
In Vitro Propagation and Acclimatization of Banana Plants: Antioxidant Enzymes, Chemical Assessments and Genetic Stability of Regenerates as a Response to Copper Sulphate

 , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
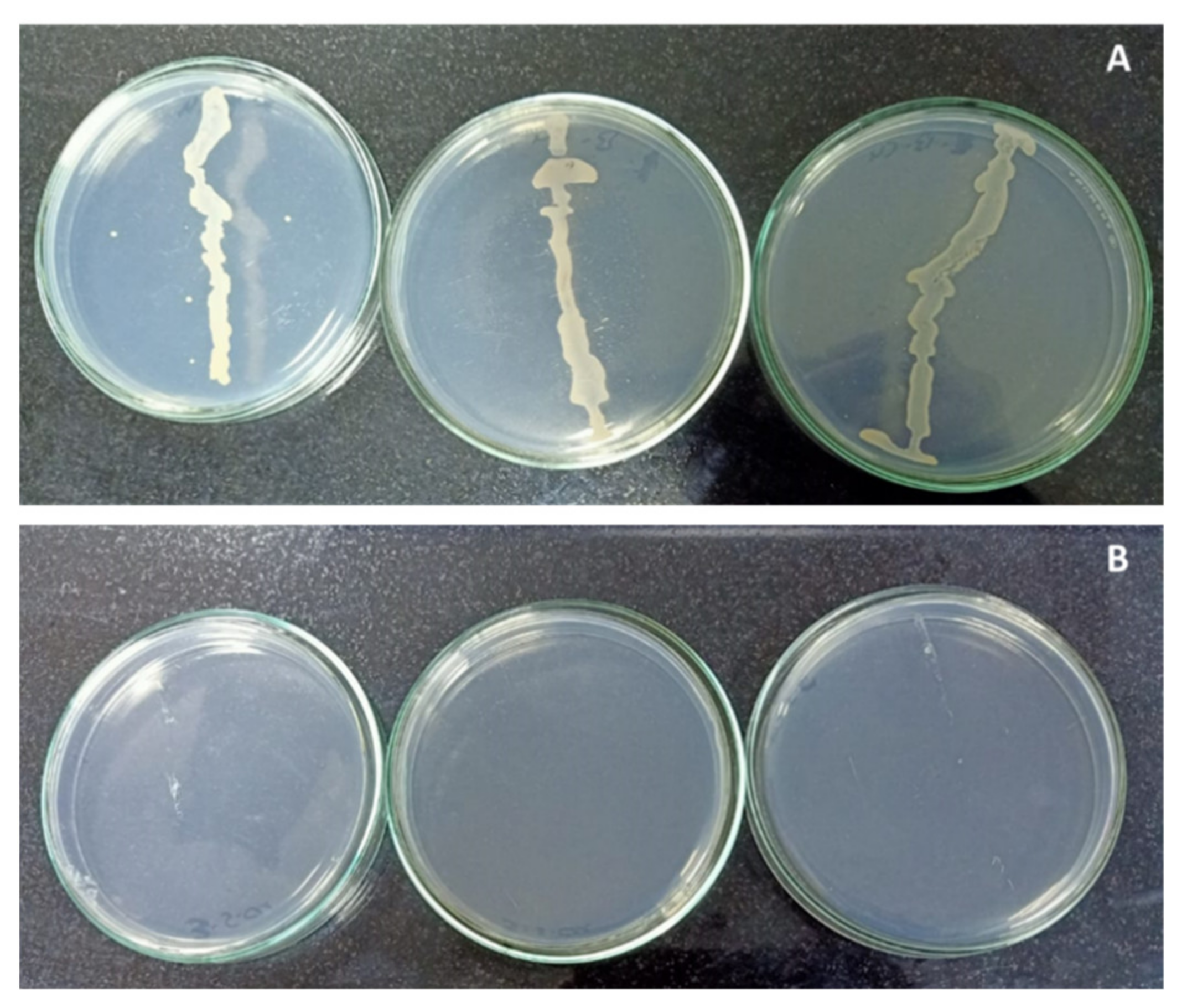
2.1. Copper Sulphate and Endophytic Bacterial Contamination Percentage
2.2. Copper Sulphate and Shoot Multiplication
2.3. Copper Sulphate and In Vitro Rooting
2.4. Copper Sulphate and Acclimatization
2.5. Copper Sulphate and Chemical Analyses
2.5.1. Copper Sulphate and Mineral Content of In Vitro Plantlets
2.5.2. Enzymatic Antioxidant Activities and Electrolyte Leakage
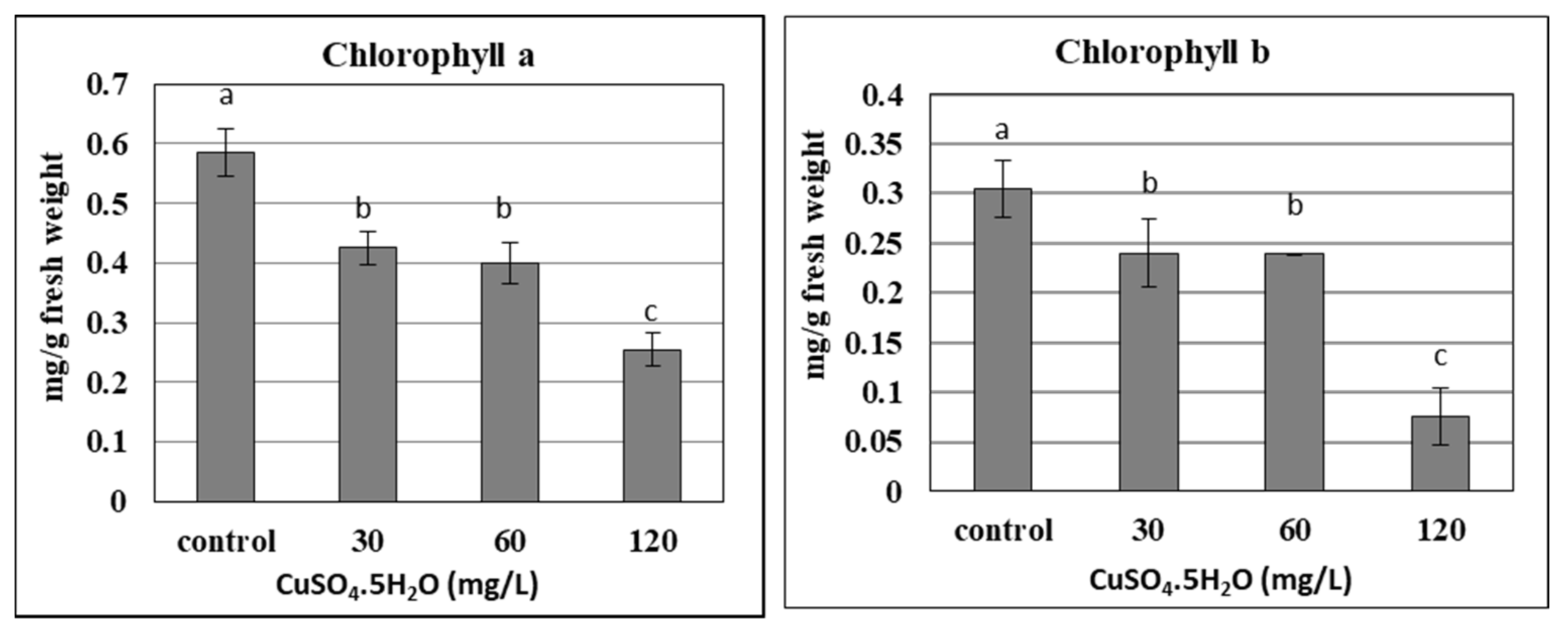
2.5.3. Copper Sulphate and Chlorophyll Content of In Vitro Plantlets
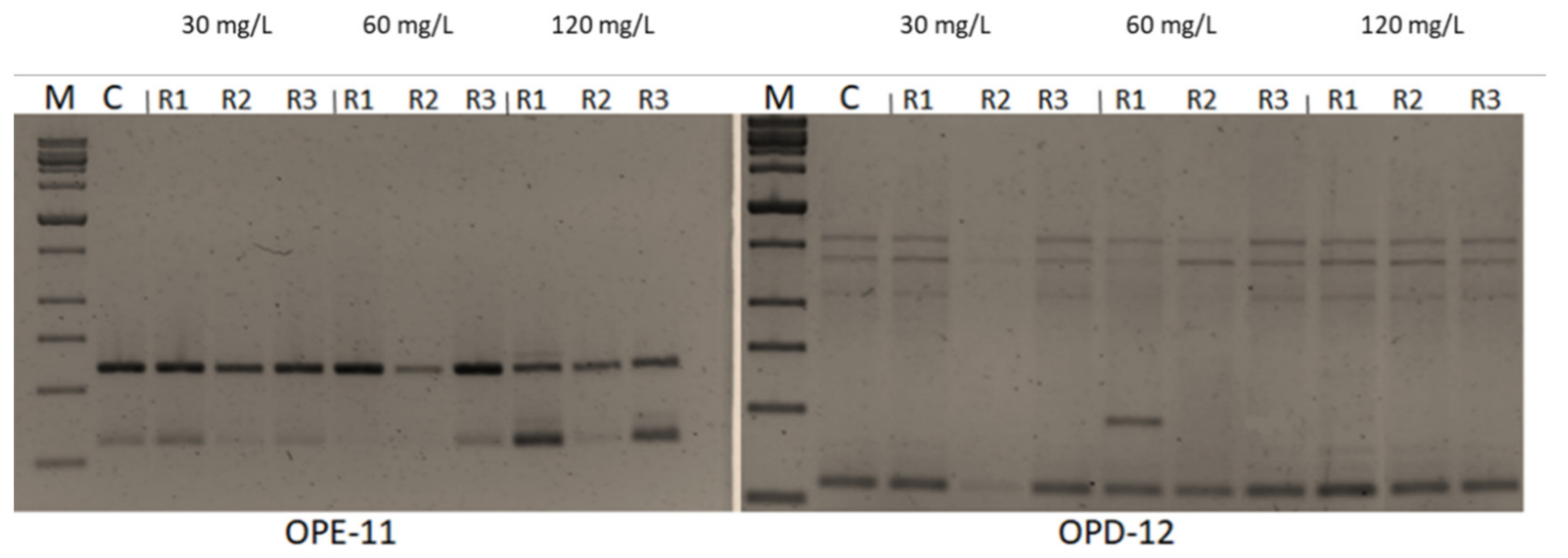
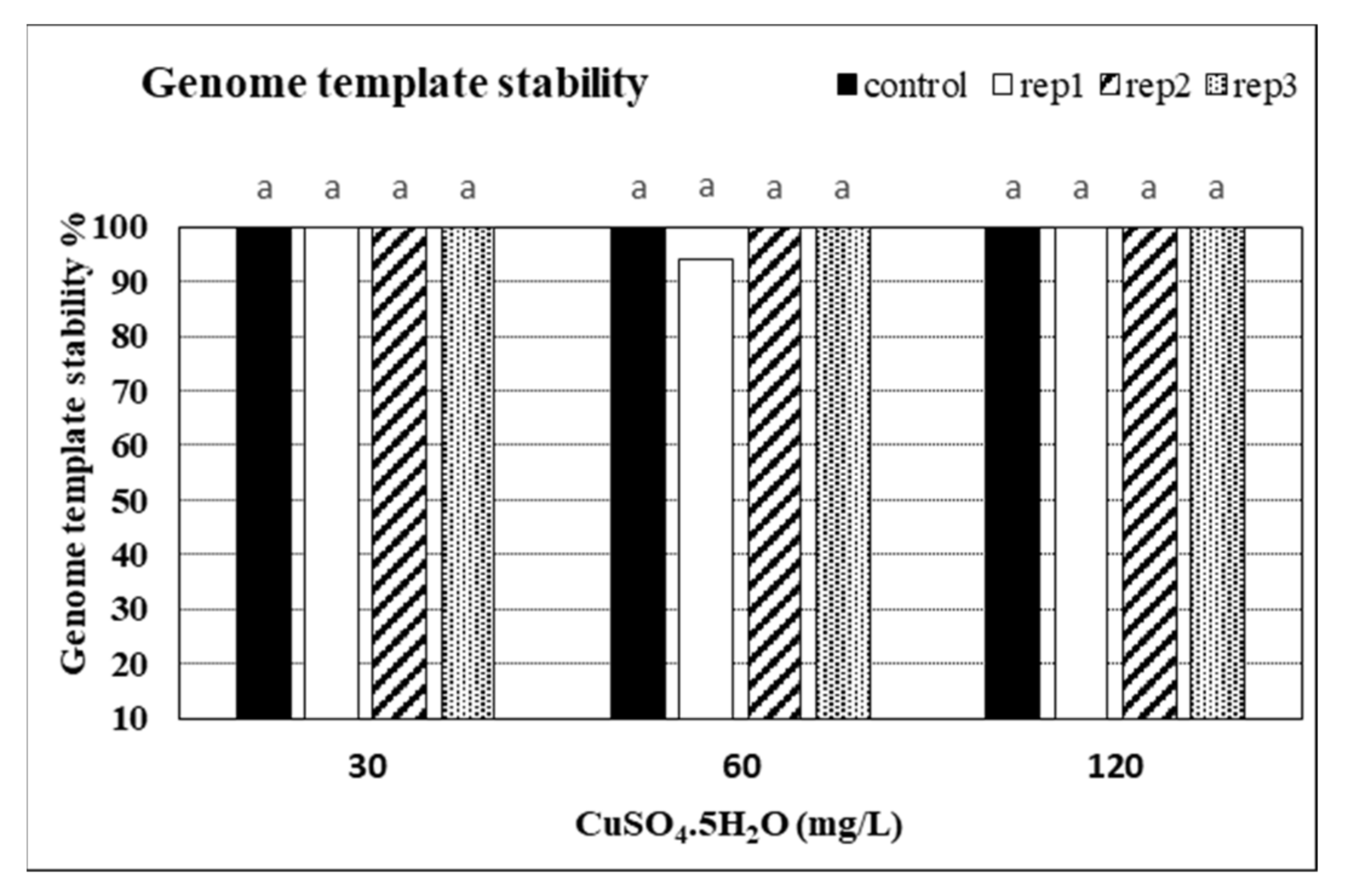
2.6. Impacts of CuSO4 H2O Treatments on Genetic Fidelity and Genomic Template Stability
3. Materials and Methods
3.1. Plant Materials
3.2. Copper Sulphate and Bacterial Contamination (Primary Experiment)
3.3. Copper Sulphate, Shoot Multiplication and In Vitro Rooting
3.4. Copper Sulphate and Acclimatization
3.5. Chemical Analyses
3.5.1. Enzymatic Antioxidants and Electrolyte Leakage
3.5.2. Measuring of Chlorophyll Contents
3.5.3. Chemical Composition of In Vitro Plantlets
3.6. Plant DNA Extraction and RAPD-PCR Conditions
3.7. Genomic Template Stability and Its Estimation
3.8. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hasan, A.S.; Khasim, S.M.; Ramudu, J. Development of standard protocols for in vitro regeneration of some selected banana cultivars (Musa spp.) from India. In Medicinal Plants: Biodiversity, Sustainable Utilization and Conservation; Khasim, S.M., Long, C., Thammasiri, K., Lutken, H., Eds.; Springer Nature: Singapore, 2020; Volume 45, pp. 743–759. [Google Scholar] [CrossRef]
- Dagnew, A.; Shibru, S.; Debebe, A.; Lemma, A.; Dessalegn, L.; Berhanu, B.; Beyene, W.; Sierra, Y.M. Micropropagation of banana varieties (Musa spp.) using shoot tip culture. Ethiop. J. Agric. Sci. 2012, 22, 14–25. [Google Scholar]
- Bhatia, S.; Sharma, K.; Dahiya, R.; Bera, T. Modern Applications of Plant Biotechnology in Pharmaceutical Sciences; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Mahmoud, L.M.; Dutt, M.; Shalan, A.M.; El-Kady, M.E.; El-Boray, M.S.; Shabana, Y.M.; Grosser, J.W. Silicon nanoparticles mitigate oxidative stress of in vitro-derived banana (Musa acuminata ‘Grand Nain’) under simulated water deficit or salinity stress. S. Afr. J. Bot. 2020, 132, 155–163. [Google Scholar] [CrossRef]
- Seliem, M.K.; El-Mahrouk, M.E.; El-Banna, A.N.; Hafez, Y.M.; Dewir, Y.H. Micropropagation of Philodendron selloum: Influence of copper sulphate on endophytic bacterial contamination, antioxidant enzyme activity, electrolyte leakage, and plant survival. S. Afr. J. Bot. 2021, 139, 230–240. [Google Scholar] [CrossRef]
- Javed, S.B.; Alatar, A.A.; Basahi, R.; Anis, M.; Faisal, M.; Husain, F.M. Copper induced suppression of systemic microbial contamination in Erythrina variegata L. during in vitro culture. Plant Cell Tissue Organ Cult. 2017, 128, 249–258. [Google Scholar] [CrossRef]
- Broadley, M.; Brown, P.; Cakmak, I.; Rengel, Z.; Zhao, F. Chapter 7—Function of nutrients: Micronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: London, UK, 2012; Volume 89, pp. 191–248. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Osdaghi, E.; Behlau, F.; Kohl, J.; Jones, J.B.; Aubertot, J. Thirteen decades of antimicrobial copper compounds applied in agriculture: A review. Agron. Sustain. Dev. 2018, 38, 1–18. [Google Scholar] [CrossRef]
- Hafez, Y.M.; Salama, A.; Kotb, H.; Moussa, Z.; Elsaed, N.; El-Kady, E.M.; Hassan, F.A.S. The influence of nano-copper and safety Compounds on vegetative growth, yield and fruit quality of “Le Conte” pear trees under infection with fire blight. Fresenius Environ. Bull. 2021, 30, 6237–6247. [Google Scholar]
- Chen, J.; Shafi, M.; Li, S.; Wang, Y.; Wu, J.; Ye, Z.; Peng, D.; Yan, W.; Liu, D. Copper induced oxidative stresses, antioxidant responses and phytoremediation potential of Moso bamboo (Phyllostachys pubescens). Sci. Rep. 2015, 5, 13554. [Google Scholar] [CrossRef]
- Cho, M.J.; Lemaux, P.G.; Jiang, W. Transformation of recalcitrant barley cultivars through improvement of regenerative ability and decreased albinism. Plant Sci. 1998, 138, 229–244. [Google Scholar] [CrossRef]
- Gori, P.; Schiff, S.; Santandrea, G.; Bennici, A. Response of in vitro cultures of Nicotiana tabacum L. to copper stress and selection of plants from Cu-tolerant callus. Plant Cell Tissue Organ Cult. 1998, 53, 161–169. [Google Scholar] [CrossRef]
- Saba, S.; Pande, D.; Iqbal, M.; Srivastava, P.S. Effect of ZnSO4 and CuSO4 on regeneration and lepidine content in Lepidium sativum L. Biol. Plant 2000, 43, 253–256. [Google Scholar] [CrossRef]
- Amarasinghe, A.A.Y. Effects of copper sulphate and cobalt chloride on in vitro performances of traditional indica rice (Oryza sativa L.) varieties in Sri Lanka. J. Agric. Sci. 2009, 4, 132–141. [Google Scholar] [CrossRef]
- Kowalska, U.; Szafranska, K.; Krzy Zanowska, D.; Kiszczak, W.; Gorecki, R.; Janas, K.; Gorecka, K. Effect of increased copper ion content in the medium on the regeneration of androgenetic embryos of carrot (Daucus carota L.). Acta Agrobot. 2012, 65, 73–82. [Google Scholar] [CrossRef]
- Chung, I.; Rajakumar, G.; Subramanian, U.; Venkidasamy, B.; Thiruvengadam, M. Impact of copper oxide nanoparticles on enhancement of bioactive compounds using cell suspension cultures of Gymnema sylvestre (Retz). R. Br. Appl. Sci. 2019, 9, 2165. [Google Scholar] [CrossRef]
- Deo, B.; Nayak, P.K. Study of copper phytotoxicity on in vitro culture of Musa acuminata cv. ‘Bantala’. J. Agric. Biotechnol. Sustain. Dev. 2011, 3, 136–140. [Google Scholar]
- Pisam, W.M.; Abdel Aal, A.H.; Khodair, O.A.; Youssef, M. In vitro performance of two banana cultivars under copper stress. Assiut J. Agric. Sci. 2020, 51, 143–153. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; Mahmood, M.; Meon, S.; Wahab, Z.; Ghobadi, C. Evaluation of in vitro water stress tolerance among EMS—induced variants of banana (Musa spp., AAA), using “Morphological, Physiological and Molecular” traits. J. Crop Sci. Biotech. 2011, 14, 255–263. [Google Scholar] [CrossRef]
- Sinha, S.K.; Deka, A.C. Effect of osmotic stress on in vitro propagation of Musa sp. (Malbhog variety). Afr. J. Biotechnol. 2016, 15, 490–496. [Google Scholar] [CrossRef][Green Version]
- El-Mahdy, M.T.; Youssef, M.; Eissa, M.A. Impact of in vitro cold stress on two banana genotypes based on physio-biochemical evaluation. S. Afr. J. Bot. 2018, 119, 219–225. [Google Scholar] [CrossRef]
- Do, G.D.; Dang, T.K.T.; Nguyen, T.H.T.; Nguyen, T.D.; Tran, T.T.; Duong, D.H. Effects of nano silver on the growth of banana (Musa spp.) cultured in vitro. J. Viet. Environ. 2018, 10, 92–98. [Google Scholar] [CrossRef]
- El-Mahdy, M.T.; Radi, A.A.; Shaaban, M.M. Impacts of exposure of banana to silver nanoparticles and sliver ions in vitro. Middle East J. Appl. Sci. 2019, 9, 727–740. [Google Scholar]
- Huong, B.T.T.; Xuan, T.D.; Trung, K.H.; Ha, T.T.T.; Duong, V.X.; Khanh, T.D.; Gioi, D.H. Influences of silver nanoparticles in vitro morphogenesis of specialty king banana (Musa ssp.) in Vietnam. Plant Cell Biotechnol. Mol. Biol. 2021, 22, 163–175. Available online: https://www.ikprress.org/index.php/PCBMB/article/view/6355 (accessed on 2 September 2021).
- Tamimi, S.M.; Othman, H. Effects of copper sulphate on shoot multiplication and rooting of banana (Musa acuminata L.) (In vitro Study). Asian J. Plant Sci. 2020, 19, 200–204. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth & bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar]
- Faúndez, G.; Troncoso, M.; Navarrete, P.; Figuero, G. Antimicrobial activity of copper surfaces against suspensions of Salmonella enterica and Campylobacter jejuni. BMC Microbiol. 2004, 4, 19. [Google Scholar] [CrossRef] [PubMed]
- Noyce, J.O.; Michels, H.; Keevil, C.W. Inactivation of influenza A virus on copper vs stainless steel. Appl. Environ. Microbiol. 2007, 73, 2748–2750. [Google Scholar] [CrossRef] [PubMed]
- Grass, G.; Rensing, C.; Solioz, M. Metallic copper as an antimicrobial surface. Appl. Environ. Microbiol. 2011, 77, 1541–1547. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S. Molecular mechanisms of plant metal tolerance and homeostasis. Planta 2001, 212, 475–486. [Google Scholar] [CrossRef]
- Prazak, R.; Prazak, R.; Molas, J. Effect of copper concentration on micropropagation and accumulation of some metals in the Dendrobium kingianum Bidwill Orchid. J. Elem. 2015, 20, 693–703. [Google Scholar]
- Ibrahim, S.M.; Hashish, K.I.; Taha, L.S.; Mazhar, A.A.; Kandil, M.M. In vitro culture protocol, micropropagation, acclimatization and chemical constituents of Spathiphyllum cannifolium plant under copper concentration effect. Int. J. PharmTech Res. 2016, 9, 33–41. [Google Scholar]
- Fatima, N.; Ahmad, N.; Anis, M. Enhanced in vitro regeneration and change in photosynthetic pigments, biomass and proline content in Withania somnifera L. (Dunal) induced by copper and zinc ions. Plant Physiol. Biochem. 2011, 49, 1465–1471. [Google Scholar] [CrossRef]
- Kumar, R.; Mehrotra, N.K.; Nautiyal, B.D.; Kumar, P.; Singh, P.K. Effect of copper on growth, yield and concentration of Fe, Mn, Zn and Cu in wheat plants (Triticum aestivum L.). J. Environ. Biol. 2009, 30, 485–488. [Google Scholar]
- Shabbir, Z.; Sardar, A.; Shabbir, A.; Abbas, G.; Shamshad, S.; Khalid, S.; Natasha, M.G.; Dumat, C.; Shahid, M. Copper uptake, essentiality, toxicity, detoxification and risk assessment in soil-plant environment. Chemosphere 2020, 259, 127436. [Google Scholar] [CrossRef]
- Aguirre, G.; Pilon, M. Copper delivery to chloroplast proteins and its regulation. Front. Plant Sci. 2016, 6, 1250. [Google Scholar] [CrossRef]
- Printz, B.; Lutts, S.; Hausman, J.F.; Sergeant, K. Copper trafficking in plants and its implication on cell wall dynamics. Front. Plant Sci. 2016, 7, 601. [Google Scholar] [CrossRef]
- Ameh, T.; Sayes, C.M. The potential exposure and hazards of copper nanoparticles: A review. Environ. Toxicol. Pharmacol. 2019, 71, 103220. [Google Scholar] [CrossRef] [PubMed]
- Gong, Q.; Wang, L.; Dai, T.; Zhou, J.; Kang, Q.; Chen, H.; Li, K.; Li, Z. Effects of copper on the growth, antioxidant enzymes and photosynthesis of spinach seedlings. Ecotoxicol. Environ. Saf. 2019, 171, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, D.; Singh, S.; Kumar, V.; Romero, R.; Prasad, R.; Singh, J. Antioxidant enzymes regulation in plants in reference to reactive oxygen species (ROS) and reactive nitrogen species (RNS). Plant Gene 2019, 19, 100182. [Google Scholar] [CrossRef]
- Rustagi, A.; Jain, S.; Kumar, D.; Shekhar, S.; Jain, M.; Bhat, V.; Sarin, N.B. High Efficiency transformation of banana [Musa acuminata L. cv. Matti (AA)] for enhanced tolerance to salt and drought stress through overexpression of a peanut salinity-induced pathogenesis-related class 10 protein. Mol. Biotechnol. 2015, 57, 27–35. [Google Scholar] [CrossRef] [PubMed]
- El-Mahrouk, M.E.; El-Shereif, A.R.; Dewir, Y.H.; Hafez, Y.M.; Abdelaal, K.A.; El-Hendawy, S.; Migdadi, H.; Al-Obeed, R.S. Micropropagation of banana: Reversion, rooting, and acclimatization of hyperhydric shoots. HortScience 2019, 54, 1384–1390. [Google Scholar] [CrossRef]
- Asmar, S.A.; Soares, J.D.R.; Silva, R.A.L.; Pasqual, M.; Pio, L.A.S.; de Castro, E.M. Anatomical and structural changes in response to application of silicon (Si) in vitro during the acclimatization of banana cv. ‘Grand Naine’. Aust. J. Crop Sci. 2015, 9, 1236–1241. [Google Scholar]
- El-Banna, A.; Khatab, I. Assessing genetic diversity of some potato (Solanum tuberosum L.) cultivars by protein and RAPD markers. Egypt. J. Genet. Cytol. 2013, 42, 89–101. [Google Scholar] [CrossRef]
- El-Banna, N.A.; El-Mahrouk, E.M.; El-Denary, E.M.; Dewir, H.Y.; Naidoo, Y. Genetic relationship and diversity in some ornamental palms based on proteins and randomly amplified polymorphic DNA markers. Hortscience 2017, 52, 338–342. [Google Scholar] [CrossRef]
- Salama, D.; Osman, S.; Abd El-Aziz, M.; Abd Elwahed, M.S.A.; Shaaban, E.A. Effect of zinc oxide nanoparticles on the growth, genomic DNA, production and the quality of common dry bean (Phaseolus vulgaris). Biocatal. Agric Biotechnol. 2019, 18, 1–11. [Google Scholar] [CrossRef]
- Mattiello, A.; Filippi, A.; Pošćić, F.; Musetti, R.; Salvatici, M.C.; Giordano, C.; Vischi, M.; Bertolini, A.; Marchiol, L. Evidence of phytotoxicity and genotoxicity in Hordeum vulgare L. exposed to CeO2 and TiO2 nanoparticles. Front. Plant Sci. 2015, 6, 1043. [Google Scholar] [CrossRef]
- Ghosh, M.; Ghosh, I.; Godderis, L.; Hoet, P.; Mukherjee, A. Genotoxicity of engineered nanoparticles in higher plants. Mutat. Res. 2019, 842, 132–145. [Google Scholar] [CrossRef]
- Al-Quraidi, A.O.; Mosa, K.A.; Ramamoorthy, K. Phytotoxic and genotoxic effects of copper nanoparticles in coriander (Coriandrum sativum—apiaceae). Plants 2019, 8, 19. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Jayaraj, M.; Manikandan, R.; Geetha, N.; Rene, E.R.; Sharma, N.C.; Sahi, S.V. Zinc oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: A physiochemical analysis. Plant Physiol. Biochem. 2017, 110, 59–69. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, R.; Nuckles, E.M.; Kuc, J. Association of enhanced peroxidase activity with induced systemic resistance of cucumber to Colletotrichum lagenarium. Physiol. Plant Pathol. 1982, 20, 73–82. [Google Scholar] [CrossRef]
- Malik, C.P.; Singh, M.B. Plant Emymology and Histoenzymology; Kalyani Publishers: Delhi, India, 1980. [Google Scholar]
- Szalai, G.; Janda, T.; Paldi, E.; Szigeti, Z. Role of light in the development of post-chilling symptoms in maize. J. Plant Physiol. 1996, 148, 378–383. [Google Scholar] [CrossRef]
- Whitlow, T.H.; Bassuk, N.L.; Ranney, T.G.; Reichert, D.L. An improved method for using electrolyte leakage to assess membrane competence in plant tissues. Plant Physiol. 1992, 98, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Dewir, Y.H.; El-Mahrouk, M.E.; AL-Shmgani, H.S.; Rihan, H.Z.; Teixeira Da Silva, J.A.; Fuller, M.P. Photosynthetic and biochemical characterization of in vitro-derived African violet (Saintpaulia ionantha H. Wendl) plants to ex vitro conditions. J. Plant Interact. 2015, 10, 101–108. [Google Scholar] [CrossRef]
- Lichtenthaler, K.; Welburn, A.R. Determination of total carotenoids and chlorophylls A and B of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Page, A.L.; Miller, R.H.; Keeny, D.R. Methods of Soil Analysis, Part II, 2nd ed.; Agronomy Monographs; Madison Book Company: Madison, WI, USA, 1982. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments (mg L−1) | 1 Contamination (%) | 2 Number of Shoots | 2 Number of Leaves/Shoots | 2 Shoot Length (cm) |
|---|---|---|---|---|
| Control | 70.0 a ± 0.176 | 10 a ± 0.192 | 4.0 ± 0.176 | 6.0 a ± 0.311 |
| 30 | 5.0 b ± 0.269 | 11 a ± 0.101 | 4.0 ± 0.117 | 5.7 a ± 0.294 |
| 60 | 0.0 c | 8 b ± 0.151 | 3.0 ± 0.212 | 4.0 b ± 0.176 |
| 120 | 0.0 c | 4 c ± 0.135 | 3.0 ± 0.117 | 2.5 b ± 0.173 |
| Significance | * | ** | NS | ** |
| Treatments (mg L−1) | 2 Plantlet Length (cm) | 2 Fresh Weight (g) | 2 Number of Leaves | 1 Roots Number | 1 Root Length (cm) |
|---|---|---|---|---|---|
| Control | 10.40 b ± 0.203 | 1.60 b ± 0.117 | 5.80 a ± 0.046 | 3.40 ab ± 0.058 | 7.78 ab ±0.196 |
| 30 | 14.26 a ± 0.152 | 2.47a ± 0.058 | 6.00 a ± 0.188 | 3.80 a ± 0.091 | 9.92 a ±0.135 |
| 60 | 11.92 b ± 0.367 | 1.38c ± 0.070 | 4.40 b ± 0.128 | 3.80 a ± 0.088 | 7.75 ab ± 0.155 |
| 120 | 10.28 b ± 0.164 | 0.61d ± 0.066 | 4.20 b ± 0.119 | 2.20 b ± 0.155 | 3.05 b ± 0.052 |
| Significance | * | ** | ** | * | * |
| Treatments (mg L−1) | 1 Survival Percent (%) | 2 Plant Length (cm) | 2 Number of Leaves | 2 Number of Roots | 2 Root Length (cm) | 1 Leaf Area (cm2) | 2 Fresh Weight (g) |
|---|---|---|---|---|---|---|---|
| Control | 100 a (±0.470) | 21.55 b (±0.421) | 6.00 ab (±0.529) | 5.33 a (±0.208) | 10.75 b (±0.327) | 46.06 a (±0.317) | 3.28 b (±0.225) |
| 30 | 100 a (±0.588) | 23.83 a (±0.503) | 6.75 a (±0.123) | 4.50 a (±0.257) | 12.67 a (±0.241) | 59.23 a (±0.169) | 5.67 a (±0.200) |
| 60 | 70 ab (±0.203) | 14.75 c (±0.500) | 5.25 c (±0.269) | 2.75 b (±0.306) | 8.00 c (±0.294) | 35.42 ab (±0.268) | 1.84 c (±0.205) |
| 120 | 50 b (±0.561) | 12.00 d (±0.212) | 4.33 d (±0.172) | 2.33 b (±0.195) | 7.50 c (±0.156) | 32.50 b (±0.094) | 0.71 c (±0.150) |
| Significance | * | ** | ** | ** | ** | * | ** |
| Treatments (mg L−1) | 1 K (%) | 1 Ca (%) | 2 P (%) | 1 Mg (%) | 1 Fe (mg kg−1) | 2 Mn (mg kg−1) | 2 Cu (mg kg−1) | 2 Zn (mg kg−1) |
|---|---|---|---|---|---|---|---|---|
| Control | 0.205 ab (±0.003) | 0.701 b (±0.06) | 0.165 d (±0.004) | 0.089 c (±0.006) | 229 b (±0.588) | 125 d (±2.941) | 1.38 d (±0.088) | 44.1 d (±0.235) |
| 30 | 0.142 b (±0.004) | 0.832 ab (±0.005) | 0.305 a (±0.003) | 0.104 bc (±0.003) | 220 b (±2.941) | 212 c (±0.1.176) | 14.88 c (±0.285) | 72.5 b (±0.294) |
| 60 | 0.548 a (±0.014) | 1.253 a (±0.055) | 0.231 c (±0.022) | 0.249 a (±0.029) | 341 a (±2.352) | 398 a (±2.695) | 23.97 b (±0.203) | 103.3 a (±0.411) |
| 120 | 0.244 ab (±0.049) | 0.638 b (±0.023) | 0.288 b (±0.024) | 0.128 ab (±0.004) | 394 a (±2.695) | 315 b (±2.941) | 44.69 a (±0.411) | 64.6 c (±0.459) |
| Treatments (mg L−1) | 1 K (%) | 1 Ca (%) | 2 P (%) | 2 Mg (%) | 2 Fe (mg kg−1) | 2 Mn (mg kg−1) | 2 Cu (mg kg−1) | 1 Zn (mg kg−1) |
|---|---|---|---|---|---|---|---|---|
| Control | 3.52 ab (±0.011) | 0.290 a (±0.002) | 0.138 c (±0.001) | 0.382 a (±0.004) | 293 a (±1.018) | 153 b (±1.556) | 1.70 c (±0.042) | 124 bc (±0.481) |
| 30 | 3.82 a (± 0.071) | 0.269 ab (±0.001) | 0.169 a (±0.005) | 0.359 b (±0.009) | 204 c (±1.176) | 169 a (±1.176) | 0.23 d (±0.013) | 169 a (±0.444) |
| 60 | 3.63 a (±0.069) | 0.255 bc (±0.003) | 0.150 b (±0.002) | 0.272 c (±0.01) | 245 b (±0.588) | 136 c (±1.060) | 3.14 b (±0.012) | 126 ab (±0.561) |
| 120 | 0.160 b (±0.018) | 0.169 c (±0.002) | 0.133 d (±0.003) | 0.169 d (±0.003) | 169 d (±1.176) | 126 d (±1.020) | 5.33 a (±0.051) | 116 c (±0.444) |
| Primers | Sequence (5′→3′) | CuSO4 mg L−1 | No. of RAPD Markers | No. of Polymorphic Bands | Average of Polymorphism% | ||
|---|---|---|---|---|---|---|---|
| R1 | R2 | R3 | |||||
| OPE-11 | GAGTCTCAGG | 30 | 2 | 2 | 2 | 0 | 0 |
| 60 | 2 | 2 | 2 | 0 | |||
| 120 | 2 | 2 | 2 | 0 | |||
| OPD-12 | CACCGTATCC | 30 | 5 | 5 | 5 | 0 | 1.8 |
| 60 | 6 | 5 | 5 | 1 | |||
| 120 | 5 | 5 | 5 | 0 | |||
| OPD-14 | CTTCCCCAAG | 30 | 7 | 7 | 7 | 0 | 0 |
| 60 | 7 | 7 | 7 | 0 | |||
| 120 | 7 | 7 | 7 | 0 | |||
| OPH-01 | GGTCGGAGAA | 30 | 3 | 3 | 3 | 0 | 0 |
| 60 | 3 | 3 | 3 | 0 | |||
| 120 | 3 | 3 | 3 | 0 | |||
| OPD-08 | GTGTGCCCCA | 30 | 3 | 3 | 3 | 0 | 0 |
| 60 | 3 | 3 | 3 | 0 | |||
| 120 | 3 | 3 | 3 | 0 | |||
| OPK-08 | GAACACTGGG | 30 | 4 | 4 | 4 | 0 | 0 |
| 60 | 4 | 4 | 4 | 0 | |||
| 120 | 4 | 4 | 4 | 0 | |||
| OPE-12 | TTATCGCCCC | 30 | 6 | 6 | 6 | 0 | 0 |
| 60 | 6 | 6 | 6 | 0 | |||
| 120 | 6 | 6 | 6 | 0 | |||
| CuSO4 mg L−1 | Replicates | Band Profile | Primers | Total Bands | a + b | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| OPD12 | OPD14 | OPH01 | OPD08 | OPK08 | OPE11 | OPE12 | |||||
| Control | - | 5 | 7 | 3 | 3 | 4 | 2 | 6 | 30 | - | |
| 30 | 1 | a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 2 | a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 3 | a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 60 | 1 | a | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 2 | a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 3 | a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 120 | 1 | a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 2 | a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| 3 | a | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| b | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |||
| Total No. of bands per primer | 6 | 7 | 3 | 3 | 4 | 2 | 6 | ||||
| The Name of Primer | Sequence Detail (5′→3′) |
|---|---|
| OPE-11 | GAGTCTCAGG |
| OPD-12 | CACCGTATCC |
| OPD-14 | CTTCCCCAAG |
| OPH-01 | GGTCGGAGAA |
| OPD-08 | GTGTGCCCCA |
| OPK-08 | GAACACTGGG |
| OPE-12 | TTATCGCCCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elyazid, D.M.A.; Salama, A.-M.; Zanaty, A.F.M.E.; Abdalla, N. In Vitro Propagation and Acclimatization of Banana Plants: Antioxidant Enzymes, Chemical Assessments and Genetic Stability of Regenerates as a Response to Copper Sulphate. Plants 2021, 10, 1853. https://doi.org/10.3390/plants10091853
Elyazid DMA, Salama A-M, Zanaty AFME, Abdalla N. In Vitro Propagation and Acclimatization of Banana Plants: Antioxidant Enzymes, Chemical Assessments and Genetic Stability of Regenerates as a Response to Copper Sulphate. Plants. 2021; 10(9):1853. https://doi.org/10.3390/plants10091853
Chicago/Turabian StyleElyazid, Doaa M. Abou, Abdel-Moety Salama, Abdel Fattah M. El Zanaty, and Neama Abdalla. 2021. "In Vitro Propagation and Acclimatization of Banana Plants: Antioxidant Enzymes, Chemical Assessments and Genetic Stability of Regenerates as a Response to Copper Sulphate" Plants 10, no. 9: 1853. https://doi.org/10.3390/plants10091853
APA StyleElyazid, D. M. A., Salama, A.-M., Zanaty, A. F. M. E., & Abdalla, N. (2021). In Vitro Propagation and Acclimatization of Banana Plants: Antioxidant Enzymes, Chemical Assessments and Genetic Stability of Regenerates as a Response to Copper Sulphate. Plants, 10(9), 1853. https://doi.org/10.3390/plants10091853