
Mercury-Induced Phytotoxicity and Responses in Upland Cotton (Gossypium hirsutum L.) Seedlings

 , ,
, , 
Abstract
1. Introduction
2. Results
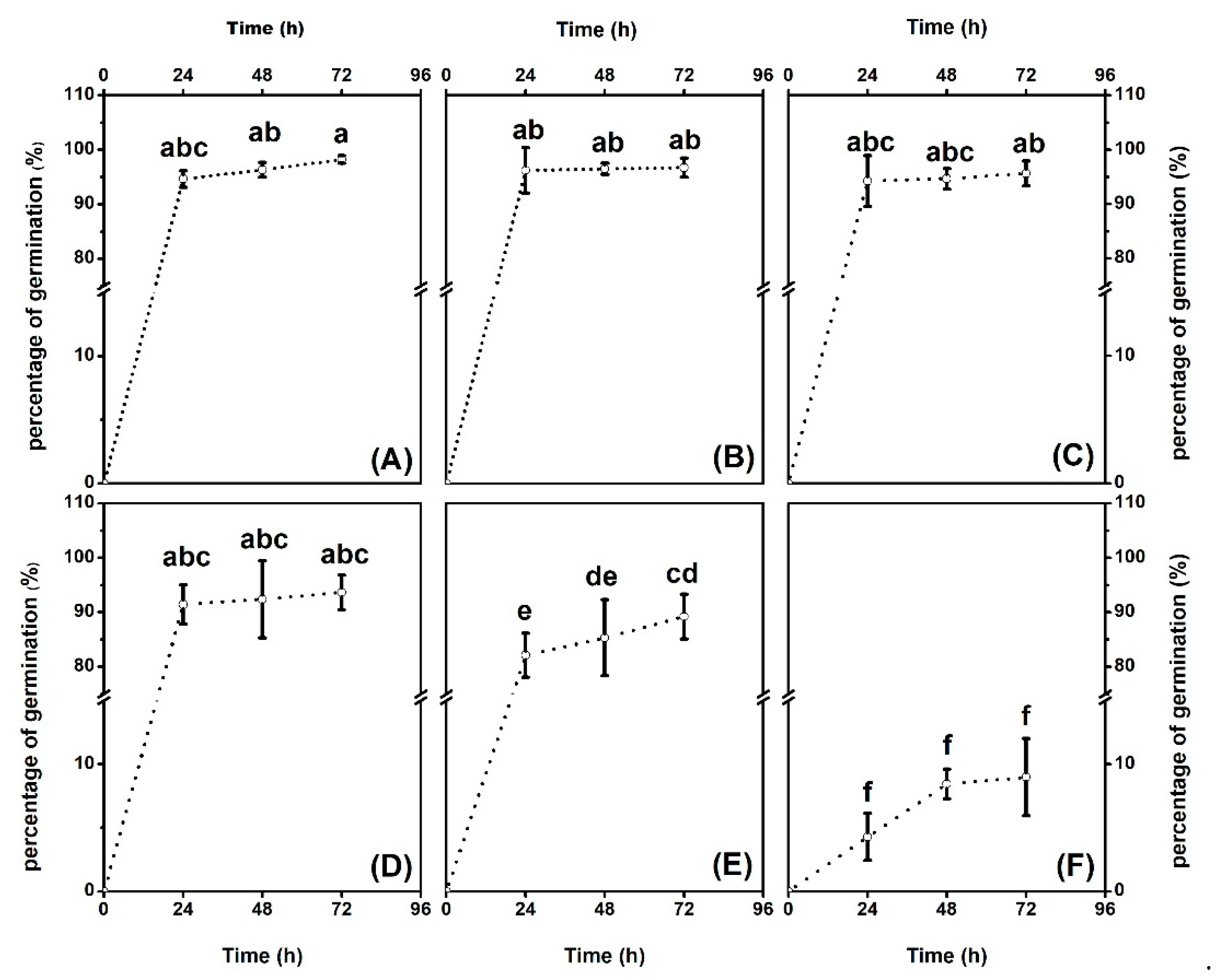
2.1. Effects of Hg on Cotton Seeds Germination
2.2. Growth and Hg Distribution
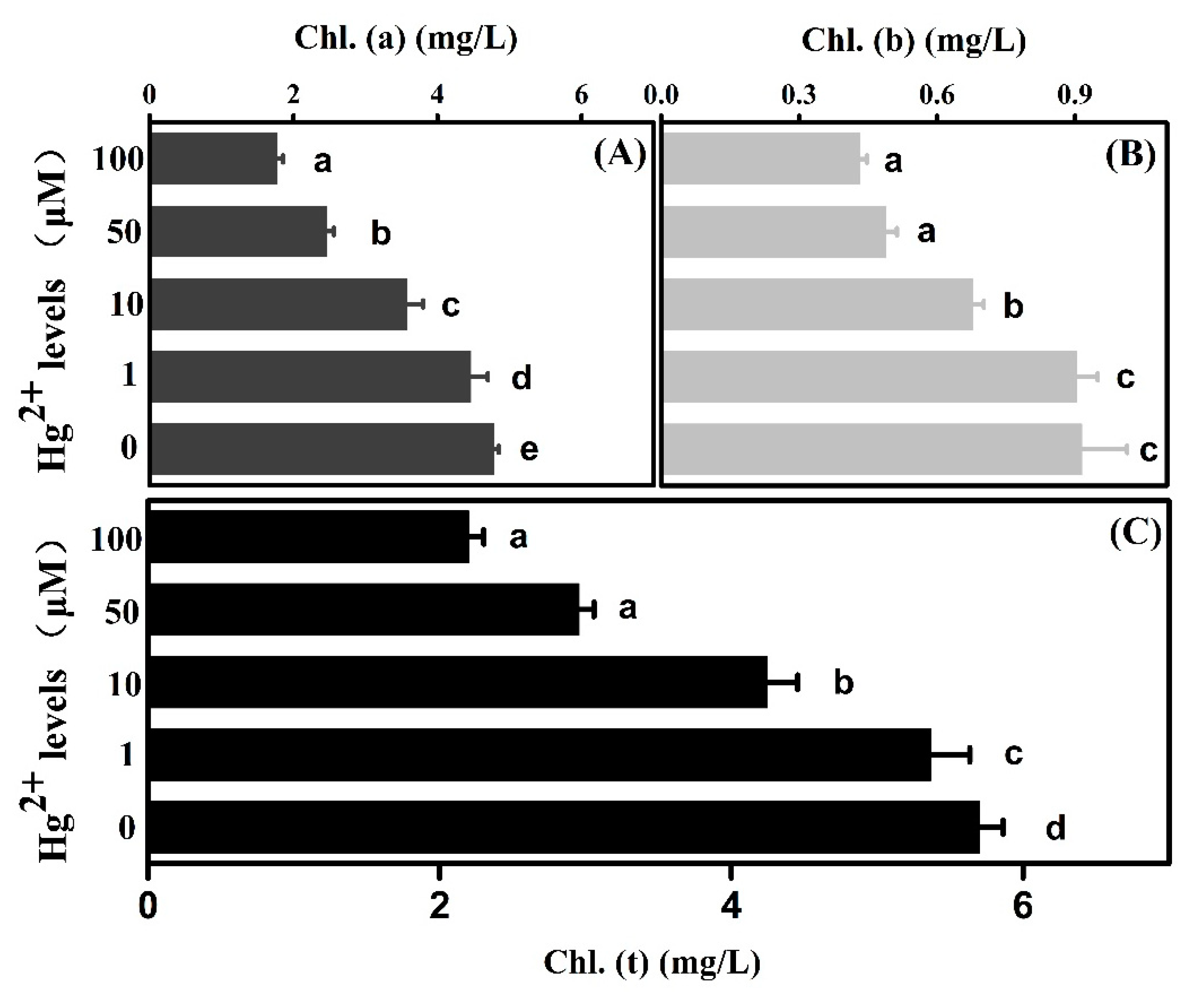
2.3. Effects of Hg on Cotton Assimilation
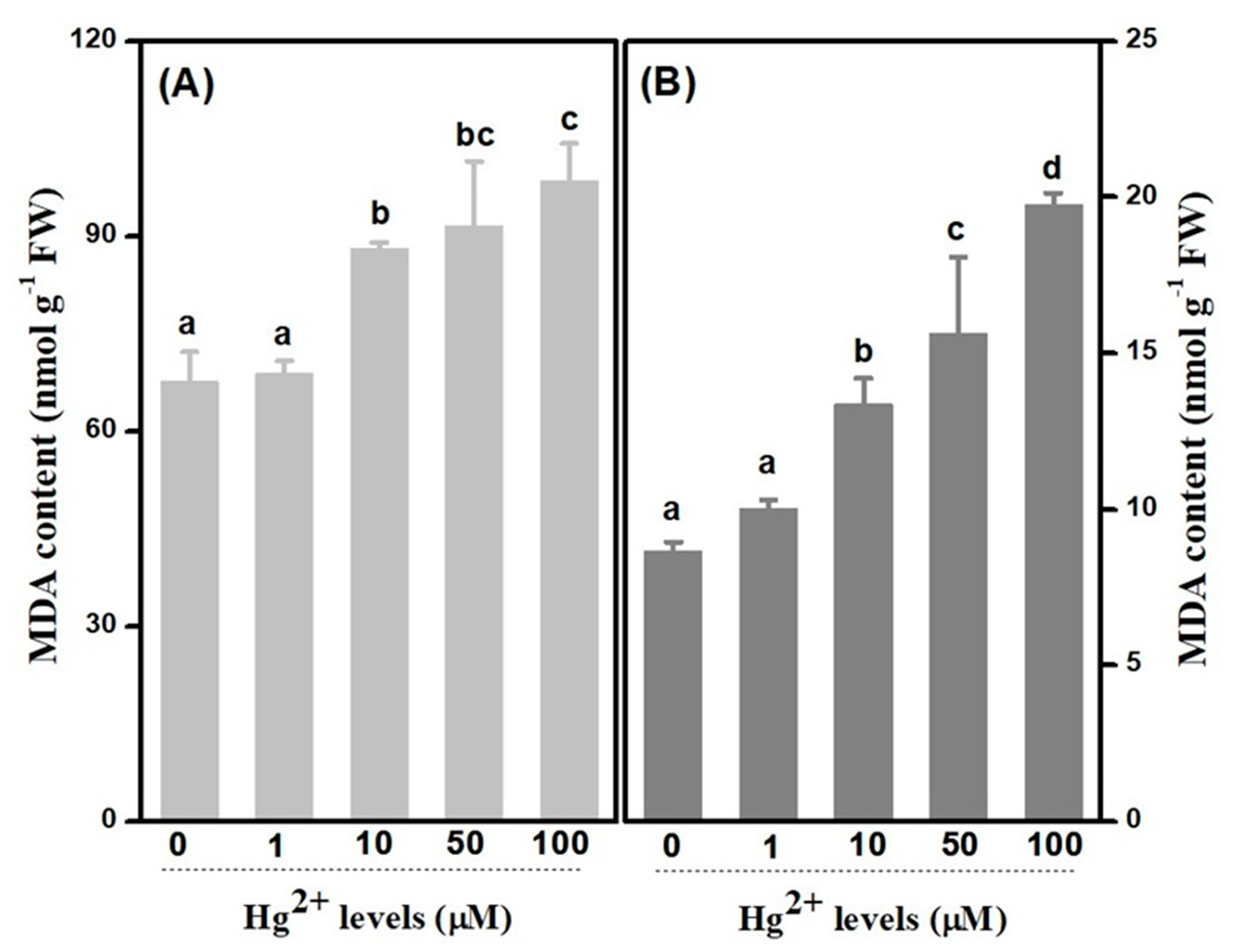
2.4. Membrane Lipid Peroxidation and Leakage
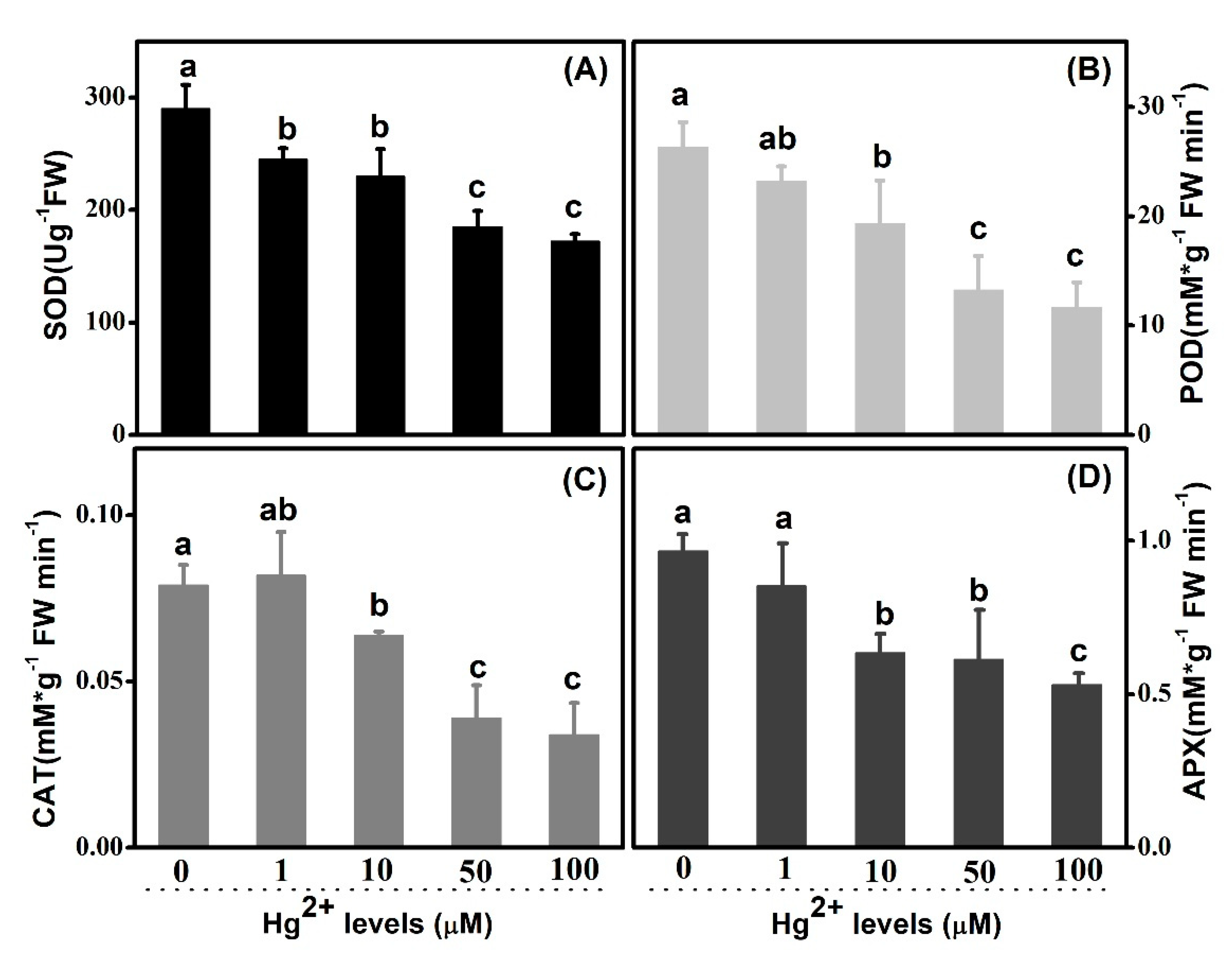
2.5. Antioxidant Enzyme Systems Response
3. Discussion
3.1. Seeds Versus Seedling Plants Exhibited Higher Tolerance to Hg Toxicity
3.2. Damage of Photosynthesis Resulting from Hg Toxicity Accounted for the Sick Growth
3.3. Decrease on inTegrity of Membrane-Symbolled Cellular Injure Resulting from Hg Toxicity
3.4. Hg Disturbed Cellular Redox Equilibrium Involving Depression on Activities of Antioxidant Enzyme System
4. Materials and Methods
4.1. Assay on Seed Germination
4.2. Investigation on Growth and Hg Distribution
4.3. Measures of Pigments and Gas Exchanges in Leaves
4.4. Assay on Malondialdehyde (MDA) Content and Electrolyte Leakage (EL)
4.5. Determination of Antioxidant Enzyme Activities in Cotton Seedlings
4.6. Statistical Analyses Plata
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cao, F.; Wang, R.; Cheng, W.; Zeng, F.; Ahmed, I.M.; Hu, X.; Zhang, G.; Wu, F. Genotypic and environmental variation in cadmium, chromium, lead and copper in rice and approaches for reducing the accumulation. Sci. Total. Environ. 2014, 496, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Suszcynsky, E.M.; Shann, J.R. Phytotoxicity and accumulation of mercury in tobacco subjected to different exposure routes. Environ. Toxicol. Chem. 1995, 14, 61–67. [Google Scholar] [CrossRef]
- Han, F.X.; Banin, A.; Su, Y.; Monts, D.L.; Plodinec, J.M. Industrial age anthropogenic inputs of heavy metals into the pedosphere. Sci. Nat. 2002, 89, 497–504. [Google Scholar] [CrossRef]
- Heaton AC, P.; Rugh, C.L.; Wang, N.; Meagher, R.B. Physiological responses of transgenic merA-Tobacco (Nicotiana tabacum) to foliar and root mercury exposure. Water Air Soil Pollut. 2005, 161, 137–155. [Google Scholar] [CrossRef]
- Han, F.X.; Su, Y.; Monts, D.L.; Waggoner, C.A.; Plodinec, M.J. Binding, distribution, and plant uptake of mercury in a soil from Oak Ridge, Tennessee, USA. Sci. Total Environ. 2006, 368, 753–768. [Google Scholar] [CrossRef]
- Maggio, A.; Joly, R.J.J. Effects of Mercuric Chloride on the Hydraulic conductivity of tomato root system. Plant Physiol. 1995, 109, 331–335. [Google Scholar] [CrossRef]
- Zhang, W.; Tyerman, S.D. Inhibition of water channels by HgCl2 in intact wheat root cells. Plant Physiol. 1999, 120, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Lenti, K.; Fodor, F.; Böddi, B. Mercury Inhibits the Activity of the NADPH:Protochlorophyllide Oxidoreductase (POR). Photosynthetica 2002, 40, 145–151. [Google Scholar] [CrossRef]
- Sahu, G.K.; Upadhyay, S.; Sahoo, B.B. Mercury induced phytotoxicity and oxidative stress in wheat (Triticum aestivum L.) plants. Physiol. Mol. Biol. Plants 2012, 18, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Lu, Y.; Zhu, S. Advance in studies of the mechanism of salt tolerance and controlling of salt damage in upland cotton. Cotton Sci. 2006, 18, 248–254. [Google Scholar]
- Daud Ali, S.; Variath, M.T.; Zhu, S.J. Differential physiological, ultramorphological and metabolic responses of cotton cultivars under cadmium stress. Chemosphere 2013, 3, 2593–2602. [Google Scholar] [CrossRef]
- Daud, M.K.; Quiling, H.; Lei, M.; Ali, B.; Zhu, S.J. Ultrastructural, metabolic and proteomic changes in leaves of upland cotton in response to cadmium stress. Chemosphere 2015, 120, 309–320. [Google Scholar] [CrossRef] [PubMed]
- Daud, M.K.; Mei, L.; Azizullah, A.; Dawood, M.; Ali, I.; Mahmood, Q.; Ullah, W.; Jamil, M.; Zhu, S.J. Leaf-based physiological, metabolic, and ultrastructural changes in cultivated cotton cultivars under cadmium stress mediated by glutathione. Environ. Sci. Pollut. R. 2016, 23, 15551–155564. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.D.; Mei, L.; Ali, B.; Chen, Y.; Cheng, X.; Zhu, S.J. Cadmium-Induced upregulation of lipid peroxidation and reactive oxygen species caused physiological, biochemical, and ultrastructural changes in upland cotton seedlings. Biomed. Res. Int. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mei, L.; Daud, M.K.; Ullah, N.; Ali, S.; Khan, M.; Malik, Z.; Zhu, S.J. Pretreatment with salicylic acid and ascorbic acid significantly mitigate oxidative stress induced by copper in cotton genotypes. Environ. Sci. Pollut. R. 2015, 22, 9922–9931. [Google Scholar] [CrossRef]
- Chaudhuri, K.; Choudhuri, M.A. Effects of short-term NaCl salinity stress on free radical mediated membrane damage in two jute species. Indian J. Exp. Biol. 1993, 3, 327–331. [Google Scholar]
- Liu, D.; Wang, X.; Chen, Z.; Xu, H.; Wang, Y. Influence of mercury on chlorophyll content in winter wheat and mercury bioaccumulation. Plant Soil Environ. 2010, 56, 139–143. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, Q.; Bao, Y.; Li, Y.; Wang, P. Ecotoxicological effects of polycyclic musks and cadmium on seed germination and seedling growth of wheat (Triticum aestivum). J. Environ. Sci. 2010, 22, 1966–1973. [Google Scholar] [CrossRef]
- Rodríguez-Alonso, J.; Sierra, M.J.; Lominchar, M.A.; Millán, R. Effects of mercury on the germination and growth of Quercus ilex L. seedlings. Environ. Sci. Pollut. Res. 2019, 26, 30930–30940. [Google Scholar] [CrossRef]
- Ling, T.; Fangke, Y.; Jun, R. Effect of Mercury to Seed Germination, Coleoptile Growth and Root Elongation of Four Vegetables. Res. J. Phytochem. 2010, 4, 225–233. [Google Scholar] [CrossRef][Green Version]
- Popa, K.; Murariu, M.; Molnar, R.; Schlosser, G.; Cecal, A.; Drochioiu, G. Effect of radioactive and non-radioactive mercury on wheat germination and the anti-toxic role of glutathione. Isot. Environ. Health Stud. 2007, 43, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Ariani, A.; Barozzi, F.; Sebastiani, L.; Di Toppi, L.S.; Di Sansebastiano, G.P.; Andreucci, A. AQUA1 is a mercury sensitive poplar aquaporin regulated at transcriptional and post-translational levels by Zn stress. Plant Physiol. Biochem. 2019, 135, 588–600. [Google Scholar] [CrossRef]
- Magistrali, P.R.; Borges, E.E.D.E.; de Oliveira, J.A.; Faria, J.M.R.; Ataide, G.D.; do Nascimento, J.F. Mercury affects aquaporins activity and germination of the embryonic axis of Schizolobium parahyba (vell.) Blake (fabaceae). Rev. Arvore 2019, 43, 430601. [Google Scholar] [CrossRef]
- Oncel, I.; Kele, Y.; Ustün, A.S. Interactive effects of temperature and heavy metal stress on the growth and some biochemical compounds in wheat seedlings. Environ. Pollut. 2000, 107, 315–320. [Google Scholar] [CrossRef]
- Ketavarapu, S.; Yathavakilla, V.; Caruso, J.A. A study of Se-Hg antagonism in Glycine Max (soybean) roots by size exclusion and reversed phase HPLC-ICOMS. Anal. Bioanal. Chem. 2007, 389, 715–723. [Google Scholar]
- Shanker, K.; Mishra, S.; Srivastava, S.; Srivastava, R.; Daas, S.; Prakash, S.; Srivastava, M.M. Effect of selenite and selenate on plant uptake and translocation of mercury by tomato (Lycopersicum esculentum). Plant Soil 1996, 183, 233–238. [Google Scholar] [CrossRef]
- Ci, D.; Jiang, D.; Wollenweber, B.; Dai, T.; Jing, Q.; Cao, W. Cadmium stress in wheat seedlings: Growth, cadmium accumulation and photosynthesis. Acta Physiol. Plant. 2009, 32, 365–373. [Google Scholar] [CrossRef]
- Rodríguez, E.; Peralta-Videa, J.R.; Israr, M.; Sahi, S.V.; Pelayo, H.; Sánchez-Salcido, B.; Gardea-Torresdey, J.L. Effect of mercury and gold on growth, nutrient uptake, and anatomical changes in Chilopsis linearis. Environ. Exp. Bot. 2009, 65, 253–262. [Google Scholar] [CrossRef]
- Chen, Z.; Pan, Y.; Wang, S.; Ding, Y.; Yang, W.; Zhu, C. Overexpression of a protein disulfide isomerase-like protein from Methanothermobacter thermoautotrophicum enhances mercury tolerance in transgenic rice. Plant Sci. 2012, 197, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Ones, M.Q.; Ruiz-Díez, B.; Fajardo, S.; López-Berdonces, M.A.; Higueras, P.L. Lupinus albus plants acquire mercury tolerance when inoculated with an Hg-resistant Bradyrhizobium strain. Plant Physiol. Bioch. 2013, 73, 168–175. [Google Scholar]
- Zhong, M.; Wang, Y.; Hou, K.; Shu, S.; Sun, J.; Guo, S. TGase positively regulates photosynthesis via activation of Calvin cycle enzymes in tomato. Hortic. Res. 2019, 6, 1–11. [Google Scholar] [CrossRef]
- Ortega-Villasante, C.; Rellan Alvarez, R.N.; Francisca FDel, C.; Carpena-Ruize RN, O.; Herna Ndez, L.E. Cellular damage induced by cadmium and mercury in Medicago sativa. J. Exp. Bot. 2005, 56, 2239–2251. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.; Choudhuri, M.A. Effects of aslicylic acid on heavy metal-induced membrane deterioration mediated by lipoxygenase in rice. Biol. Plant. 1999, 42, 409–415. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Huang, S.Q.; Guo, K.; Mehta, S.K.; Zhang, P.C.; Yang, Z.M. Metabolic adaptations to mercury-induced oxidative stress in roots of Medicago sativa L. J. Inorg. Biochem. 2007, 101, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bhaduri, A.M.; Fulekar, M.H. Antioxidant enzyme responses of plants to heavy metal stress. Rev. Environ. Sci. Bio/Technol. 2012, 11, 55–69. [Google Scholar] [CrossRef]
- Mohammad, A.H.; Pukclai, P.; Jaime AT, S.; Masayuki, F. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxifification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 872875, 1–37. [Google Scholar]
- Israr, M.; Sahi, S.V. Antioxidative responses to mercury in the cell cultures of Sesbania drummondii. Plant Physiol. Bioch. 2006, 44, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Pedron, F.; Petruzzelli, G.; Barbafieri, M.; Tassi, E.; Ambrosini, P.; Patata, L. Mercury Mobilization in a Contaminated Industrial Soil for Phytoremediation. Commun. Soil Sci. Plant Anal. 2011, 42, 2767–2777. [Google Scholar] [CrossRef]
- Moreno-Jiménez, E.; Alosa, J.P.; Esteban, E.; Carpena-Ruiz, R.O. Mercury accumulation and resistance to mercury stress in Rumex induratus and Marrubium vulgare grown in perlite. J. Plant Nutr. Soil. Sci. 2007, 170, 485–494. [Google Scholar] [CrossRef]
- Lin, H.; Santa-Rios, A.; Barst, B.D.; Basu, N.; Bayen, S. Occurrence and bioaccessibility of mercury in commercial rice samples in Montreal (Canada). Food Chem. Toxicol. 2019, 126, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Porra, R.J.; Thompson, W.A.; Kriedemann, P.E. Determination of Accurate Extinction Coefficients and Simultaneous Equations for Assaying Chlorophylls a and b Extracted with Four Different Solvents: Verification of the Concentration of Chlorophyll Standards by Atomic Absorption Spectroscopy. Biochim. Biophys. Acta 1989, 975, 384–394. [Google Scholar] [CrossRef]
- Zhou, W.; Leul, M. Uniconazole-induced alleviation of freezing injury in relation to changes in hormonal balance, enzyme activities and lipid peroxidation in winter rape. Plant Growth Regul. 1998, 26, 41–47. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hg2+ Treatment (μM/L) | Shoots | Roots | ||||||
|---|---|---|---|---|---|---|---|---|
| Fresh Weight (g) | IR (FW) (%) | Dry Weight (g) | IR (DW) (%) | Fresh Weight (g) | IR (FW) (%) | Dry Weight (g) | IR (DW) (%) | |
| 0 | 80.73 ± 1.22 a | 0 | 6.76 ± 0.69 a | 0 | 21.62 ± 0.06 a | 0 | 1.91 ± 0.28 a | 0 |
| 1 | 79.89 ± 1.97 a | 1.04 | 6.54 ± 0.44 a | 3.25 | 20.78 ± 1.03 a | 3.89 | 1.81 ± 0.10 a | 5.24 |
| 10 | 65.55 ± 5.37 b | 18.8 | 5.95 ± 0.69 ab | 11.98 | 17.58 ± 1.36 b | 18.69 | 1.51 ± 0.13 b | 20.94 |
| 50 | 51.42 ± 3.23 c | 36.26 | 5.42 ± 0.11 b | 19.82 | 14.68 ± 1.90 c | 32.10 | 1.07 ± 0.15 c | 43.98 |
| 100 | 50.58 ± 4.14 c | 37.32 | 5.40 ± 0.10 b | 20.19 | 13.91 ± 0.69 c | 36.16 | 0.94 ± 0.07 c | 49.21 |
| Hg2+ Treatments (μM) | Hg Concentration (μgg−1 DW) | CI (Concentration Index) | TF (Translocation Factor) | ||||
|---|---|---|---|---|---|---|---|
| Leaves | Stems | Root | Leaves | Stems | Roots | ||
| 0 | 1.45 ± 0.10 a | 1.63 ± 0.28 a | 5.16 ± 0.05 a | 1.00 | 1.00 | 1.00 | 0.60 |
| 1 | 2.23 ± 0.12 b | 2.28 ± 0.19 b | 28.60 ± 0.36 b | 1.54 | 1.40 | 5.54 | 0.16 |
| 10 | 3.86 ± 0.10 b | 2.42 ± 0.01 c | 54.81 ± 0.33 c | 2.66 | 1.48 | 10.62 | 0.11 |
| 50 | 9.37 ± 0.45 c | 4.05 ± 0.06 d | 288.23 ± 20.75 d | 6.46 | 2.48 | 55.86 | 0.05 |
| 100 | 10.47 ± 0.27 d | 6.56 ± 0.01 e | 303.89 ± 11.58 d | 7.22 | 4.02 | 58.89 | 0.06 |
| Hg2+ Levels (μM) | Relative Conductivity (RC) (%) | |
|---|---|---|
| Leaves | Roots | |
| 0 | 15.54 ± 1.83 a | 20.21 ± 3.53 a |
| 1 | 15.85 ± 2.83 a | 35.22 ± 3.59 b |
| 10 | 30.30 ± 7.81 b | 38.64 ± 0.87 b |
| 50 | 53.95 ± 5.07 c | 52.10 ± 7.73 c |
| 100 | 78.36 ± 8.68 d | 54.01 ± 5.97 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, L.; Zhu, Y.; Zhang, X.; Zhou, X.; Zhong, Z.; Li, H.; Li, Y.; Li, X.; Daud, M.K.; Chen, J.; et al. Mercury-Induced Phytotoxicity and Responses in Upland Cotton (Gossypium hirsutum L.) Seedlings. Plants 2021, 10, 1494. https://doi.org/10.3390/plants10081494
Mei L, Zhu Y, Zhang X, Zhou X, Zhong Z, Li H, Li Y, Li X, Daud MK, Chen J, et al. Mercury-Induced Phytotoxicity and Responses in Upland Cotton (Gossypium hirsutum L.) Seedlings. Plants. 2021; 10(8):1494. https://doi.org/10.3390/plants10081494
Chicago/Turabian StyleMei, Lei, Yueyi Zhu, Xianwen Zhang, Xiujuan Zhou, Zhentao Zhong, Huazu Li, Yingjun Li, Xiaohu Li, Muhammad Khan Daud, Jinhong Chen, and et al. 2021. "Mercury-Induced Phytotoxicity and Responses in Upland Cotton (Gossypium hirsutum L.) Seedlings" Plants 10, no. 8: 1494. https://doi.org/10.3390/plants10081494
APA StyleMei, L., Zhu, Y., Zhang, X., Zhou, X., Zhong, Z., Li, H., Li, Y., Li, X., Daud, M. K., Chen, J., & Zhu, S. (2021). Mercury-Induced Phytotoxicity and Responses in Upland Cotton (Gossypium hirsutum L.) Seedlings. Plants, 10(8), 1494. https://doi.org/10.3390/plants10081494