
Involvement of Auxin Biosynthesis and Transport in the Antheridium and Prothalli Formation in Lygodium japonicum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
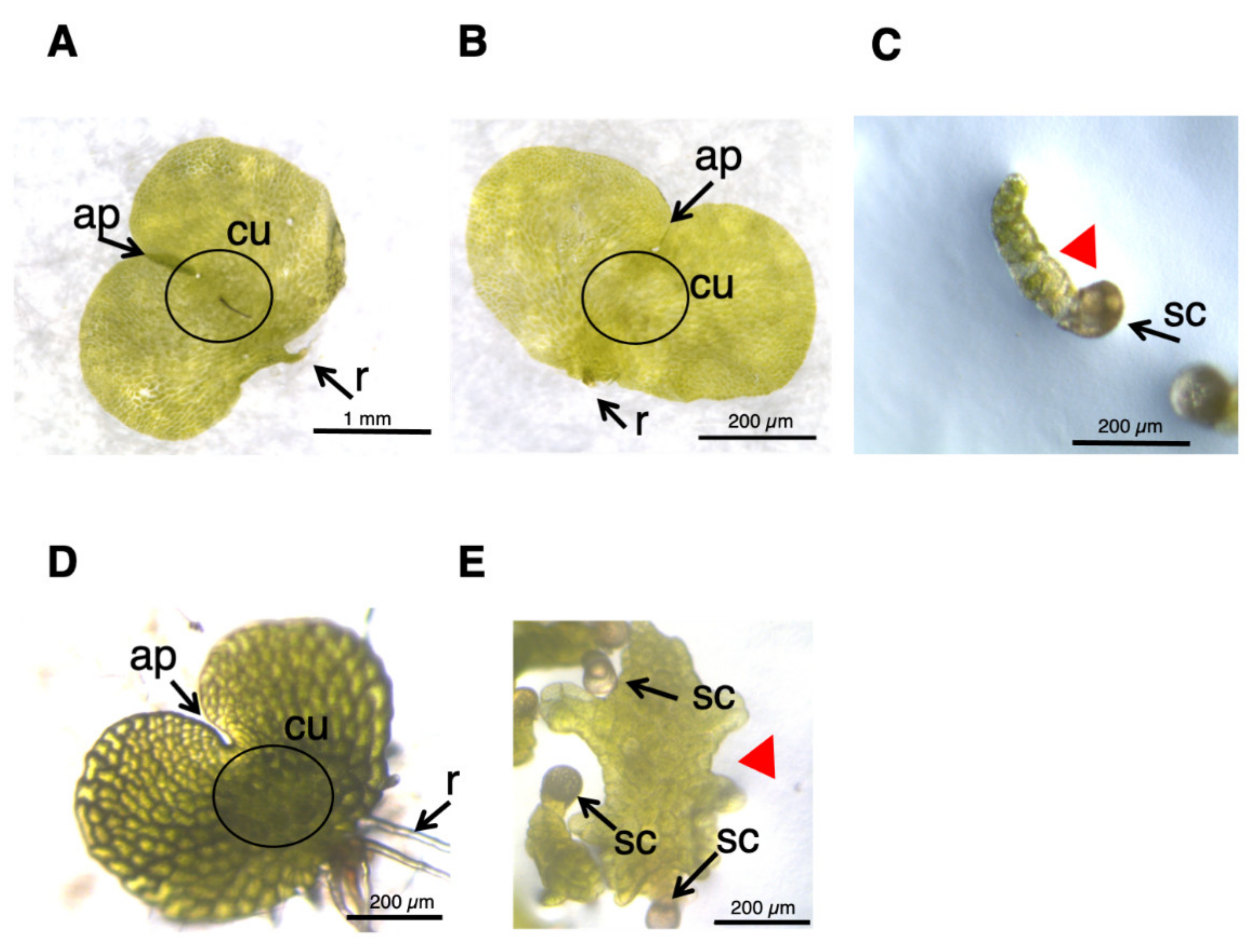{kind=link}
Abstract
1. Introduction
2. Results and Discussion
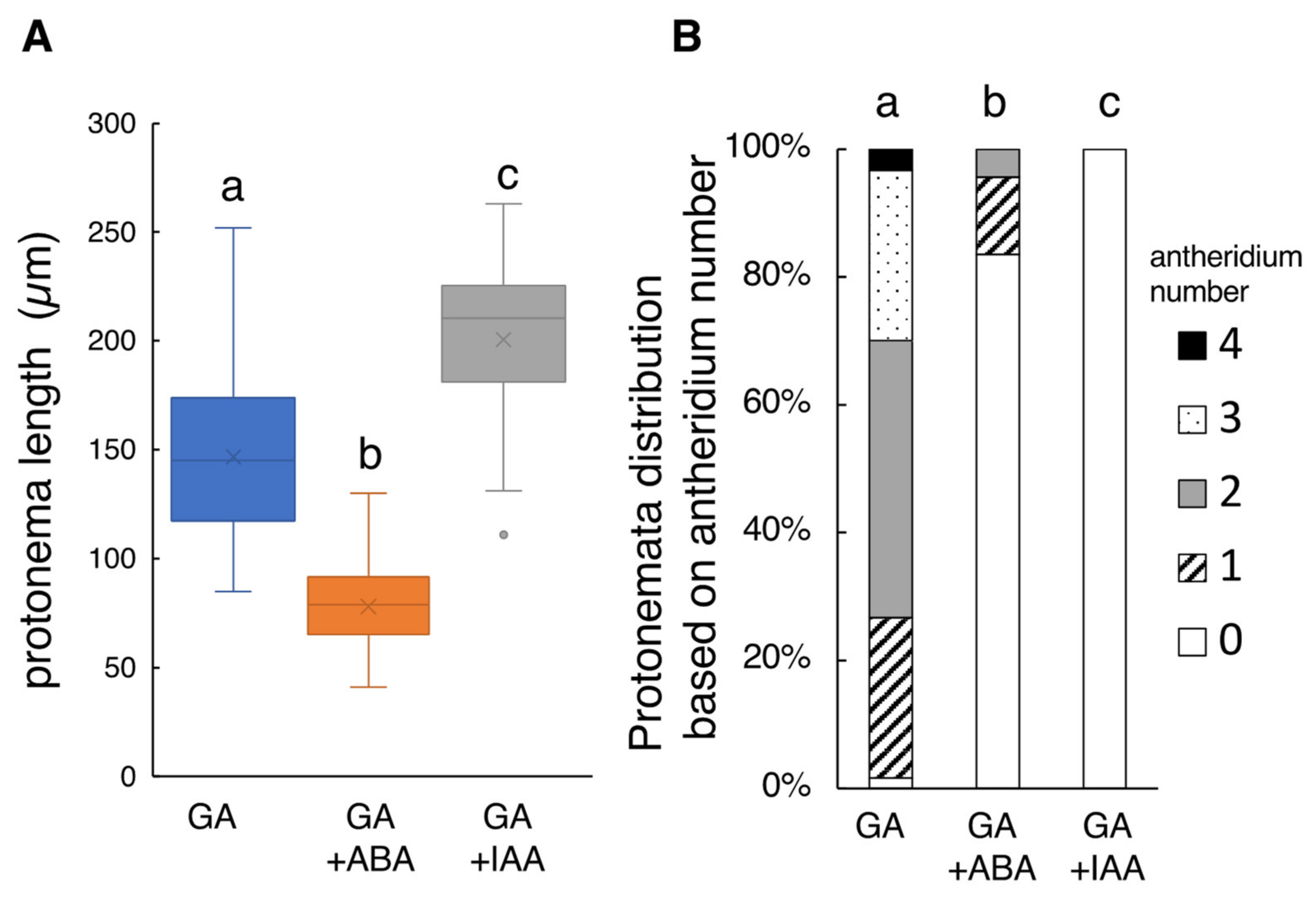
2.1. Effect of ABA and IAA on GA4-Me-Medaited Protonema Elongation and Antheridium Formation of L. japonicum
2.2. Effect of Transferring of IAA-Treated Cells to an IAA-Free Medium
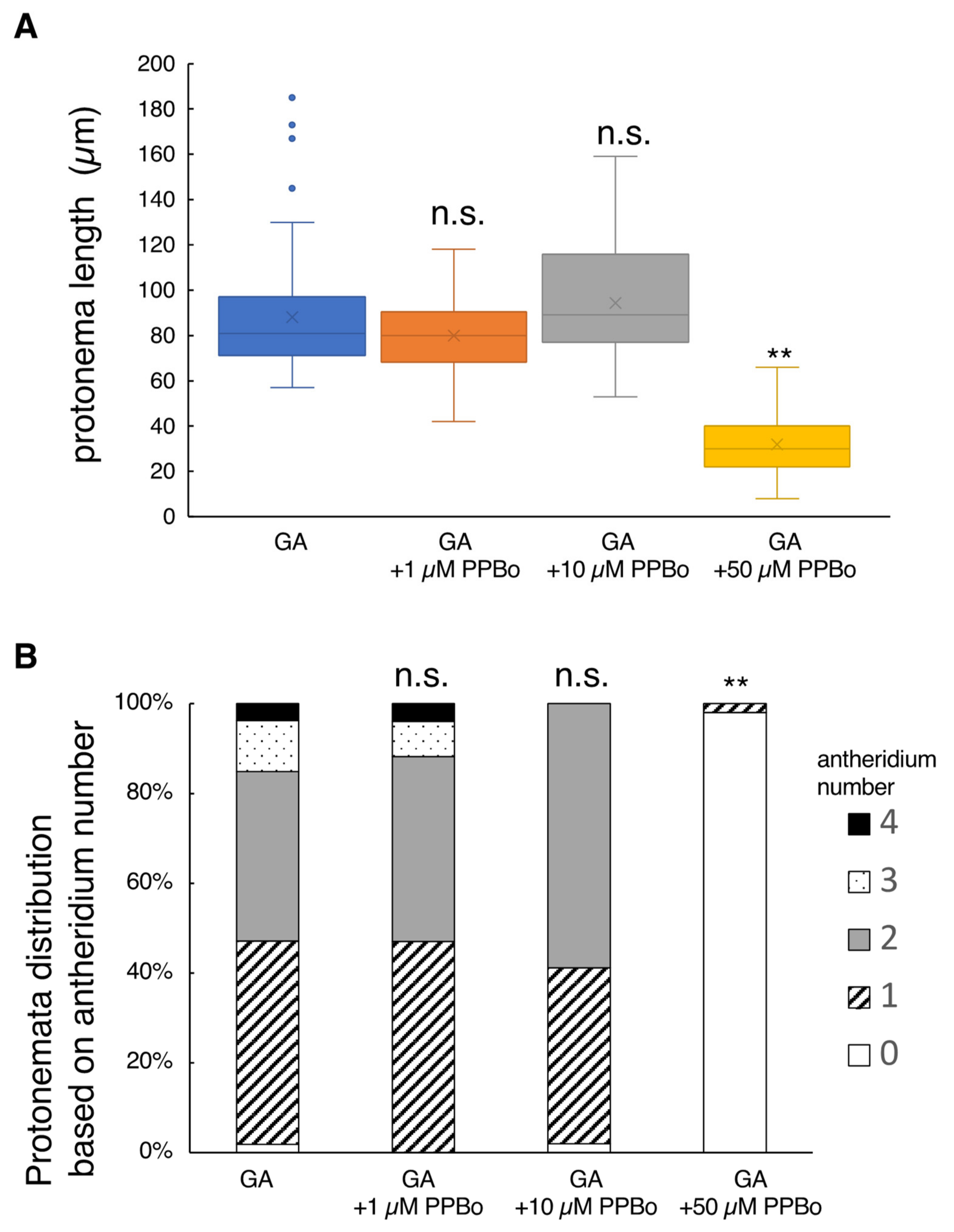
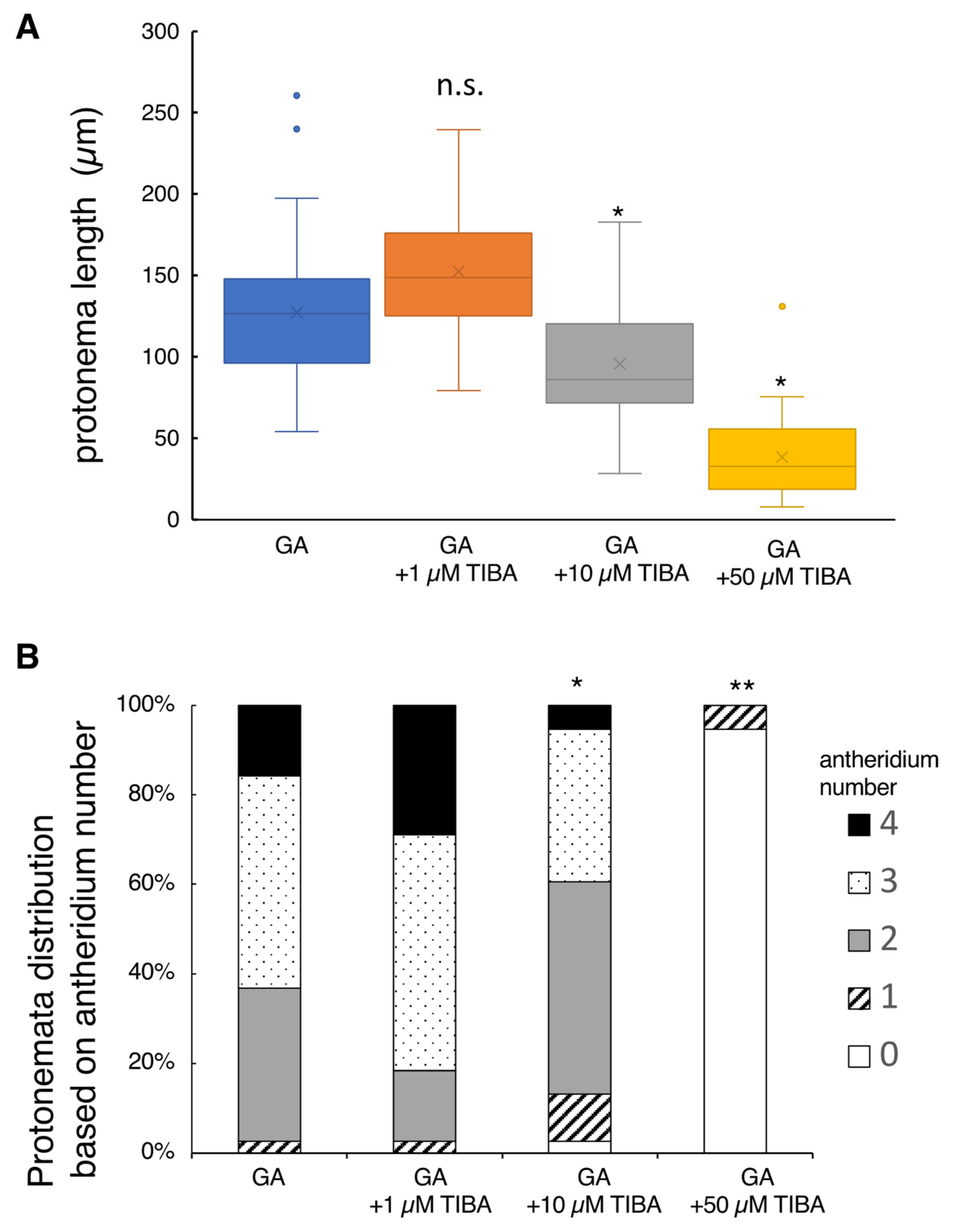
2.3. Effect of Auxin Inhibitors on Protonema Elongation and Antheridium Formation
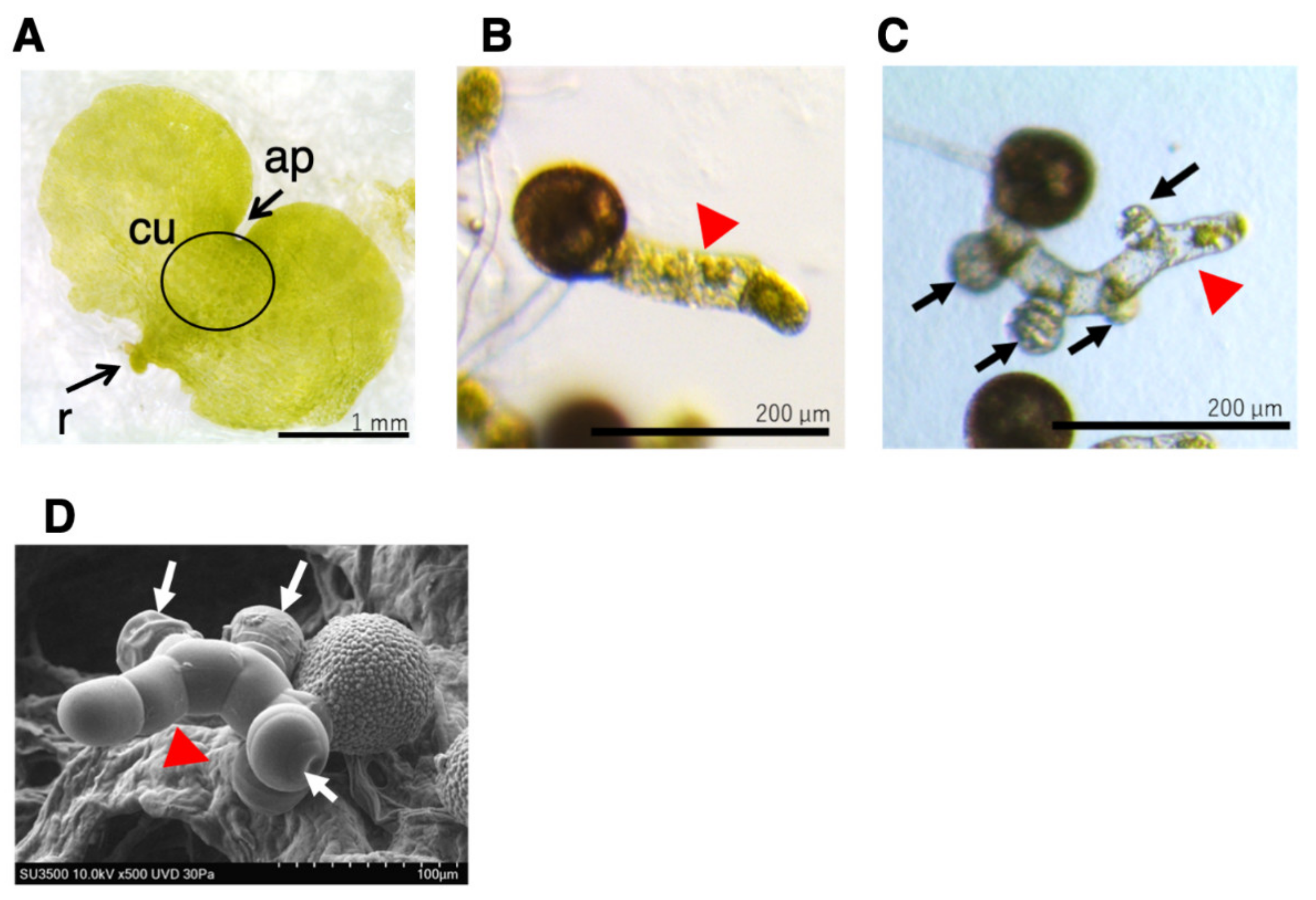
2.4. Effects of Auxin Inhibitors on the Morphology of L. japonicum Prothalli
3. Materials and Methods
3.1. Plant Materials and Culture Conditions
3.2. Measurement of Protonema Elongation and Antheridium Formation
3.3. Effect of Auxin Inhibitors on Prothalli Development
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vasco, A.; Moran, R.C.; Ambrose, B.A. The Evolution, Morphology, and Development of Fern Leaves. Front. Plant Sci. 2013, 4, 345. [Google Scholar] [CrossRef] [PubMed]
- Kenrick, P. The Relationships of Vascular Plants. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2000, 355, 847–855. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, J.; Fu, X.-X.; Li, R.-Q.; Zhao, X.; Liu, Y.; Li, M.-H.; Zwaenepoel, A.; Ma, H.; Goffinet, B.; Guan, Y.-L.; et al. The Hornwort Genome and Early Land Plant Evolution. Nat. Plants 2020, 6, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Gifford, E.M.; Foster, A.S. Morphology and Evolution of Vascular Plants, 3rd ed.; Freeman: New York, NY, USA, 1988; ISBN 0-7167-1946-0. [Google Scholar]
- Takahashi, N.; Kami, C.; Ota, I.; Morita, N.; Imaichi, R. Developmental Morphology of the Typical Cordate Gametophyte of a Homosporous Leptosporangiate Fern, Lygodium japonicum (Lygodiaceae), Focusing on the Initial Cell Behavior of Two Distinct Meristems. Am. J. Bot. 2015, 102, 197–207. [Google Scholar] [CrossRef]
- Takeno, K.; Furuya, M. Bioassay of Antheridiogen in Lygodium japonicum. Dev. Growth Differ. 1975, 17, 9–18. [Google Scholar] [CrossRef]
- Takeno, K.; Furuya, M. Inhibitory Effect of Gibberellins on Archegonial Differentiation in Lygodium japonicum. Physiol. Plant. 1977, 39, 135–138. [Google Scholar] [CrossRef]
- Yamauchi, T.; Oyama, N.; Yamane, H.; Murofushi, N.; Schraudolf, H.; Pour, M.; Furber, M.; Mander, L.N. Identification of Antheridiogens in Lygodium circinnatum and Lygodium flexuosum. Plant Physiol. 1996, 111, 741–745. [Google Scholar] [CrossRef]
- Tanaka, J.; Yano, K.; Aya, K.; Hirano, K.; Takehara, S.; Koketsu, E.; Ordonio, R.L.; Park, S.-H.; Nakajima, M.; Ueguchi-Tanaka, M.; et al. Antheridiogen Determines Sex in Ferns via a Spatiotemporally Split Gibberellin Synthesis Pathway. Science 2014, 346, 469–473. [Google Scholar] [CrossRef]
- Ueguchi-Tanaka, M.; Ashikari, M.; Nakajima, M.; Itoh, H.; Katoh, E.; Kobayashi, M.; Chow, T.; Hsing, Y.C.; Kitano, H.; Yamaguchi, I.; et al. GIBBERELLIN INSENSITIVE DWARF1 Encodes a Soluble Receptor for Gibberellin. Nature 2005, 437, 693–698. [Google Scholar] [CrossRef]
- Ueguchi-Tanaka, M.; Nakajima, M.; Katoh, E.; Ohmiya, H.; Asano, K.; Saji, S.; Hongyu, X.; Ashikari, M.; Kitano, H.; Yamaguchi, I.; et al. Molecular Interactions of a Soluble Gibberellin Receptor, GID1, with a Rice DELLA Protein, SLR1, and Gibberellin. Plant Cell 2007, 19, 2140–2155. [Google Scholar] [CrossRef]
- Yamane, H.; Takahashi, N.; Takeno, K.; Furuya, M. Identification of Gibberellin A9 Methyl Ester as a Natural Substance Regulating Formation of Reproductive Organs in Lygodium japonicum. Planta 1979, 147, 251–256. [Google Scholar] [CrossRef]
- Yamane, H.; Yamaguchi, I.; Kobayashi, M.; Takahashi, M.; Sato, Y.; Takahashi, N.; Iwatsuki, K.; Phinney, B.O.; Spray, C.R.; Gaskin, P.; et al. Identification of Ten Gibberellins from Sporophytes of the Tree Fern, Cyathea australis. Plant Physiol. 1985, 78, 899–903. [Google Scholar] [CrossRef]
- Takeno, K.; Yamane, H.; Yamauchi, T.; Takahashi, N.; Furber, M.; Mander, L.N. Biological Activities of the Methyl Ester of Gibberellin A73, a Novel and Principal Antheridiogen in Lygodium japonicum. Plant Cell Physiol. 1989, 30, 201–205. [Google Scholar] [CrossRef]
- Sugai, M.; Takeno, K.; Furuya, M. Diverse Responses of Spores in the Light-Dependent Germination of Lygodium japonicum. Plant Sci. Lett. 1977, 8, 333–338. [Google Scholar] [CrossRef]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The Main Auxin Biosynthesis Pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef]
- Sugawara, S.; Mashiguchi, K.; Tanaka, K.; Hishiyama, S.; Sakai, T.; Hanada, K.; Kinoshita-Tsujimura, K.; Yu, H.; Dai, X.; Takebayashi, Y.; et al. Distinct Characteristics of Indole-3-Acetic Acid and Phenylacetic Acid, Two Common Auxins in Plants. Plant Cell Physiol. 2015, 56, 1641–1654. [Google Scholar] [CrossRef]
- Kato, H.; Ishizaki, K.; Kouno, M.; Shirakawa, M.; Bowman, J.L.; Nishihama, R.; Kohchi, T. Auxin-Mediated Transcriptional System with a Minimal Set of Components Is Critical for Morphogenesis through the Life Cycle in Marchantia polymorpha. PLoS Genet. 2015, 11, e1005084. [Google Scholar] [CrossRef]
- Landberg, K.; Pederson, E.R.A.; Viaene, T.; Bozorg, B.; Friml, J.; Jönsson, H.; Thelander, M.; Sundberg, E. The Moss Physcomitrella patens Reproductive Organ Development Is Highly Organized, Affected by the Two SHI/STY Genes and by the Level of Active Auxin in the SHI/STY Expression Domain. Plant Physiol. 2013, 162, 1406–1419. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, Y.; Liu, W.; Wang, H.; Wen, S.; Zhang, Y.; Xu, L. Molecular Evolution of Auxin-Mediated Root Initiation in Plants. Mol. Biol. Evol. 2020, 37, 1387–1393. [Google Scholar] [CrossRef]
- Hickok, L.G.; Kiriluk, R.M. Effects of Auxins on Gametophyte Development and Sexual Differentiation in the Fern Ceratopteris Thalictroides (L.) Brongn. Bot. Gaz. 1984, 145, 37–42. [Google Scholar] [CrossRef]
- Yamane, H.; Sato, Y.; Takahashi, N.; Takeno, K.; Furuya, M. Endogenous Inhibitors for Spore Germination in Lygodium japonicum and Their Inhibitory Effects on Pollen Germinations in Camellia Japonica and Camellia sinensis. Agric. Biol. Chem. 1980, 44, 1697–1699. [Google Scholar] [CrossRef]
- Hõrak, H.; Kollist, H.; Merilo, E. Fern Stomatal Responses to ABA and CO2 Depend on Species and Growth Conditions. Plant Physiol. 2017, 174, 672–679. [Google Scholar] [CrossRef]
- McAdam, S.A.M.; Brodribb, T.J.; Banks, J.A.; Hedrich, R.; Atallah, N.M.; Cai, C.; Geringer, M.A.; Lind, C.; Nichols, D.S.; Stachowski, K.; et al. Abscisic Acid Controlled Sex before Transpiration in Vascular Plants. Proc. Natl. Acad. Sci. USA 2016, 113, 12862–12867. [Google Scholar] [CrossRef]
- Nishimura, T.; Hayashi, K.; Suzuki, H.; Gyohda, A.; Takaoka, C.; Sakaguchi, Y.; Matsumoto, S.; Kasahara, H.; Sakai, T.; Kato, J.; et al. Yucasin Is a Potent Inhibitor of YUCCA, a Key Enzyme in Auxin Biosynthesis. Plant J. 2014, 77, 352–366. [Google Scholar] [CrossRef]
- Kakei, Y.; Yamazaki, C.; Suzuki, M.; Nakamura, A.; Sato, A.; Ishida, Y.; Kikuchi, R.; Higashi, S.; Kokudo, Y.; Ishii, T.; et al. Small-Molecule Auxin Inhibitors That Target YUCCA Are Powerful Tools for Studying Auxin Function. Plant J. 2015, 84, 827–837. [Google Scholar] [CrossRef]
- Suzuki, H.; Yokawa, K.; Nakano, S.; Yoshida, Y.; Fabrissin, I.; Okamoto, T.; Baluška, F.; Koshiba, T. Root Cap-Dependent Gravitropic U-Turn of Maize Root Requires Light-Induced Auxin Biosynthesis via the YUC Pathway in the Root Apex. J. Exp. Bot. 2016, 67, 4581–4591. [Google Scholar] [CrossRef]
- Aya, K.; Kobayashi, M.; Tanaka, J.; Ohyanagi, H.; Suzuki, T.; Yano, K.; Takano, T.; Yano, K.; Matsuoka, M. De Novo Transcriptome Assembly of a Fern, Lygodium japonicum, and a Web Resource Database, Ljtrans DB. Plant Cell Physiol. 2015, 56, e5. [Google Scholar] [CrossRef]
- Swarup, R.; Parry, G.; Graham, N.; Allen, T.; Bennett, M. Auxin Cross-Talk: Integration of Signalling Pathways to Control Plant Development. In Auxin Molecular Biology; Perrot-Rechenmann, C., Hagen, G., Eds.; Springer: Dordrecht, The Netherlands, 2002; pp. 411–426. ISBN 978-94-010-3917-8. [Google Scholar]
- Weiss, D.; Ori, N. Mechanisms of Cross Talk between Gibberellin and Other Hormones. Plant Physiol. 2007, 144, 1240–1246. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant Hormone-Mediated Regulation of Stress Responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Fu, X.; Harberd, N.P. Auxin Promotes Arabidopsis Root Growth by Modulating Gibberellin Response. Nature 2003, 421, 740–743. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, T.; Kadota, A.; Hasebe, M.; Wada, M. Cryptochrome Light Signals Control Development to Suppress Auxin Sensitivity in the Moss Physcomitrella patens. Plant Cell 2002, 14, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Horie, K.; Hiwatashi, Y.; Kawaide, H.; Yamaguchi, S.; Hanada, A.; Nakashima, T.; Nakajima, M.; Mander, L.N.; Yamane, H.; et al. Endogenous Diterpenes Derived from Ent-Kaurene, a Common Gibberellin Precursor, Regulate Protonema Differentiation of the Moss Physcomitrella patens. Plant Physiol. 2010, 153, 1085–1097. [Google Scholar] [CrossRef] [PubMed]
- Hirano, K.; Nakajima, M.; Asano, K.; Nishiyama, T.; Sakakibara, H.; Kojima, M.; Katoh, E.; Xiang, H.; Tanahashi, T.; Hasebe, M.; et al. The GID1-Mediated Gibberellin Perception Mechanism Is Conserved in the Lycophyte Selaginella moellendorffii but Not in the Bryophyte Physcomitrella patens. Plant Cell 2007, 19, 3058–3079. [Google Scholar] [CrossRef]
- Anterola, A.; Shanle, E.; Mansouri, K.; Schuette, S.; Renzaglia, K. Gibberellin Precursor Is Involved in Spore Germination in the Moss Physcomitrella patens. Planta 2009, 229, 1003–1007. [Google Scholar] [CrossRef]
- Yokota, T.; Ohnishi, T.; Shibata, K.; Asahina, M.; Nomura, T.; Fujita, T.; Ishizaki, K.; Kohchi, T. Occurrence of Brassinosteroids in Non-Flowering Land Plants, Liverwort, Moss, Lycophyte and Fern. Phytochemistry 2017, 136, 46–55. [Google Scholar] [CrossRef]
- Clouse, S.D.; Sasse, J.M. BRASSINOSTEROIDS: Essential Regulators of Plant Growth and Development. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 427–451. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohishi, N.; Hoshika, N.; Takeda, M.; Shibata, K.; Yamane, H.; Yokota, T.; Asahina, M. Involvement of Auxin Biosynthesis and Transport in the Antheridium and Prothalli Formation in Lygodium japonicum. Plants 2021, 10, 2709. https://doi.org/10.3390/plants10122709
Ohishi N, Hoshika N, Takeda M, Shibata K, Yamane H, Yokota T, Asahina M. Involvement of Auxin Biosynthesis and Transport in the Antheridium and Prothalli Formation in Lygodium japonicum. Plants. 2021; 10(12):2709. https://doi.org/10.3390/plants10122709
Chicago/Turabian StyleOhishi, Natsumi, Nanami Hoshika, Mizuho Takeda, Kyomi Shibata, Hisakazu Yamane, Takao Yokota, and Masashi Asahina. 2021. "Involvement of Auxin Biosynthesis and Transport in the Antheridium and Prothalli Formation in Lygodium japonicum" Plants 10, no. 12: 2709. https://doi.org/10.3390/plants10122709
APA StyleOhishi, N., Hoshika, N., Takeda, M., Shibata, K., Yamane, H., Yokota, T., & Asahina, M. (2021). Involvement of Auxin Biosynthesis and Transport in the Antheridium and Prothalli Formation in Lygodium japonicum. Plants, 10(12), 2709. https://doi.org/10.3390/plants10122709