
Roots of Apium graveolens and Petroselinum crispum—Insight into Phenolic Status against Toxicity Level of Trace Elements

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
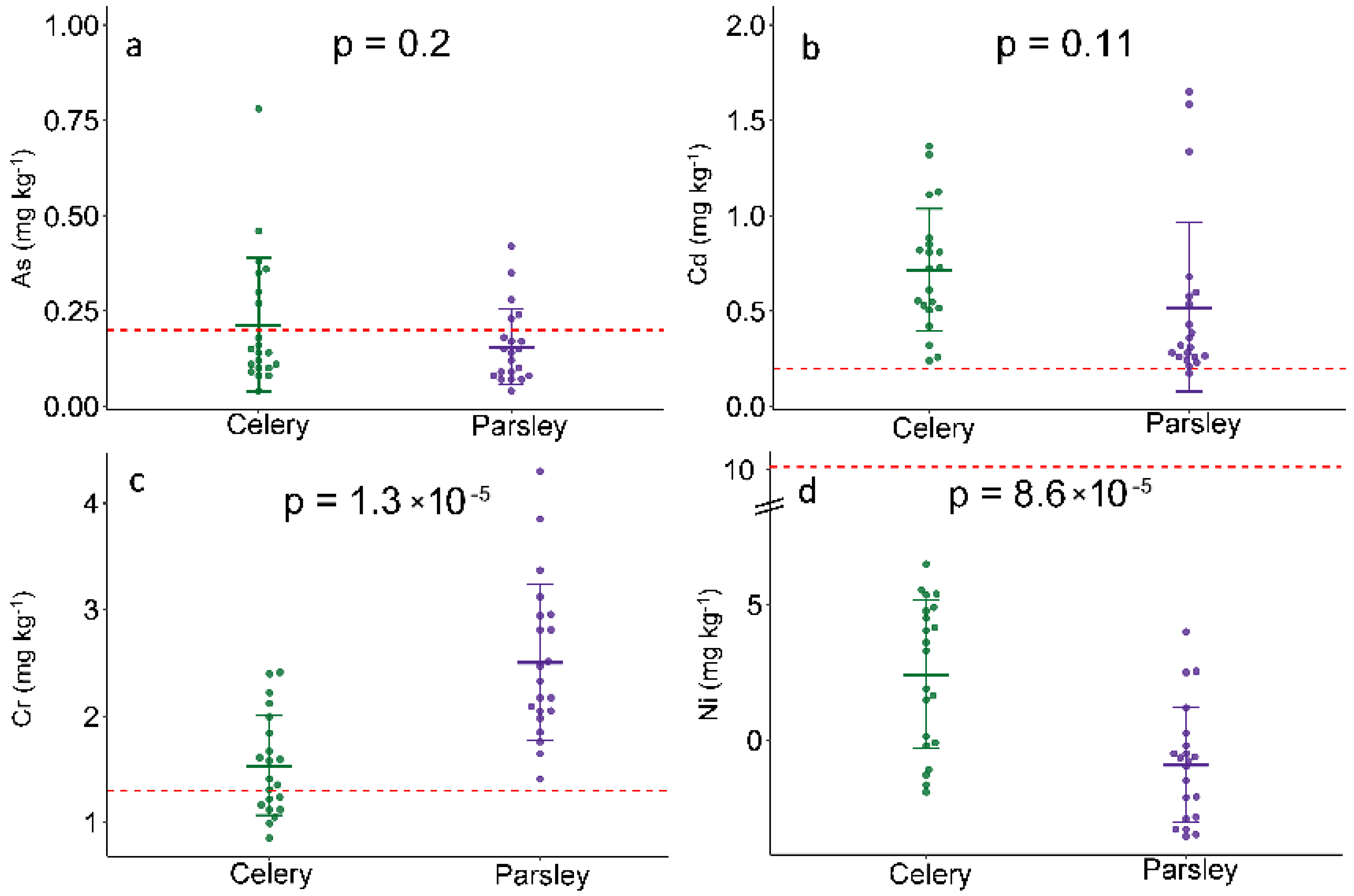
2.1. Trace Elements Accumulation
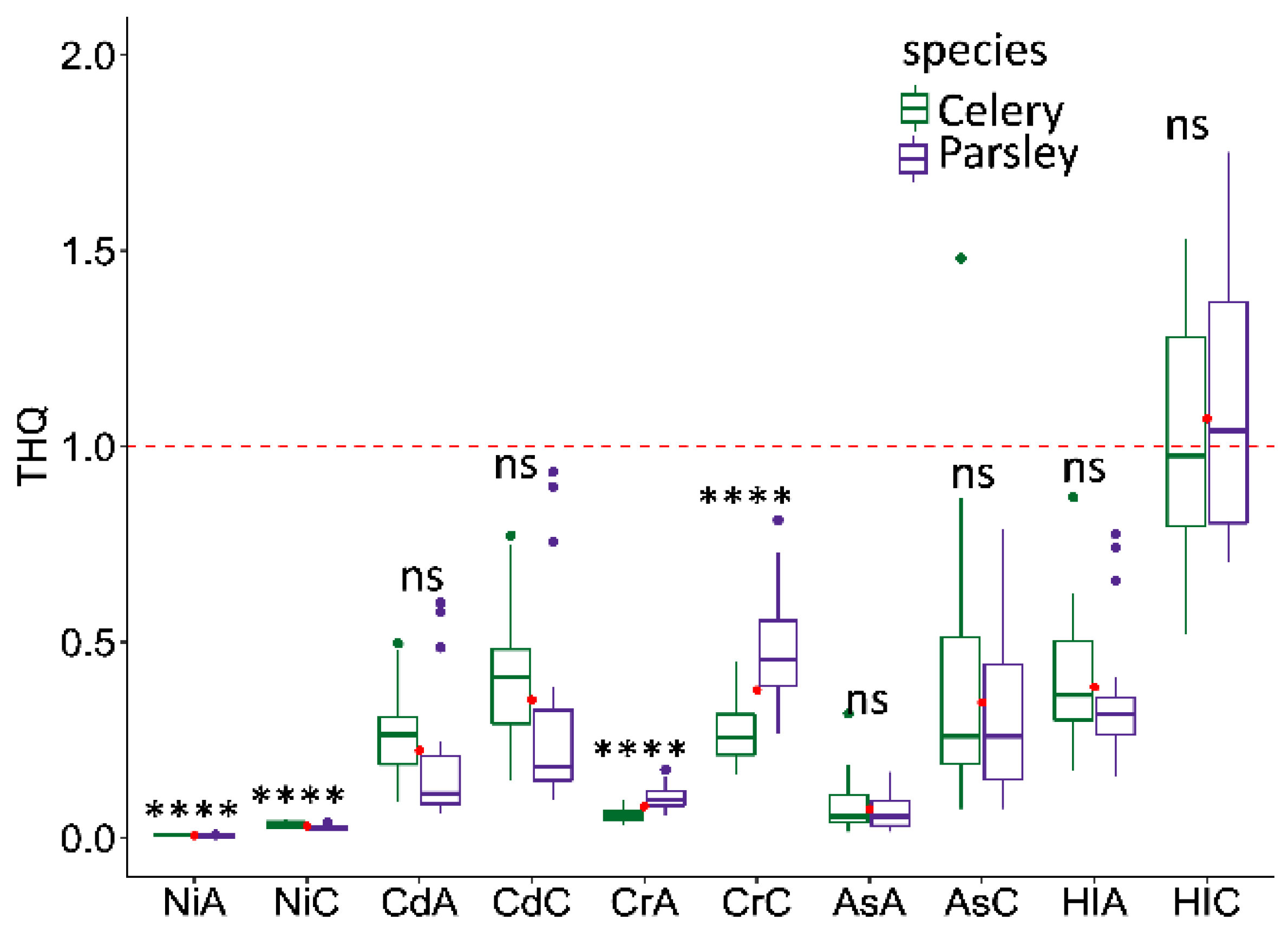
2.2. Health Risk Assessment
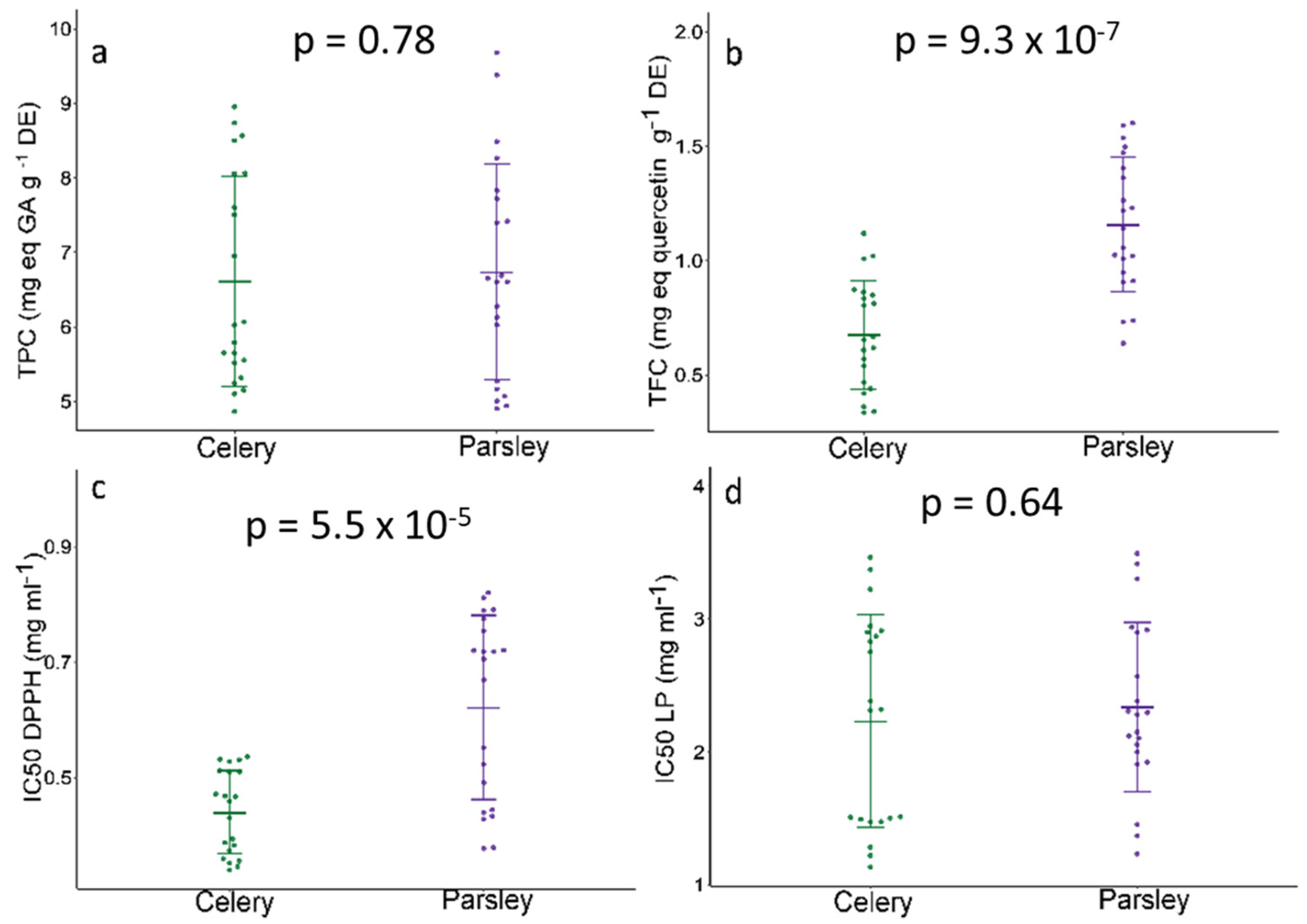
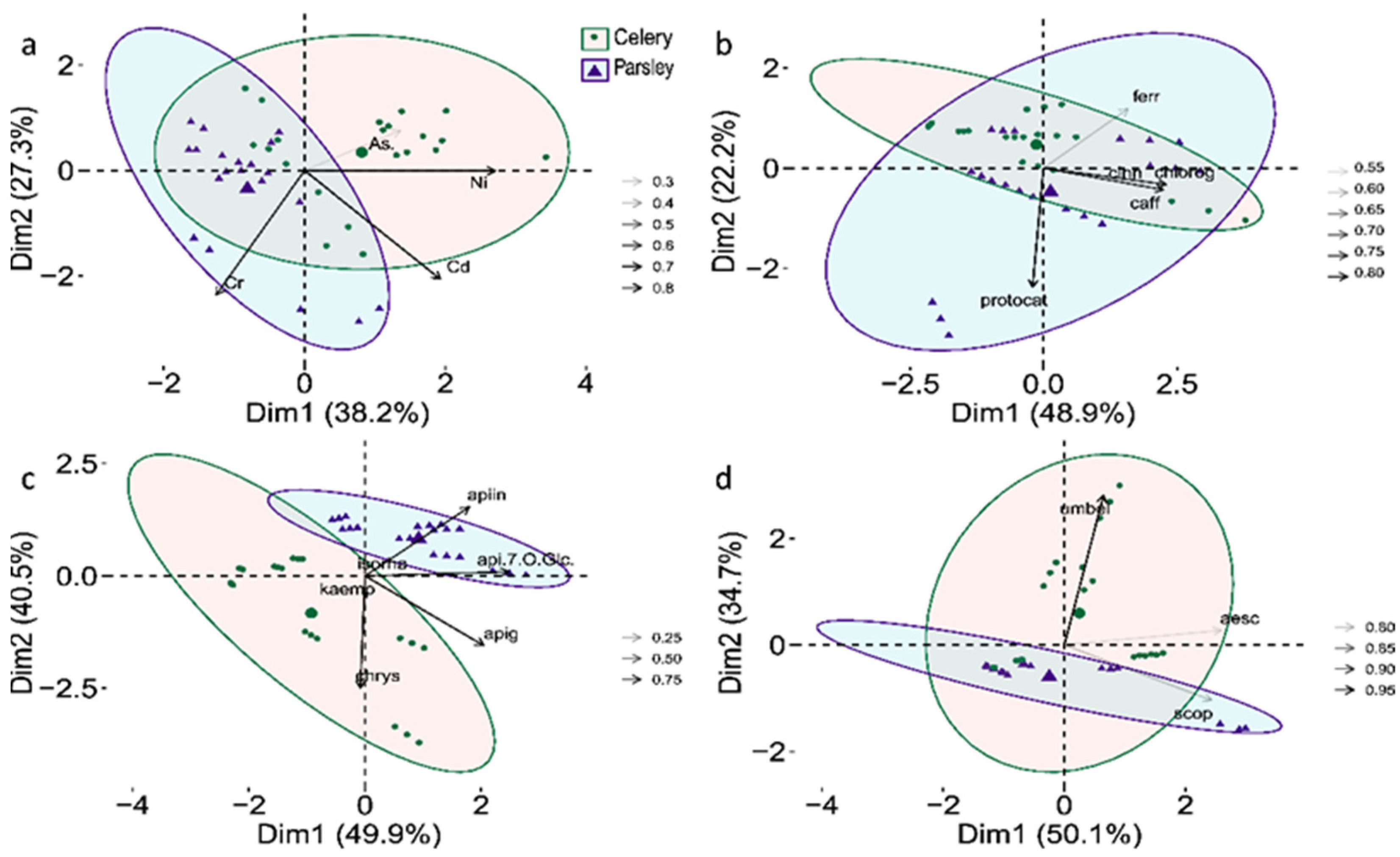
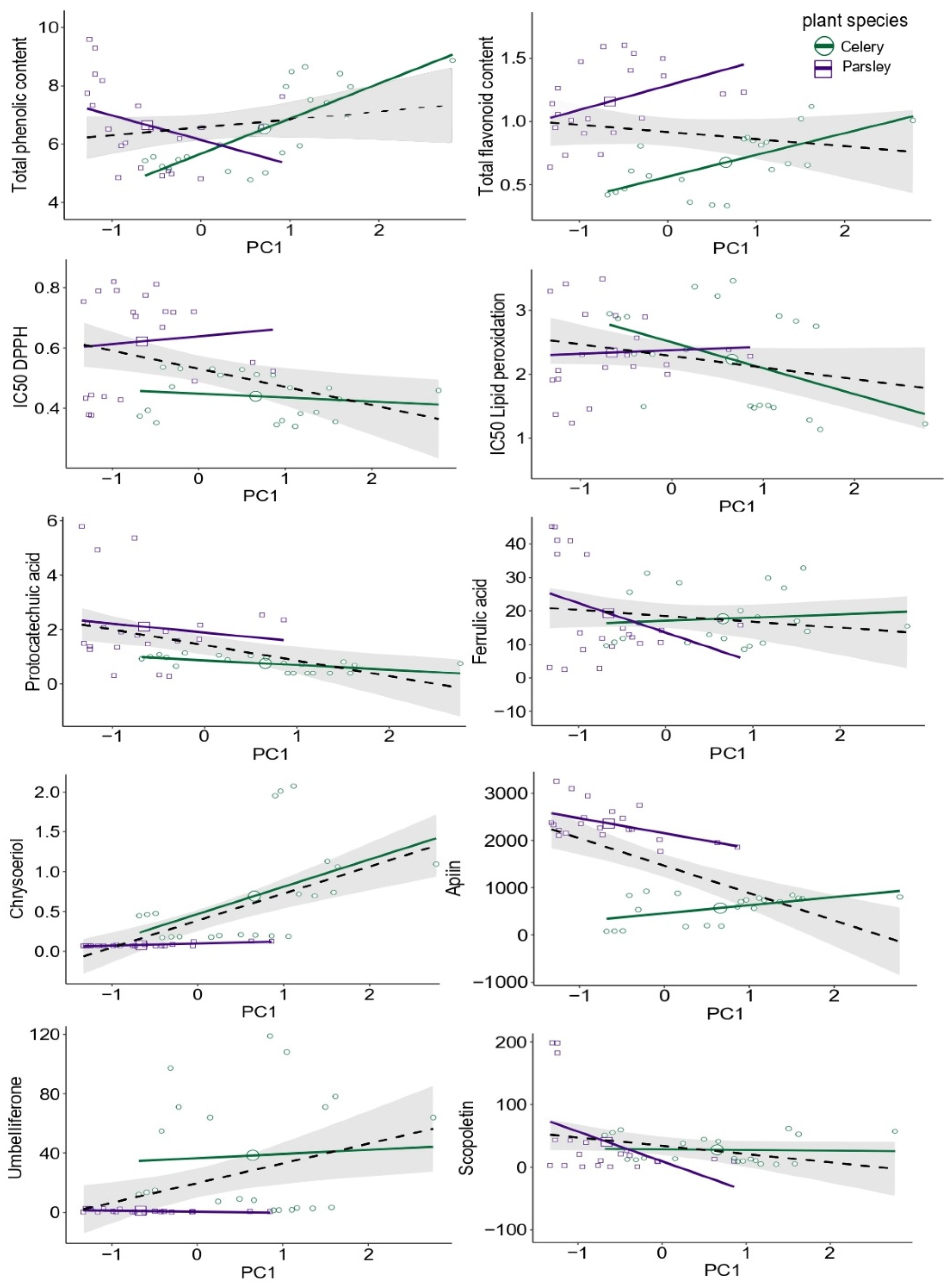
2.3. Antioxidative Potential and Phenolic Status—SMA Model Fitting
3. Materials and Methods
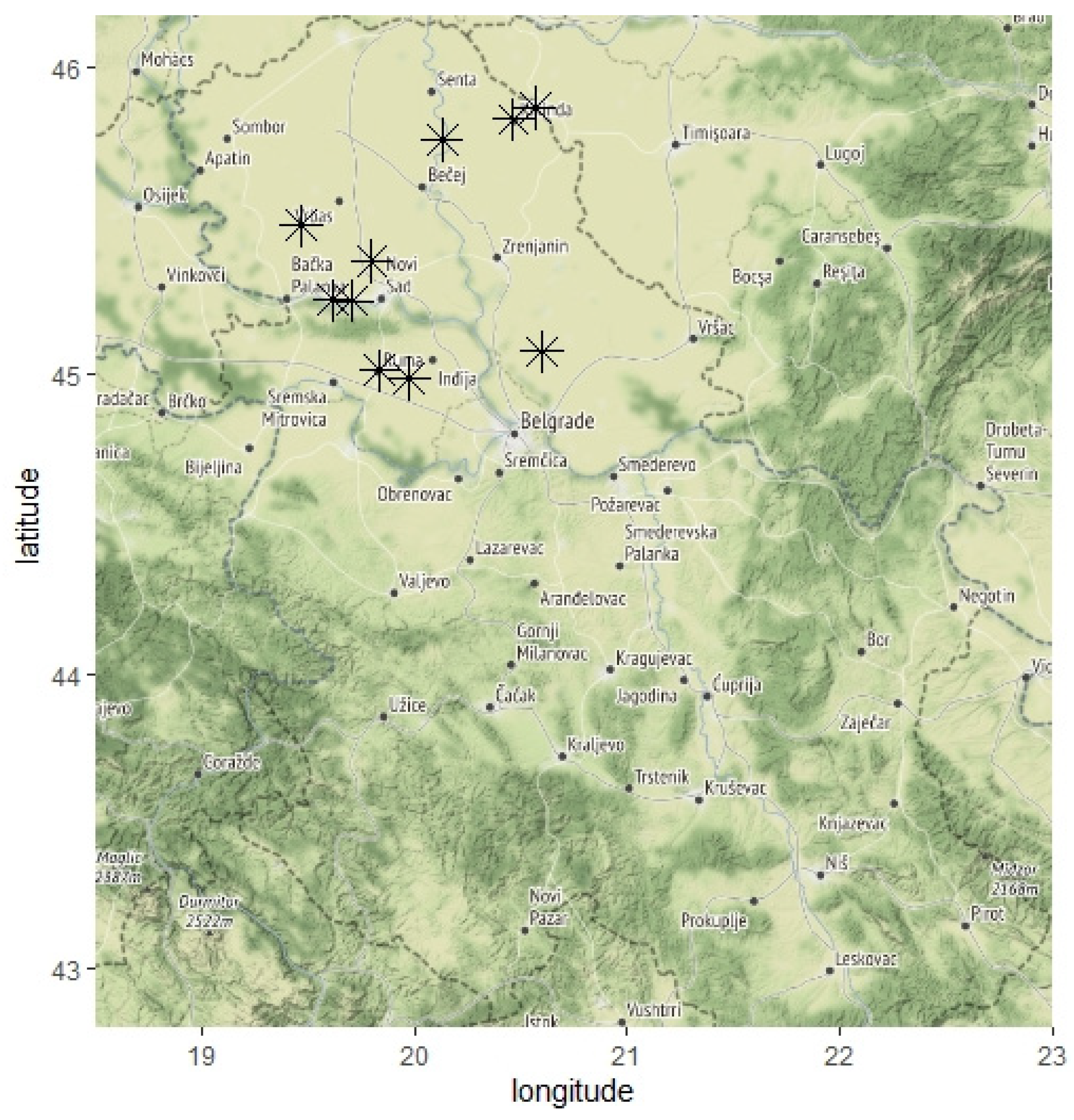
3.1. Vegetable Sampling
3.2. Trace Elements Analyses
3.3. Human Health Risk Assessment
3.4. Extract Preparation
3.5. Total Phenolic and Total Flavonoid Content
3.6. Quantitative LC-MS/MS Analysis of the Selected Phenolics
3.7. Antioxidant Activity
3.8. Data Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shankar, E.; Goel, A.; Gupta, K.; Gupta, S. Plant Flavone Apigenin: An Emerging Anticancer. Agent Curr. Pharmacol. Rep. 2017, 3, 423–446. [Google Scholar] [CrossRef] [PubMed]
- Cherng, J.M.; Chiang, W.; Chiang, L.C. Immunomodulatory activities of common vegetables and spices of Umbelliferae and its related coumarins and flavonoids. Food. Chem. 2008, 106, 944–950. [Google Scholar] [CrossRef]
- Pereira, M.P.; Tavano, O.L. Use of different spices as potential natural antioxidant additives on cooked beans (Phaseolus vulgaris). Increase of DPPH radical scavenging activity and total phenolic content. Plant. Foods Hum. Nutr. 2014, 69, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.M.; Selim, S.; Al, J.S.; Elgawad, H. CO2 enrichment can enhance the nutritional and health benefits of parsley (Petroselinum crispum L.) and dill (Anethum graveolens L.). Food Chem. 2018, 269, 519–526. [Google Scholar] [CrossRef]
- Farzaei, M.H.; Abbasabadi, Z.; Rahimi, R.; Farzaei, F. Parsley: A review of ethnopharmacology, phytochemistry and biological activities. J. Tradit. Chin. Med. 2013, 33, 815–826. [Google Scholar] [CrossRef] [Green Version]
- Golubkina, N.A.; Kharchenko, V.A.; Moldovan, A.I.; Koshevarov, A.A.; Zamana, S.; Nadezhkin, S.; Soldatenko, A.; Sekara, A.; Tallarita, A.; Caruso, G. Yield, Growth, Quality, Biochemical Characteristics and Elemental Composition of Plant Parts of Celery Leafy, Stalk and Root Types Grown in the Northern Hemisphere. Plants 2020, 9, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulusu, Y.; Ozturk, L.; Elmastas, M. Antioxidant capacity and cadmium accumulation in parsley seedlings exposed to cadmium stress. Russ. J. Plant Physiol. 2017, 64, 883–888. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Mikallou, M.; Petropoulos, S.; Tzortzakis, N. Profiling of Essential Oils Components and Polyphenols for Their Antioxidant Activity of Medicinal and Aromatic Plants Grown in Different Environmental Conditions. Agronomy 2020, 10, 727. [Google Scholar] [CrossRef]
- Popović, M.; Kaurinović, B.; Trivić, S.; Mimica-Dukić, N.; Bursać, M. Effect of celery (Apium graveolens) extracts on some biochemical parameters of oxidative stress in mice treated with carbon tetrachloride. Phytother. Res. 2007, 20, 531–537. [Google Scholar] [CrossRef]
- Bruznican, S.; De Clercq, H.; Eeckhaut, T.; Van Huylenbroeck, J.; Geelen, D. Celery and Celeriac: A critical view on present and future breeding. Front. Plant Sci. 2020, 10, 1699. [Google Scholar] [CrossRef] [PubMed]
- Kharchoufa, L.; Bouhrim, M.; Bencheikh, N.; Addi, M.; Hano, C.; Mechchate, H.; Elachouri, M. Potential Toxicity of Medicinal Plants Inventoried in Northeastern Morocco: An Ethnobotanical Approach. Plants 2021, 10, 1108. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [Green Version]
- Bibi, A.; Farooq, U.; Naz, S.; Khan, A.; Khan, K.; Sarwar, K.; Mahmood, Q.; Alam, A.; Mirza, N. Phytoextraction of Hg by parsley (Petroselinum crispum) and its growth responses. Int. J. Phytoremed. 2016, 18, 354–357. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, B.; Lu, Y.; Lu, F.; Wu, X.; You, W.; Huang, B. Bioavailability of cadmium to celery (Apium graveolens L.) grown in acidic and Cd-contaminated greenhouse soil as affected by the application of hydroxyapatite with different particle sizes. Chemosphere 2020, 240, 124916. [Google Scholar] [CrossRef]
- Roba, C.; Roşu, C.; Piştea, I.; Ozonu, A.; Baciu, C. Heavy metal content in vegetables and fruits cultivated in Baia Mare mining area (Romania) and health risk assessment. Environ. Sci. Pollut. Res. 2016, 23, 6062–6073. [Google Scholar] [CrossRef] [PubMed]
- Arsenov, D.; Župunski, M.; Pajević, S.; Borišev, M.; Nikolić, N.; Mimica-Dukić, N. Health assessment of medicinal herbs, celery and parsley related to cadmium soil pollution-potentially toxic elements (PTEs) accumulation, tolerance capacity and antioxidative response. Environ. Geochem. Health 2021, 43, 2927–2943. [Google Scholar] [CrossRef] [PubMed]
- Warton, D.I.; Wright, I.J.; Falster, D.S.; Westoby, M. Bivariate line-fitting methods for allometry. Biol. Rev. 2006, 81, 259–291. [Google Scholar] [CrossRef] [PubMed]
- De Temmerman, L.; Ruttens, A.; Waegeneers, N. Impact of atmospheric deposition of As, Cd and Pb on their concentration in carrot and celeriac. Environ. Pollut. 2012, 166, 187–195. [Google Scholar] [CrossRef]
- Ninkov, J.; Banjac, D.; Milić, S.; Vasin, J.; Marinković, J.; Banjac, B.; Mihailović, A. Arsenic content and distribution in agricultural soils of Vojvodina Province, Serbia. In Congress Proceedings 2nd International and 14th National Congress of Soil Science Society of Serbia Solutions and Projections for Sustainable Soil Management; Soil Science Society of Serbia: Novi Sad, Serbia, 2017; pp. 93–101. [Google Scholar]
- Spahić, M.P.; Sakan, S.; Cvetković, Ž.; Tančić, P.; Trifković, J.; Nikić, Z.; Manojlović, D. Assessment of contamination, environmental risk, and origin of heavy metals in soils surrounding industrial facilities in Vojvodina, Serbia. Environ. Monit. Assess. 2018, 190, 208. [Google Scholar] [CrossRef]
- Škrbić, B.D.; Živančev, J.; Antić, I.; Buljovčić, M. Pollution status and health risk caused by heavy elements in the flooded soil and vegetables from typical agricultural region in Vojvodina Province, Serbia. Environ. Sci. Pollut. Res. 2021, 28, 16065–16080. [Google Scholar] [CrossRef]
- Gupta, D.K.; Tripathi, R.D.; Mishra, S.; Srivastava, S.; Dwivedi, S.; Rai, U.N.; Yang, X.E.; Huanji, H.; Inouhe, M. Arsenic accumulation in root and shoot vis-a-vis its effects on growth and level of phytochelatins in seedlings of Cicer arietinum L. J. Environ. Biol. 2008, 29, 281–286. [Google Scholar]
- Monboonpitak, N.; Ruangwises, S.; Buranaphalin, S.; Ruangwises, N. Probabilistic risk assessment of inorganic arsenic via consumption of herbs collected in Thailand. Evid. Based Complement. Altern. Med. 2018, 2018, 8646579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontcuberta, M.; Calderon, J.; Villalbí, J.R.; Centrich, F.; Portana, S.; Espelt, A.; Duran, J.; Nebot, M. Total and inorganic arsenic in marketed food and associated health risks for the Catalan (Spain) Population. J. Agric. Food Chem. 2011, 59, 10013–10022. [Google Scholar] [CrossRef] [PubMed]
- Srinavas, N.; Ramakrishna, R.S.; Suresh, K.K. Trace metal accumulation in vegetables grown in industrial and semi-urban areas—A case study. Appl. Ecol. Environ. Res. 2009, 7, 131–139. [Google Scholar] [CrossRef]
- Dziubanek, G.; Baranowska, R.; Ćwieląg-Drabeka, M.; Spychała, A.; Piekut, A.; Rusina, M.; Hajok, I. Cadmium in edible plants from Silesia, Poland, and its implications for health risk in populations. Ecotoxicol. Environ. Saf. 2017, 142, 8–13. [Google Scholar] [CrossRef]
- Ćwieląg-Drabek, M.; Piekut, A.; Gut, K.; Grabowski, M. Risk of cadmium, lead and zinc exposure from consumption of vegetables produced in areas with mining and smelting past. Sci. Rep. 2020, 10, 3363. [Google Scholar] [CrossRef] [PubMed]
- Babula, P.; Klejdus, B.; Kovacik, J.; Hedbavny, J.; Hlavna, M. Lanthanum rather than cadmium induces oxidative stress and metabolite changes in Hypericum perforatum. J. Hazard. Mater. 2015, 286, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Qi, M.; Li, P.; Zhan, Y.; Shao, H. Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, B.; Young Chung, H.Y.; Kim, N.D. Role of Apigenin in Cancer Prevention via the Induction of Apoptosis and Autophagy. J. Cancer Prev. 2016, 21, 216–226. [Google Scholar] [CrossRef] [Green Version]
- Kolarovic, J.; Popovic, M.; Zlinská, J.; Trivic, S.; Vojnovic, M. Antioxidant Activities of Celery and Parsley Juices in Rats Treated with Doxorubicin. Molecules 2010, 15, 6193–6204. [Google Scholar] [CrossRef] [Green Version]
- Hameed, A.; Akram, N.A.; Saleem, M.H.; Ashraf, M.; Ahmed, S.; Ali, S.; Abdullah Alsahli, A.; Alyemeni, M.N. Seed Treatment with α-Tocopherol Regulates Growth and Key Physio-Biochemical Attributes in Carrot (Daucus carota L.) Plants under Water Limited Regimes. Agronomy 2021, 11, 469. [Google Scholar] [CrossRef]
- Chen, Y.G.; Huang, J.H.; Luo, R.; Ge, H.Z.; Wołowicz, A.; Wawrzkiewicz, M.; Gładysz-Płaska, A.; Li, B.; Yu, Q.X.; Kołodyńska, D.; et al. Impacts of heavy metals and medicinal crops on ecological systems, environmental pollution, cultivation, and production processes in China. Ecotoxicol. Environ. Saf. 2021, 219, 112336. [Google Scholar] [CrossRef] [PubMed]
- Tariq, M.; Saleem, M.H.; Aslam, S.; Rehman, M.; Iqbal, N.; Begum, R.; Ali, S.; Abdullah Alsahli, A.; Alyemeni, M.N.; Wijaya, L. Elucidating silicon-mediated distinct morpho-physio-biochemical attributes and organic acid exudation patterns of cadmium stressed Ajwain (Trachyspermum ammi L.). Plant Physiol. Biochem. 2020, 157, 23–37. [Google Scholar]
- Pajević, S.; Arsenov, D.; Nikolić, N.; Borišev, M.; Orčić, D.; Župunski, M.; Mimica-Dukić, N. Heavy metal accumulation in vegetable species and health risk assessment in Serbia. Environ. Monit. Assess. 2018, 190, 459. [Google Scholar] [CrossRef]
- Gebeyehu, H.R.; Bayissa, L.D. Levels of heavy metals in soil and vegetables and associated health risks in Mojo area, Ethiopia. PLoS ONE 2020, 15, e0227883. [Google Scholar]
- Harmanescu, M.; Alda, L.A.; Bordean, D.M.; Gogoasa, I.; Gerge, I. Heavy metals health risk assessment for population via consumption of vegetables grown in old mining area; a case study: Banat County, Romania. Chem. Cent. J. 2011, 5, 64. [Google Scholar] [CrossRef] [Green Version]
- U.S. EPA. Supplemental Guidance for Developing Soil Screening Levels for Superfund Sites; OSWER 9355.4-24; U.S. Environmental Protection Agency, Office of Emergency and Remedial Response: Washington, DC, USA, 2002.
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-ciocalteu reagent. Method Enzymol. 1999, 299, 152–178. [Google Scholar]
- Chang, C.C.; Yang, M.H.; Wen, H.M.; Chern, J.C. Estimation of total flavonoid content in propolis by two complementary colorimetric methods. J. Food Drug Anal. 2002, 10, 3. [Google Scholar]
- Beara, I.N.; Lesjak, M.M.; Orčić, D.Z.; Simin, N.Đ.; Četojević-Simin, D.D.; Božin, B.N.; Mimica-Dukić, N.M. Comparative analysis of phenolic profile, antioxidant, anti-inflammatory and cytotoxic activity of two closely-related Plantain species: Plantago altissima L. and Plantago lanceolata L. LWT-Food Sci. Technol. 2012, 47, 64–70. [Google Scholar] [CrossRef]
- Soler-Rivas, C.; Espín, J.C.; Wichers, H.J. An easy and fast test to compare total free radical scavenger capacity of foodstuffs. Phytochem. Anal. 2000, 11, 330–338. [Google Scholar] [CrossRef]
- Aust, S.D. Lipid peroxidation. In Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC Press: Boca Raton, FL, USA, 1985; pp. 203–207. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Celery | Parsley | MTDI | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Adults | Children | Adults | Children | ||||||
| Mean | SD | Mean | SD | Mean | SD | Mean | SD | ||
| Ni | 1.53 × 10−4 | 3.18 × 10−5 | 8.73 × 10−4 | 7.14 × 10−4 | 1.12 × 10−4 | 5.24 × 10−4 | 5.24 × 10−4 | 1.14 × 10−4 | 0.1–0.3 |
| Cd | 2.64 × 10−4 | 1.10 × 10−4 | 4.65 × 10−4 | 4.11 × 10−4 | 1.83 × 10−4 | 1.60 × 10−4 | 2.95 × 10−4 | 2.46 × 10−4 | 0.02–0.07 |
| Cr | 1.79 × 10−4 | 5.13 × 10−5 | 6.91 × 10−4 | 8.36 × 10−4 | 3.06 × 10−4 | 8.65 × 10−5 | 1.37 × 10−3 | 1.43 × 10−3 | 0.035–0.2 |
| As | 2.43 × 10−5 | 2.07 × 10−5 | 1.54 × 10−4 | 1.53 × 10−4 | 2.02 × 10−5 | 1.31 × 10−5 | 2.15 × 10−4 | 1.12 × 10−4 | 0.13 |
| Celery | Parsley | ANOVA | ||||
|---|---|---|---|---|---|---|
| Median | MAD | Median | MAD | p Values | Sign 1 | |
| p-OH-benzoic acid | 2.3 | 0 | LOD | 0 | 0.08 | . |
| cinnamic acid | 2.4 | 0.37 | 2.18 | 0.75 | 0.13 | ns |
| protocatechuic acid | 0.77 | 0.35 | 1.79 | 0.64 | 2.00 × 10−4 | *** |
| 2,5-dihydroxybenzoic acid | LOD | 0 | 2.68 | 0.56 | 4.30 × 10−5 | *** |
| p-coumaric acid | LOD | 0 | 1.55 | 0.66 | 1.10 × 10−5 | *** |
| caffeic acid | 2.46 | 0.55 | 2.84 | 0.76 | 0.08 | . |
| ferulic acid | 13.5 | 7.22 | 15.4 | 6.09 | 0.65 | ns |
| chlorogenic acid | 9.14 | 6.4 | 9.83 | 12.9 | 0.29 | ns |
| apigenin | 0.89 | 0.96 | 1.83 | 0.96 | 0.51 | ns |
| apigenin-7-O-glucoside | 0.8 | 0.69 | 2.32 | 0.87 | 3.80 × 10−5 | *** |
| chrysoeriol | 0.46 | 0.41 | 0.07 | 0 | 7.89 × 10−5 | *** |
| apiin | 705.41 | 207.23 | 2266.3 | 303.65 | <2.2× 10−16 | *** |
| umbelliferone | 13.5 | 17.8 | 0.56 | 0.53 | 1.25 × 10−4 | *** |
| aesculetin | 0.37 | 0.4 | 0.1 | 0 | 0.31 | * |
| scopoletin | 13.9 | 13 | 13.0 | 15.3 | 0.39 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arsenov, D.; Župunski, M.; Pajević, S.; Nemeš, I.; Simin, N.; Alnuqaydan, A.M.; Watson, M.; Aloliqi, A.A.; Mimica-Dukić, N. Roots of Apium graveolens and Petroselinum crispum—Insight into Phenolic Status against Toxicity Level of Trace Elements. Plants 2021, 10, 1785. https://doi.org/10.3390/plants10091785
Arsenov D, Župunski M, Pajević S, Nemeš I, Simin N, Alnuqaydan AM, Watson M, Aloliqi AA, Mimica-Dukić N. Roots of Apium graveolens and Petroselinum crispum—Insight into Phenolic Status against Toxicity Level of Trace Elements. Plants. 2021; 10(9):1785. https://doi.org/10.3390/plants10091785
Chicago/Turabian StyleArsenov, Danijela, Milan Župunski, Slobodanka Pajević, Ivana Nemeš, Nataša Simin, Abdullah M. Alnuqaydan, Malcolm Watson, Abdulaziz A. Aloliqi, and Neda Mimica-Dukić. 2021. "Roots of Apium graveolens and Petroselinum crispum—Insight into Phenolic Status against Toxicity Level of Trace Elements" Plants 10, no. 9: 1785. https://doi.org/10.3390/plants10091785
APA StyleArsenov, D., Župunski, M., Pajević, S., Nemeš, I., Simin, N., Alnuqaydan, A. M., Watson, M., Aloliqi, A. A., & Mimica-Dukić, N. (2021). Roots of Apium graveolens and Petroselinum crispum—Insight into Phenolic Status against Toxicity Level of Trace Elements. Plants, 10(9), 1785. https://doi.org/10.3390/plants10091785