aminic axial ligands impact harvesting of visible light and quenching of fluorescence. Photochem. Photobiol. 2012, 88, 98–106. [Google Scholar] [CrossRef]
- Sang, T.; Shan, X.; Li, B.; Shu, S.; Sun, J.; Guo, S. Comparative proteomic analysis reveals the positive effect of exogenous spermidine on photosynthesis and salinity tolerance in cucumber seedlings. Plant Cell Rep. 2016, 35, 1769–1782. [Google Scholar] [CrossRef]
- Wu, Z.; Wang, J.; Yan, D.; Yuan, H.; Wang, Y.; He, Y.; Wang, X.; Li, Z.; Mei, J.; Hu, M.; et al. Exogenous spermidine improves salt tolerance of pecan-grafted seedlings via activating antioxidant system and inhibiting the enhancement of Na+/K+ ratio. Acta Physiol. Plant. 2020, 42, 83. [Google Scholar] [CrossRef]
- Jiang, D.; Hou, J.; Gao, W.; Tong, X.; Li, M.; Chu, X.; Chen, G. Exogenous spermidine alleviates the adverse effects of aluminium toxicity on photosystem II through improved antioxidant system and endogenous polyamine contents. Ecotoxicol. Environ. Saf. 2021, 207, 111265. [Google Scholar] [CrossRef]
- Zhou, C.; Sun, Y.; Ma, S.; Wang, J. Heterologous expression of EsSPDS1 in tobacco plants improves drought tolerance with efficient reactive oxygen species scavenging systems. South Afr. J. Bot. 2015, 96, 19–28. [Google Scholar] [CrossRef]
- Majumdar, R.; Barchi, B.; Turlapati, S.A.; Gagne, M.; Minocha, R.; Long, S.; Minocha, S.C. Glutamate, ornithine, arginine, proline, and polyamine metabolic interactions: The pathway is regulated at the post-transcriptional level. Front. Plant Sci. 2016, 7, 78. [Google Scholar] [CrossRef] [PubMed]
- Lasanajak, Y.; Minocha, R.; Minocha, S.C.; Goyal, R.; Fatima, T.; Handa, A.K.; Mattoo, A.K. Enhanced flux of substrates into polyamine biosynthesis but not ethylene in tomato fruit engineered with yeast S-adenosylmethionine decarboxylase gene. Amino Acids 2014, 46, 729–742. [Google Scholar] [CrossRef]
- Jia, T.; Hou, J.; Iqbal, M.Z.; Zhang, Y.; Cheng, B.; Feng, H.; Li, Z.; Liu, L.; Zhou, J.; Feng, G.; et al. Overexpression of the with clover TrSAMDC1 gene enhanced salt and drought resistance in Arabidopsis thaliana. Plant Physiol. Biochem. 2021, 165, 147–160. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Bull. 1950, 347, 1–32. [Google Scholar]
- Al Hassan, M.; López-Gresa, M.P.; Boscaiu, M.; Vicente, O. Stress tolerance mechanisms in Juncus: Responses to salinity and drought in three Juncus species adapted to different natural environments. Funct. Plant Biol. 2016, 43, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Dudonné, S.; Vitrac, X.; Coutière, P.; Woillez, M.; Mérillon, J.M. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interesting using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Food Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Davies, P.J. Separation and quantification of polyamines in plant tissue by high performance liquid chromatography of their dansyl derivatives. Plant Physiol. 1985, 78, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.; Lendínez, M.L.; Aparicio, C.; Cordovilla, M.P. Effect of salinity on polyamines and ethylene in Atriplex prostrata and Plantago coronopus. Biol. Plant. 2015, 59, 596–600. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SLDW (g/plant) | RDW (g/plant) | SLWC (%) | RWC (%) |
| Control (no PGR) | 0.134 ± 0.0038 c | 0.0198 ± 0.0014 b | 94.16 ± 0.47 a | 85.07 ± 0.24 c |
| IAA | 0.171 ± 0.0056 ab | 0.0321 ± 0.0037 a | 95.45 ± 0.52 a | 92.05 ± 0.28 ab |
| Kinetin | 0.179 ± 0.0062 a | 0.0214 ± 0.0016 b | 95.67 ± 0.55 a | 90.63 ± 0.63 ab |
| GA3 | 0.147 ± 0.0052 bc | 0.0199 ± 0.0018 b | 95.51 ± 0.46 a | 89.80 ± 0.47 b |
| Spd | 0.197 ± 0.0089 a | 0.0369 ± 0.0021 a | 95.91 ± 0.56 a | 92.88 ± 0.66 a |
| SA | 0.186 ± 0.0063 a | 0.0341 ± 0.0012 a | 95.74 ± 0.39 a | 92.53 ± 0.45 a |
| SLDW (g/plant) | RDW (g/plant) | SLWC (%) | RWC (%) |
| Control (salt) | 0.080 ± 0.0110 d | 0.0101 ± 0.0005 d | 91.54 ± 0.35 b | 82.63 ± 0.33 c |
| IAA + salt | 0.132 ± 0.0078 bc | 0.0167 ± 0.0005 b | 92.50 ± 0.38 b | 85.54 ± 0.38 ab |
| Kinetin + salt | 0.161 ± 0.0064 b | 0.0128 ± 0.0004 c | 93.18 ± 0.42 b | 83.55 ± 0.90 bc |
| GA3 + salt | 0.085 ± 0.0063 d | 0.0117 ± 0.0006 cd | 92.11 ± 0.52 b | 83.09 ± 0.56 bc |
| Spd + salt | 0.219 ± 0.0100 a | 0.0300 ± 0.0058 a | 95.72 ± 0.45 a | 87.55 ± 0.40 a |
| SA + salt | 0.092 ± 0.0090 cd | 0.0094 ± 0.0008 d | 92.50 ± 0.44 b | 82.43 ± 0.70 c |
| PGRs Application with Salt | Total Phenols (mg GAE g−1 DW) | Total Flavonoids (mg CE g−1 DW) |
|---|---|---|
| Control (Salt) | 4.5 ± 0.11 c | 3.1 ± 0.11 b |
| IAA + salt | 4.9 ± 0.12 bc | 3.5 ± 0.12 ab |
| Kinetin + salt | 5.3 ± 0.10 ab | 3.7 ± 0.23 ab |
| GA3 + salt | 4.6 ± 0.17 bc | 2.9 ± 0.23 b |
| Spd + salt | 5.9 ± 0.21 a | 4.0 ± 0.14 a |
| SA + salt | 5.0 ± 0.20 bc | 3.2 ± 0.20 b |
| Total PAs (nmol g−1 DW) | Ethylene (nL g−1 FW h−1) |
| Control (no PGRs) | 21.87 ± 0.68 d | 10.30 ± 0.64 a |
| IAA | 24.54 ± 0.75 d | 8.35 ± 0.61 ab |
| Kinetin | 46.00 ± 1.81 b | 5.62 ± 0.71 b |
| GA3 | 25.01 ± 0.88 d | 7.68 ± 0.67 ab |
| Spd | 61.29 ± 1.45 a | 6.35 ± 0.53 b |
| SA | 36.50 ± 0.69 c | 5.81 ± 0.65 b |
| Total PAs (nmol g−1 DW) | Ethylene (nL g−1 FW h−1) |
| Control (salt) | 32.11 ± 0.58 c | 7.61 ± 0.52 a |
| IAA + salt | 49.94 ± 0.42 b | 3.30 ± 0.68 b |
| Kinetin + salt | 54.70 ± 2.42 b | 3.77 ± 0.47 b |
| GA3 + salt | 36.76 ± 1.16 c | 5.23 ± 0.59 ab |
| Spd + salt | 67.79 ± 2.54 a | 3.74 ± 0.64 b |
| SA + salt | 37.50 ± 1.94 c | 5.76 ± 0.53 ab |
| SLDW | RDW | SLWC | RWC | SOR | PUT | SPD | SPM | Total PAs | C2H2 | |
|---|---|---|---|---|---|---|---|---|---|---|
| SLDW | 1 | |||||||||
| RDW | 0.7522 ** | 1 | ||||||||
| SLWC | 0.7629 ** | 0.7559 ** | 1 | |||||||
| RWC | 0.7049 ** | 0.8769 ** | 0.7941 ** | 1 | ||||||
| SOR | 0.0723 | −0.2711 | −0.2874 | −0.440 ** | 1 | |||||
| PUT | 0.4624 ** | 0.4943 ** | 0.3212 | 0.4720 ** | −0.0267 | 1 | ||||
| SPD | 0.6824 ** | 0.4669 ** | 0.3550 * | 0.3892 * | 0.3662 * | 0.7729 ** | 1 | |||
| SPM | 0.1200 * | −0.3134 | −0.3501 * | −0.470 ** | 0.8465 ** | −0.0308 | 0.4360 ** | 1 | ||
| Total PAs | 0.5148 ** | 0.2064 | 0.0816 * | 0.0840 | 0.5897 ** | 0.6838 ** | 0.9193 ** | 0.7184 ** | 1 | |
| C2H2 | −0.2150 | 0.1044 | 0.2573 | 0.1496 | −0.659 ** | -0.2616 | −0.588 ** | −0.732 ** | −0.723 ** | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bueno, M.; Cordovilla, M.d.P. Plant Growth Regulators Application Enhance Tolerance to Salinity and Benefit the Halophyte Plantago coronopus in Saline Agriculture. Plants 2021, 10, 1872. https://doi.org/10.3390/plants10091872
Bueno M, Cordovilla MdP. Plant Growth Regulators Application Enhance Tolerance to Salinity and Benefit the Halophyte Plantago coronopus in Saline Agriculture. Plants. 2021; 10(9):1872. https://doi.org/10.3390/plants10091872
Chicago/Turabian StyleBueno, Milagros, and María del Pilar Cordovilla. 2021. "Plant Growth Regulators Application Enhance Tolerance to Salinity and Benefit the Halophyte Plantago coronopus in Saline Agriculture" Plants 10, no. 9: 1872. https://doi.org/10.3390/plants10091872
APA StyleBueno, M., & Cordovilla, M. d. P. (2021). Plant Growth Regulators Application Enhance Tolerance to Salinity and Benefit the Halophyte Plantago coronopus in Saline Agriculture. Plants, 10(9), 1872. https://doi.org/10.3390/plants10091872