
Advances in Genomics Approaches Shed Light on Crop Domestication

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Genomic Methods for Studying Crop Domestication
2.1. Population Genomics
2.2. Genome-Wide Association Studies (GWAS)
2.3. QTL Mapping
2.4. Genome Editing Using CRISPR-Cas Technology
3. Current Status of Research on Crop Domestication
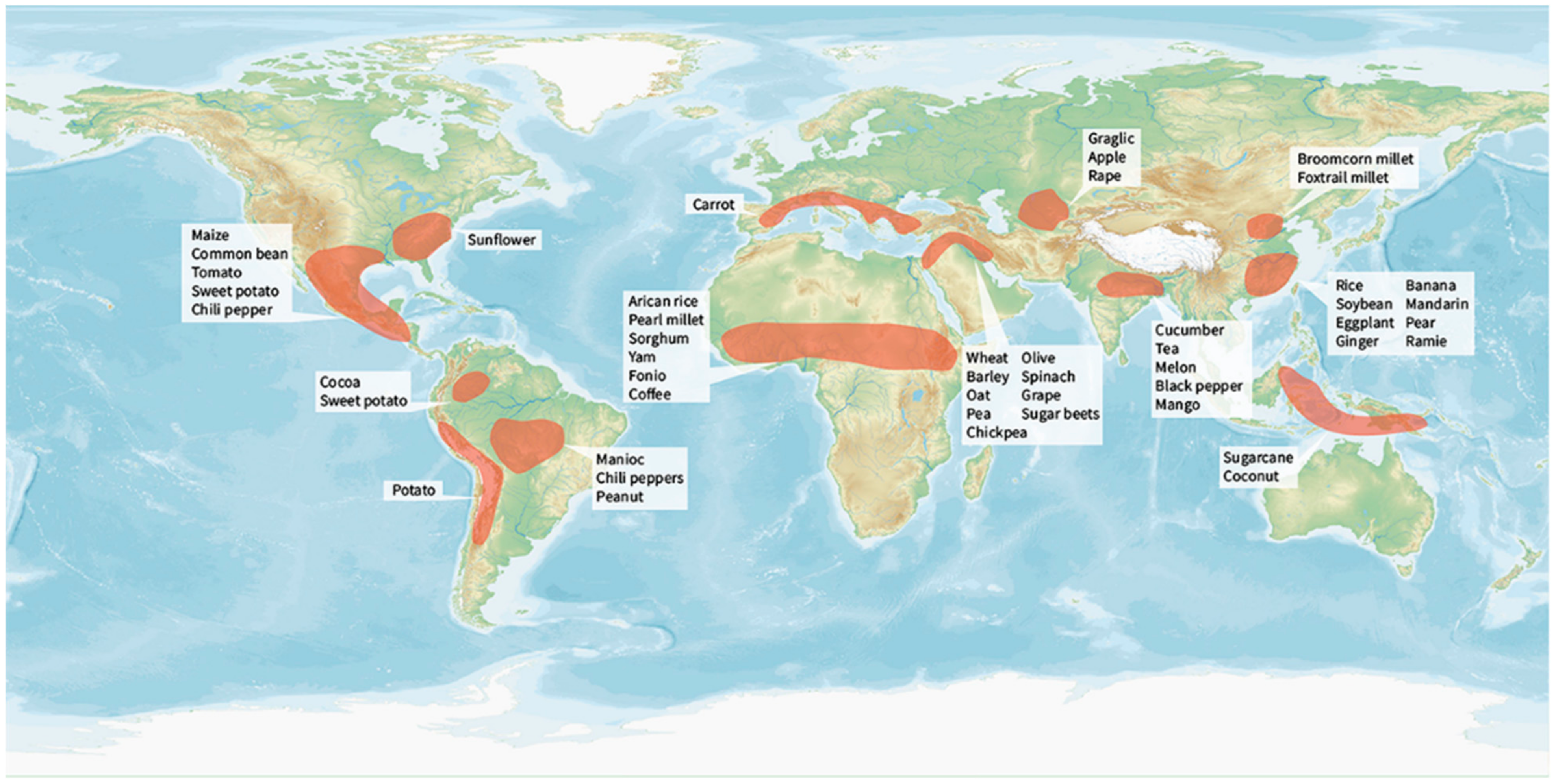
3.1. Domestication Centers and Their Spread
3.2. Domestication Theory
3.3. Genetic Architecture and the Molecular Basis of Genes Mediating Crop Domestication
4. Domestication in Modern Crop Breeding
4.1. Rice
4.2. Tomato
4.3. Potato
4.4. Orphan Crops
5. Crop Domestication Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Purugganan, M.D.; Fuller, D.Q. The nature of selection during plant domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef]
- Meyer, R.S.; Purugganan, M.D. Evolution of crop species: Genetics of domestication and diversification. Nat. Rev. Genet. 2013, 14, 840–852. [Google Scholar] [CrossRef]
- Ross-Ibarra, J.; Morrell, P.L.; Gaut, B.S. Plant domestication, a unique opportunity to identify the genetic basis of adaptation. Proc. Natl. Acad. Sci. USA 2007, 104, 8641–8648. [Google Scholar] [CrossRef] [PubMed]
- Smýkal, P.; Nelson, M.N.; Berger, J.D.; von Wettberg, E.J.B. The impact of genetic changes during crop domestication. Agronomy 2018, 8, 119. [Google Scholar] [CrossRef]
- Lobell, D.B.; Gourdji, S.M. The influence of climate change on global crop productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [PubMed]
- Doebley, J.; Stec, A.; Hubbard, L. The evolution of apical dominance in maize. Nature 1997, 386. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.; Li, X.; Liu, F.; Sun, X.; Li, C.; Zhu, Z.; Fu, Y.; Cai, H.; Wang, X.; Xie, D.; et al. Control of a key transition from prostrate to erect growth in rice domestication. Nat. Genet. 2008, 40, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Li, X.; Shannon, L.M.; Yeh, C.T.; Wang, M.L.; Bai, G.; Peng, Z.; Li, J.; Trick, H.N.; Clemente, T.E. Parallel domestication of the Shattering1 genes in cereals. Nat. Genet. 2012, 44, 720–724. [Google Scholar] [CrossRef]
- Doust, A.N.; Lukens, L.; Olsen, K.M.; Mauro-Herrera, M.; Meyer, A.; Rogers, K. Beyond the single gene: How epistasis and gene-by-environment effects influence crop domestication. Proc. Natl. Acad. Sci. USA 2014, 111, 6178–6183. [Google Scholar] [CrossRef]
- Pourkheirandish, M.; Hensel, G.; Kilian, B.; Senthil, N.; Chen, G.; Sameri, M.; Azhaguvel, P.; Sakuma, S.; Dhanagond, S.; Sharma, R.; et al. Evolution of the grain dispersal system in barley. Cell 2015, 162, 527–539. [Google Scholar] [CrossRef]
- Guerra-García, A.; Piñero, D. Current approaches and methods in plant domestication studies. Bot. Sci. 2017, 95, 345. [Google Scholar] [CrossRef][Green Version]
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.X.; Wang, A.; Zhao, Q.; Zhao, Y.; Liu, K.; Lu, H.; Li, W.; et al. A map of rice genome variation reveals the origin of cultivated rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef]
- Hufford, M.B.; Xu, X.; van Heerwaarden, J.; Pyhajarvi, T.; Chia, J.M.; Cartwright, R.A.; Elshire, R.J.; Glaubitz, J.C.; Guill, K.E.; Kaeppler, S.M.; et al. Comparative population genomics of maize domestication and improvement. Nat. Genet. 2012, 44, 808–811. [Google Scholar] [CrossRef]
- Schreiber, M.; Stein, N.; Mascher, M. Genomic approaches for studying crop evolution. Genome Biol. 2018, 19, 140. [Google Scholar] [CrossRef]
- Kistler, L.; Maezumi, S.Y.; de Souza, J.; Przelomska, N.A.S.; Malaquias Costa, F.; Smith, O.; Loiselle, H.; Ramos-Madrigal, J.; Wales, N.; Ribeiro, E.R.; et al. Multiproxy evidence highlights a complex evolutionary legacy of maize in South America. Science 2018, 362, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.F.; Botigue, L.R.; Brace, S.; Stevens, C.J.; Mullin, V.E.; Stevenson, A.; Thomas, M.G.; Fuller, D.Q.; Mott, R. A 3000-year-old Egyptian emmer wheat genome reveals dispersal and domestication history. Nat. Plants 2019, 5, 1120–1128. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tu, L.; Lin, M.; Lin, Z.; Wang, P.; Yang, Q.; Ye, Z.; Shen, C.; Li, J.; Zhang, L.; et al. Asymmetric subgenome selection and cis-regulatory divergence during cotton domestication. Nat. Genet. 2017, 49, 579–587. [Google Scholar] [CrossRef]
- Zhu, C.; Gore, M.; Buckler, E.S.; Yu, J. Status and prospects of association mapping in plants. Plant Genome 2008, 1, 5–20. [Google Scholar] [CrossRef]
- Guo, S.; Zhao, S.; Sun, H.; Wang, X.; Wu, S.; Lin, T.; Ren, Y.; Gao, L.; Deng, Y.; Zhang, J.; et al. Resequencing of 414 cultivated and wild watermelon accessions identifies selection for fruit quality traits. Nat. Genet. 2019, 51, 1616–1623. [Google Scholar] [CrossRef]
- Zhao, G.; Lian, Q.; Zhang, Z.; Fu, Q.; He, Y.; Ma, S.; Ruggieri, V.; Monforte, A.J.; Wang, P.; Julca, I.; et al. A comprehensive genome variation map of melon identifies multiple domestication events and loci influencing agronomic traits. Nat. Genet. 2019, 51, 1607–1615. [Google Scholar] [CrossRef]
- Mackay, T.F.C. The genetic architecture of quantitative traits. Annu. Rev. Genet. 2001. [Google Scholar] [CrossRef]
- Grisel, J.E.; Crabbe, J.C. Quantitative trait loci mapping. Alcohol. Health Res. World 1995, 19, 220–227. [Google Scholar]
- Konishi, S.; Izawa, T.; Lin, S.Y.; Ebana, K.; Yano, M. An SNP caused loss of seed shattering during rice domestication. Science 2006, 312, 1392–1396. [Google Scholar] [CrossRef]
- Li, C.; Zhou, A.; Sang, T. Rice domestication by reducing shattering. Science 2006, 1936–1939. [Google Scholar] [CrossRef]
- Zhang, L.-B.; Zhu, Q.; Wu, Z.-Q.; Ross-Ibarra, J.; Gaut, B.S.; Ge, S.; Sang, T. Selection on grain shattering genes and rates of rice domestication. New Phytol. 2009. [Google Scholar] [CrossRef]
- Gasiunas, G.; Barrangou, R.; Horvath, P.; Siksnys, V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, E2579–E2586. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 2012. [Google Scholar] [CrossRef] [PubMed]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169. [Google Scholar] [CrossRef] [PubMed]
- Makarova, K.S.; Haft, D.H.; Barrangou, R.; Brouns, S.J.J.; Charpentier, E.; Philippe Horvath, S.M.; Mojica, F.J.M.; Wolf, Y.I.; Yakunin, A.F.; van der Oost, J.; et al. Evolution and classification of the CRISPR–Cas systems. Nat. Rev. Microbiol. 2011, 9, 467–477. [Google Scholar] [CrossRef]
- Li, J.F.; Norville, J.E.; Aach, J.; Mccormack, M.; Zhang, D.; Bush, J.; Church, G.M.; Sheen, J. Multiplex and homologous recombination-mediated genome editing in arabidopsis and nicotiana benthamiana using guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.D.G.; Kamoun, S. Targeted mutagenesis in the model plant nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31. [Google Scholar] [CrossRef]
- Shan, Q.; Wang, Y.; Li, J.; Zhang, Y.; Chen, K.; Liang, Z.; Zhang, K.; Liu, J.; Xi, J.J.; Qiu, J.L. Targeted genome modification of crop plants using a CRISPR-Cas system. Nat. Biotechnol. 2013, 31, 686. [Google Scholar] [CrossRef]
- Haque, E.; Taniguchi, H.; Hassan, M.M.; Bhowmik, P.; Karim, M.R.; Smiech, M.; Zhao, K.; Rahman, M.; Islam, T. Application of CRISPR/Cas9 genome editing technology for the improvement of crops cultivated in tropical climates: Recent progress, prospects, and challenges. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Chen, K.; Wang, Y.; Zhang, R.; Zhang, H.; Gao, C. CRISPR/Cas genome editing and precision plant breeding in agriculture. Annu. Rev. Plant Biol. 2019, 70. [Google Scholar] [CrossRef]
- Schindele, A.; Dorn, A.; Puchta, H. CRISPR/Cas brings plant biology and breeding into the fast lane. Curr. Opin. Biotechnol. 2019, 61, 7–14. [Google Scholar] [CrossRef]
- Sedbrook, J.C.; Phippen, W.B.; Marks, M.D. New approaches to facilitate rapid domestication of a wild plant to an oilseed crop: Example pennycress (Thlaspi arvense L.). Plant Sci. 2014, 227, 122–132. [Google Scholar] [CrossRef]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.; Yu, Y.; Si, X.; Zhai, X.; Zhang, H.; Dong, W.; Gao, C.; Xu, C. Domestication of wild tomato is accelerated by genome editing. Nat. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Zsögön, A.; Cermak, T.; Naves, E.R.; Notini, M.M.; Edel, K.H.; Weinl, S.; Freschi, L.; Voytas, D.F.; Kudla, J.; Peres, L.E.P. De novo domestication of wild tomato using genome editing. Nat. Biotechnol. 2018. [Google Scholar] [CrossRef]
- Beying, N.; Schmidt, C.; Pacher, M.; Houben, A.; Puchta, H. CRISPR–Cas9-mediated induction of heritable chromosomal translocations in arabidopsis. Nat. Plants 2020, 1–8. [Google Scholar] [CrossRef]
- Fernie, A.R.; Yan, J. De Novo domestication: An alternative route toward new crops for the future. Mol. Plant 2019, 12, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Van Eck, J. Genome editing and plant transformation of solanaceous food crops. Curr. Opin. Biotechnol. 2018, 49. [Google Scholar] [CrossRef]
- Yamamoto, T.; Kashojiya, S.; Kamimura, S.; Kameyama, T.; Ariizumi, T.; Ezura, H.; Miura, K. Application and development of genome editing technologies to the solanaceae plants. Plant Physiol. Biochem. 2018, S0981942818300718. [Google Scholar] [CrossRef]
- Zeng, Y.; Wen, J.; Zhao, W.; Wang, Q.; Huang, W. Rational improvement of rice yield and cold tolerance by editing the three genes OsPIN5b, GS3, and OsMYB30 with the CRISPR–Cas9 system. Front. Plant Sci. 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, Z.; Zong, Y.; Wang, Y.; Liu, J.; Chen, K.; Qiu, J.L.; Gao, C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA. Nat. Commun. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Hu, B.; Li, D.; Liu, X.; Qi, J.; Gao, D.; Zhao, S.; Huang, S.; Sun, J.; Yang, L. Engineering Non-transgenic gynoecious cucumber using an improved transformation protocol and optimized CRISPR/Cas9 system. Mol. Plant 2017, 10, 1575–1578. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.; Piperno, D.R.; Allaby, R.G.; Purugganan, M.D.; Andersson, L.; Arroyo-Kalin, M.; Barton, L.; Climer Vigueira, C.; Denham, T.; Dobney, K.; et al. Current perspectives and the future of domestication studies. Proc. Natl. Acad. Sci. USA 2014, 111, 6139–6146. [Google Scholar] [CrossRef]
- Piperno, D.R.; Ranere, A.J.; Holst, I.; Iriarte, J.; Dickau, R. Starch grain and phytolith evidence for early ninth millennium B.P. maize from the Central Balsas River Valley, Mexico. Proc. Natl. Acad. Sci. USA 2009, 106, 5019–5024. [Google Scholar] [CrossRef] [PubMed]
- Van Heerwaarden, J.; Doebley, J.; Briggs, W.H.; Glaubitz, J.C.; Goodman, M.M.; Gonzalez, J.D.J.S.; Ross-Ibarra, J. Genetic signals of origin, spread, and introgression in a large sample of maize landraces. Proc. Natl. Acad. Sci. USA 2011, 108, 1088–1092. [Google Scholar] [CrossRef] [PubMed]
- Gepts, P. The contribution of genetic and genomic approaches to plant domestication studies. Curr. Opin. Plant Biol. 2014, 18, 51–59. [Google Scholar] [CrossRef]
- Olsen, K.M.; Wendel, J.F. A bountiful harvest: Genomic insights into crop domestication phenotypes. Annu. Rev. Plant Biol. 2013, 64, 47–70. [Google Scholar] [CrossRef] [PubMed]
- Doust, A.N.; Devos, K.M.; Gadberry, M.D.; Gale, M.D.; Kellogg, E.A. Genetic control of branching in foxtail millet. Proc. Natl. Acad. Sci. USA 2004, 101, 9045–9050. [Google Scholar] [CrossRef] [PubMed]
- Lai, X.; Yan, L.; Lu, Y.; Schnable, J.C. Largely unlinked gene sets targeted by selection for domestication syndrome phenotypes in maize and sorghum. Plant J. 2018, 93, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Sang, T.; Ge, S. Understanding rice domestication and implications for cultivar improvement. Curr. Opin. Plant Biol. 2013, 16, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Sang, T.; Ge, S. Genetics and phylogenetics of rice domestication. Curr. Opin. Genet. Dev. 2007, 17, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, H.; Zhu, H. CRISPR technology is revolutionizing the improvement of tomato and other fruit crops. Hortic. Res. 2019, 6, 1–13. [Google Scholar] [CrossRef]
- Hua, K.; Zhang, J.; Botella, J.R.; Ma, C.; Kong, F.; Liu, B.; Zhu, J.K. Perspectives on the Application of Genome-Editing Technologies in Crop Breeding. Mol. Plant 2019, 12, 1047–1059. [Google Scholar] [CrossRef]
- Sheng, X.; Sun, Z.; Wang, X.; Tan, Y.; Yu, D.; Yuan, G.; Yuan, D.; Duan, M. Improvement of the Rice “Easy-to-Shatter” Trait via CRISPR/Cas9-Mediated Mutagenesis of the qSH1 Gene. Front. Plant Sci. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Jung, Y.J.; Lee, H.J.; Bae, S.; Kim, J.H.; Kim, D.H.; Kim, H.K.; Nam, K.H.; Nogoy, F.M.; Cho, Y.G.; Kang, K.K. Acquisition of seed dormancy breaking in rice (Oryza sativa L.) via CRISPR/Cas9-targeted mutagenesis of OsVP1 gene. Plant Biotechnol. Rep. 2019, 13, 511–520. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, R.; Huang, G.; Li, Y.; Melaku, G.; Zhang, S.; Chen, H.; Zhao, Y.; Zhang, J.; Zhang, Y.; et al. Developing superior alleles of yield genes in rice by artificial mutagenesis using the CRISPR/Cas9 system. Crop. J. 2018, 6, 475–481. [Google Scholar] [CrossRef]
- Swinnen, G.; Goossens, A.; Pauwels, L. Lessons from Domestication: Targeting Cis-Regulatory Elements for Crop Improvement. Trends Plant Sci. 2016, 21, 506–515. [Google Scholar] [CrossRef]
- Yu, H.; Lin, T.; Meng, X.; Du, H.; Zhang, J.; Liu, G.; Chen, M.; Jing, Y.; Kou, L.; Li, X.; et al. A route to de novo domestication of wild allotetraploid rice. Cell 2021, 184, 1156–1170. [Google Scholar] [CrossRef]
- Sims, W.L. History of tomato production for industry around the world. Acta Hortic. 1980, 25–26. [Google Scholar] [CrossRef]
- Rick, C.M.; Chetelat, R.T. Utilization of related wild species for tomato improvement. Acta Hortic. 1995, 412, 21–38. [Google Scholar] [CrossRef]
- Larry, R.; Joanne, L. Genetic resources of tomato. In Genetic Improvement of Solanaceous Crops; Razdan, M.K., Mattoo, A.K., Eds.; Science Publishers: Enfield, NH, USA, 2007. [Google Scholar]
- Miller, J.C.; Tanksley, S.D. RFLP analysis of phylogenetic relationships and genetic variation in the genus Lycopersicon. Theor. Appl. Genet. 1990, 80, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Doganlar, S.; Frary, A.; Tanksley, S.D. The genetic basis of seed-weight variation: Tomato as a model system. Theor. Appl. Genet. 2000, 100, 1267–1273. [Google Scholar] [CrossRef]
- Van Der Knaap, E.; Lippman, Z.B.; Tanksley, S.D. Extremely elongated tomato fruit controlled by four quantitative trait loci with epistatic interactions. Theor. Appl. Genet. 2002, 104, 241–247. [Google Scholar] [CrossRef]
- Tanksley, S.D. The genetic, developmental, and molecular bases of fruit size and shape variation in tomato. Plant Cell 2004, 16, S181–S189. [Google Scholar] [CrossRef] [PubMed]
- Pnueli, L.; Carmel-Goren, L.; Hareven, D.; Gutfinger, T.; Alvarez, J.; Ganal, M.; Zamir, D.; Lifschitz, E. The SELF-PRUNING gene of tomato regulates vegetative to reproductive switching of sympodial meristems and is the ortholog of CEN and TFL1. Development 1998, 125, 1979–1989. [Google Scholar] [CrossRef]
- Lippman, Z.B.; Cohen, O.; Alvarez, J.P.; Abu-Abied, M.; Pekker, I.; Paran, I.; Eshed, Y.; Zamir, D. The making of a compound inflorescence in tomato and related nightshades. PLoS Biol. 2008, 6, e288. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Van Eck, J.; Cong, B.; Tanksley, S.D. A new class of regulatory genes underlying the cause of pear-shaped tomato fruit. Proc. Natl. Acad. Sci. USA 2002, 99. [Google Scholar] [CrossRef] [PubMed]
- Frary, A.; Nesbitt, T.C.; Grandillo, S.; Knaap, E.; Cong, B.; Liu, J.; Meller, J.; Elber, R.; Alpert, K.B.; Tanksley, S.D. fw2.2: A quantitative trait locus key to the evolution of tomato fruit size. Science 2000, 289, 85–88. [Google Scholar] [CrossRef]
- Xu, C.; Liberatore, K.L.; Macalister, C.A.; Huang, Z.; Chu, Y.H.; Jiang, K.; Brooks, C.; Ogawa-Ohnishi, M.; Xiong, G.; Pauly, M. A cascade of arabinosyltransferases controls shoot meristem size in tomato. Nat. Genet. 2015, 47, 784–792. [Google Scholar] [CrossRef]
- Ronen, G.; Carmel-Goren, L.; Zamir, D.; Hirschberg, J. An alternative pathway to β-carotene formation in plant chromoplasts discovered by map-based cloning of Beta and old-gold color mutations in tomato. Proc. Natl. Acad. Sci. USA 2000, 97, 11102–11107. [Google Scholar] [CrossRef]
- Spooner, D.M.; Ghislain, M.; Simon, R.; Jansky, S.H.; Gavrilenko, T. Systematics, diversity, genetics, and evolution of wild and cultivated potatoes. Bot. Rev. 2014, 80, 283–383. [Google Scholar] [CrossRef]
- Jansky, S.H.; Charkowski, A.O.; Douches, D.S.; Gusmini, G.; Richael, C.; Bethke, P.C.; Spooner, D.M.; Novy, R.G.; De Jong, H.; De Jong, W.S. Reinventing potato as a diploid inbred line–based crop. Crop. Sci. 2016, 56, 1412. [Google Scholar] [CrossRef]
- Enciso-Rodriguez, F.; Manrique-Carpintero, N.; Nadakuduti, S.S.; Buell, C.R.; Zarka, D.; Douches, D. Overcoming self-incompatibility in diploid potato using CRISPR-Cas9. Front. Plant Sci. 2019, 10, 376. [Google Scholar] [CrossRef]
- Ye, M.; Peng, Z.; Tang, D.; Yang, Z.; Li, D.; Xu, Y.; Zhang, C.; Huang, S. Generation of self-compatible diploid potato by knockout of S-RNase. Nat. Plants 2018, 4, 651–654. [Google Scholar] [CrossRef]
- Bohra, A.; Jha, U.C.; Kishor, P.B.; Pandey, S.; Singh, N.P. Genomics and molecular breeding in lesser explored pulse crops: Current trends and future opportunities. Biotechnol. Adv. 2014, 32, 1410–1428. [Google Scholar] [CrossRef] [PubMed]
- Tadele, Z.; Bartels, D. Promoting orphan crops research and development. Planta 2019, 250. [Google Scholar] [CrossRef] [PubMed]
- Särkinen, T.; Bohs, L.; Olmstead, R.; Knapp, S. A phylogenetic framework for evolutionary study of the nightshades (solanaceae): A dated 1000-tip tree. BMC Evol. Biol. 2013, 13, 1–15. [Google Scholar] [CrossRef]
- Harlan, J.R.; Yeatman, C.W.; Kafton, D.; Wilkes, E.G. Plant Genetic Resources: A Conservation Imperative; Westview: Boulder, CO, USA, 1987. [Google Scholar]
- Doebley, J.F.; Gaut, B.S.; Smith, B.D. The molecular genetics of crop domestication. Cell 2006, 127, 1309–1321. [Google Scholar] [CrossRef]
- Van Tassel, D.L.; Tesdell, O.; Schlautman, B.; Rubin, M.J.; DeHaan, L.R.; Crews, T.E.; Streit Krug, A. New Food Crop Domestication in the Age of Gene Editing: Genetic, Agronomic and Cultural Change Remain Co-evolutionarily Entangled. Front. Plant Sci. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tanksley, S.D.; McCouch, S.R. Seed banks and molecular maps: Unlocking genetic potential from the wild. Science 1997, 277, 1063–1066. [Google Scholar] [CrossRef] [PubMed]
- Vavilov, N.I. The new systematic of cultivated plants. In The New Systematic; Huxley, J., Ed.; Oxford University Press: London, UK, 1940; pp. 549–556. [Google Scholar]
- Peleman, J.D.; Van Der Voort, J.R. Breeding by design. Trends Plant Sci. 2003, 8, 330–334. [Google Scholar] [CrossRef]
- Islam, T. CRISPR-Cas technology in modifying food crops. CAB Rev. 2019, 14, 1–16. [Google Scholar] [CrossRef]
- Islam, M.T.; Bhowmik, P.K.; Molla, K.A. CRISPR-Cas Methods; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar] [CrossRef]


{kind=link}
| Crop | Population Type | Population Size | Key Statistic | Discovery | Ref. |
|---|---|---|---|---|---|
| Rice | Ancestral progenitor; cultivated indica and japonica varieties | 1529 | Sequence diversity (π) population-differentiation (FST), cross-population extended haplotype homozygosity (XP-EHH) | Identify 55 domestication sweeps, and reveal the domesticaiton and development of cultivated rice | [12] |
| Maize | Wild, landraces and improved maize lines | 75 | π, ρ, FST, Tajima’s D, normalized Fay and Wu’s H, and a composite likelihood approach (XP-CLR) | Evidence of recovery of diversity after domestication, and stronger selection for domestication than improvement | [13] |
| Maize | Ancient samples, modern maizes landraces, and teosintes | 134 | Mutation load, D-statistics, and f3 and f4 statistic | Reveal domestication center and human-mediated spread of maize | [15] |
| Wheat | Ancient and modern domestic emmer | 64 | Haplotype structure | Uncover the history and diversity of emmar wheat | [16] |
| Cotton | Wild and domesticated cotton accessions | 352 | π, FST, and XP-CLR | Identify 93 domestication sweeps | [17] |
| Crop | Domestication Trait | Population Type | Population Size | Genotype Method | Model | Discovery | Ref. |
|---|---|---|---|---|---|---|---|
| Rice | Leaf sheath color and tiller angle | Ancestral progenitor Oryza rufipogon | 446 | Whole genome resequencing | Compressed mixed linear model | Identify assoicaitons for OSC1 and PROG1 | [12] |
| Cotton | Fiber quality related traits | Cotton accessions | 267 | Whole genome resequencing | Compressed mixed linear model | Identify 19 assoicaiton signals (16 were new) | [17] |
| Watermelon | Fruit quality traits | Cultivated and wild watermelon accessions | 414 | Whole genome resequencing | Linear mixed model algorithm | Identify 43 associaiton signals (35 were new) | [19] |
| Crop | Domestication Trait | Population Type | Population Size | Marker | Discovery | Refs. |
|---|---|---|---|---|---|---|
| Rice | Seed shattering | F2 | 304 | RFLP, RAPD, SNP, SSR | Localized the gene qSH1 and gene sh4 | [23,24] |
| Barley | Rachis non-brittle | F2 | >10,000 | SNP | Localized the gene btr1 and btr2 | [10] |
| Foxtail millet | Shattering and flowering time | Recombinantinbred line | 182 | SNP, SSR, and sequence-tagged site markers | Two significant QTLs | [9] |
| Crop | Domestication Traits | Target Gene | Method | Discovery | Ref. |
|---|---|---|---|---|---|
| Rice | Panicle length, grain size, cold tolerance | OsPIN5b, GS3, OsMYB30 | CRISPR-Cas9 system edits three genes simultaneously | Higher yield and better cold tolerance in gene-edited rice | [44] |
| Wheat | Grein length, weight and yield, TKW, Inflorescence architecture, branching and tillering | Tagasr7-A1 (-B1 and –D1), TaDEP1, TaNAC2, TaPIN 1, and TaLOX2 | Transient expression of CRISPR-Cas9 in callus cells | Changes on target traits in wheat callus and regeneration of plants | [45] |
| Tomato | Fruit size, number and nutrition | SP, O, FW2.2, CycB | CRISPR-Cas9 system edits six genes simultaneously | Gene-edited tomato has at least a threefold increase in target traits | [39] |
| Cucumber | Carpel development | CsWip1 | Optimized CRISPR/Cas9 system with CsU6 promoter and GFP | Seven times more female flowers in gene-edited cumcumber | [46] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Feng, M.; Paudel, D.; Islam, T.; Momotaz, A.; Luo, Z.; Zhao, Z.; Wei, N.; Li, S.; Xia, Q.; et al. Advances in Genomics Approaches Shed Light on Crop Domestication. Plants 2021, 10, 1571. https://doi.org/10.3390/plants10081571
Zhao Y, Feng M, Paudel D, Islam T, Momotaz A, Luo Z, Zhao Z, Wei N, Li S, Xia Q, et al. Advances in Genomics Approaches Shed Light on Crop Domestication. Plants. 2021; 10(8):1571. https://doi.org/10.3390/plants10081571
Chicago/Turabian StyleZhao, Yang, Mengfan Feng, Dev Paudel, Tofazzal Islam, Aliya Momotaz, Ziliang Luo, Zifan Zhao, Ni Wei, Sicheng Li, Qing Xia, and et al. 2021. "Advances in Genomics Approaches Shed Light on Crop Domestication" Plants 10, no. 8: 1571. https://doi.org/10.3390/plants10081571
APA StyleZhao, Y., Feng, M., Paudel, D., Islam, T., Momotaz, A., Luo, Z., Zhao, Z., Wei, N., Li, S., Xia, Q., Kuang, B., Yang, X., & Wang, J. (2021). Advances in Genomics Approaches Shed Light on Crop Domestication. Plants, 10(8), 1571. https://doi.org/10.3390/plants10081571