
Ulva intestinalis Extract Acts as Biostimulant and Modulates Metabolites and Hormone Balance in Basil (Ocimum basilicum L.) and Parsley (Petroselinum crispum L.)


 , ,
, ,  , , and
, , and 
Abstract
:1. Introduction
2. Results
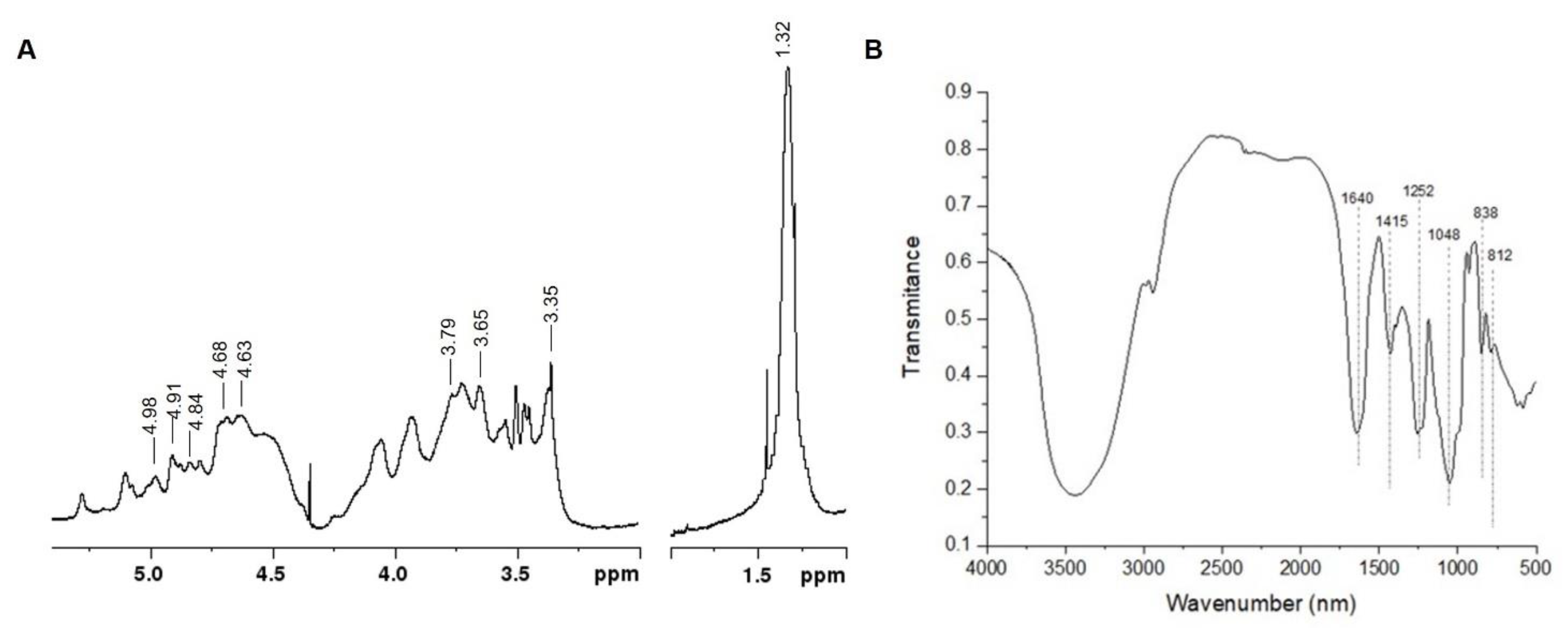
2.1. Ulva intestinalis Extract (UIE) Analyses
2.2. Elicitation of Leaves with UIE: Biochemical Analyses
2.3. Hormonal Profiles of Elicited Leaves
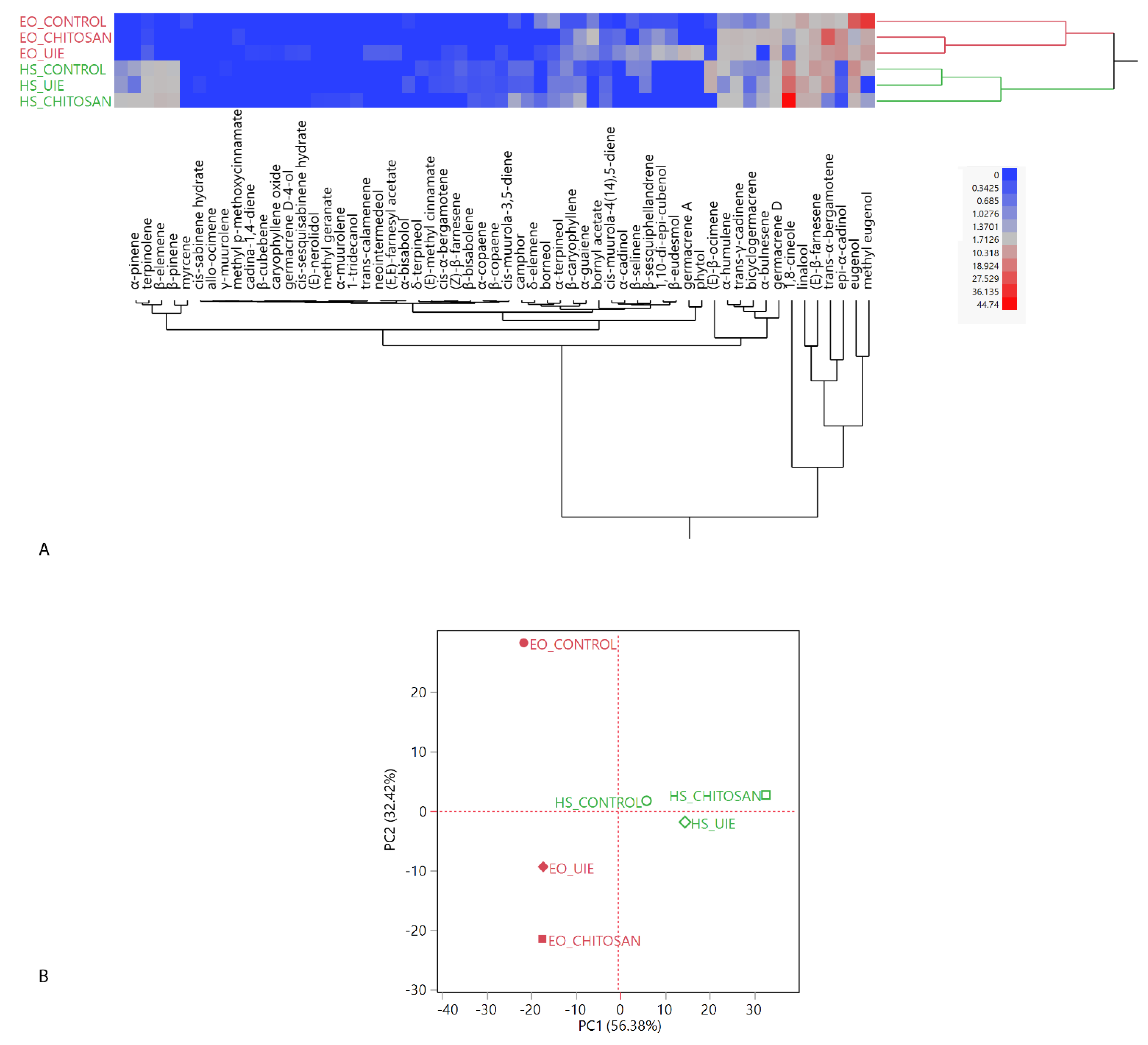
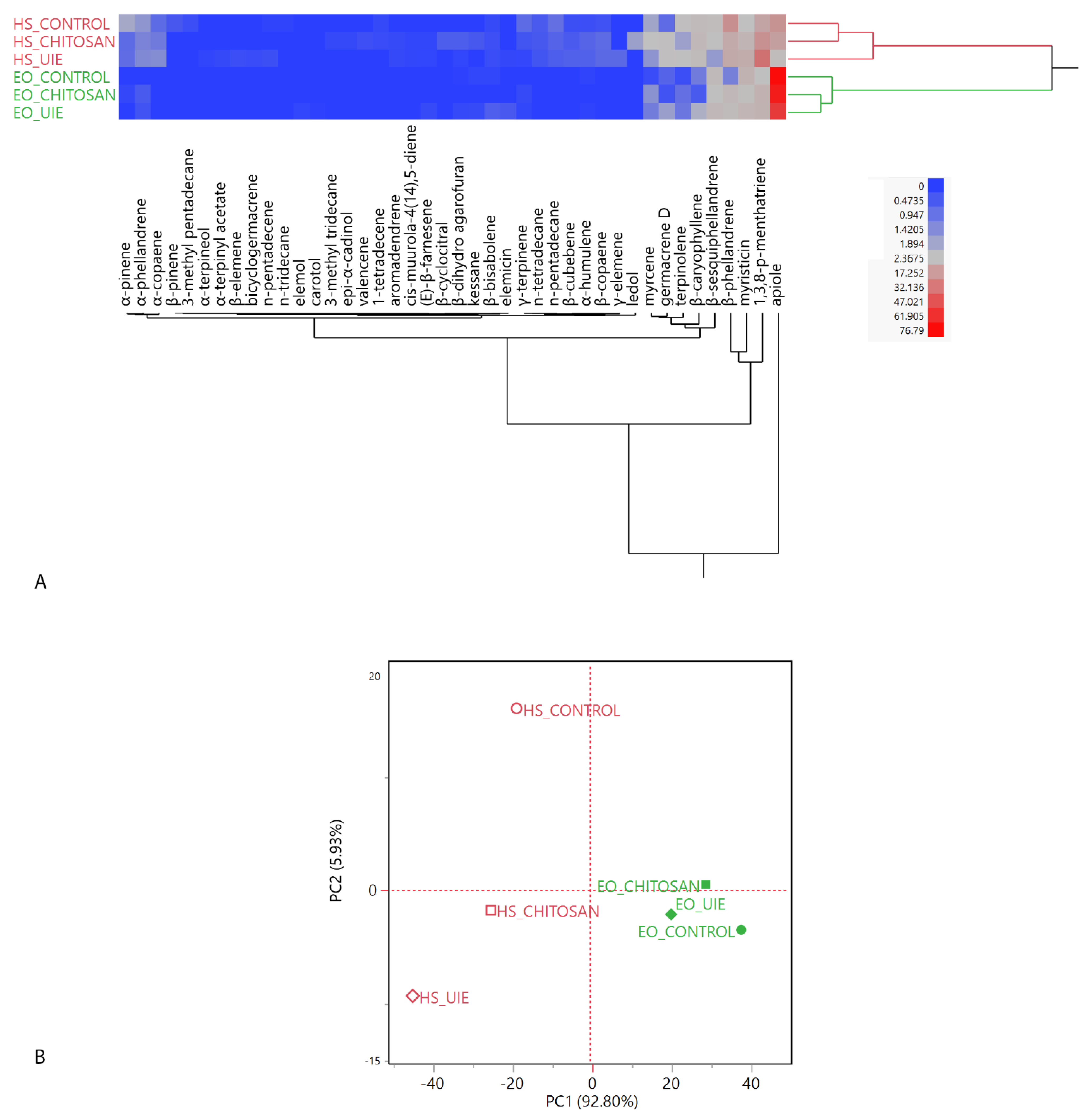
2.4. Volatiles Analyses: Headspaces (HSs) and Essential Oils (EOs)
2.4.1. Basil
2.4.2. Parsley
3. Discussion
4. Materials and Methods
4.1. Algal Material
4.2. Preparation of Ulva Intestinalis Extract (UIE)
4.3. UIE Composition Analyses
4.4. High-Pressure Size-Exclusion Chromatography (HPSEC) Coupled to Refractive Index (RI) and Multi-Angle Laser Light Scattering (MALLS) Detectors Analysis of UIE
4.5. NMR and FTIR Analyses of UIE
4.6. Plant Material and Treatment
4.7. Biochemical Analyses of Leaves
4.8. Hormonal Level Quantification
4.9. Headspace Solid Phase Micro-Extraction (HS-SPME)
4.10. Essential oil (EO) Hydrodistillations
4.11. Gas Chromatography-Electron Impact Mass Spectrometry (GC–EIMS) Analyses and Peak Identifications
4.12. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vera, J.; Castro, J.; Gonzalez, A.; Moenne, A. Seaweed Polysaccharides and Derived Oligosaccharides Stimulate Defense Responses and Protection Against Pathogens in Plants. Mar. Drugs 2011, 9, 2514–2525. [Google Scholar] [CrossRef] [PubMed]
- Rabea, E.I.; Badawy, M.E.-T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as Antimicrobial Agent: Applications and Mode of Action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Trouvelot, S.; Héloir, M.-C.; Poinssot, B.; Gauthier, A.; Paris, F.; Guillier, C.; Combier, M.; Trdá, L.; Daire, X.; Adrian, M. Carbohydrates in plant immunity and plant protection: Roles and potential application as foliar sprays. Front. Plant Sci. 2014, 5, 592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kidgell, J.T.; Magnusson, M.; de Nys, R.; Glasson, C.R.K. Ulvan: A systematic review of extraction, composition and function. Algal Res. 2019, 39, 101422. [Google Scholar] [CrossRef]
- Alves, A.; Sousa, R.A.; Reis, R.L. A practical perspective on ulvan extracted from green algae. J. Appl. Phycol. 2013, 25, 407–424. [Google Scholar] [CrossRef] [Green Version]
- Berri, M.; Olivier, M.; Holbert, S.; Dupont, J.; Demais, H.; Le Goff, M.; Collen, P.N. Ulvan from Ulva armoricana (Chlorophyta) activates the PI3K/Akt signalling pathway via TLR4 to induce intestinal cytokine production. Algal Res. 2017, 28, 39–47. [Google Scholar] [CrossRef]
- Jaulneau, V.; Lafitte, C.; Jacquet, C.; Fournier, S.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.-T.; Dumas, B. Ulvan, a Sulfated Polysaccharide from Green Algae, Activates Plant Immunity through the Jasmonic Acid Signaling Pathway. J. Biomed. Biotechnol. 2010, 2010, 525291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Massironi, A.; Morelli, A.; Grassi, L.; Puppi, D.; Braccini, S.; Maisetta, G.; Esin, S.; Batoni, G.; Della Pina, C.; Chiellini, F. Ulvan as novel reducing and stabilizing agent from renewable algal biomass: Application to green synthesis of silver nanoparticles. Carbohydr. Polym. 2019, 203, 310–321. [Google Scholar] [CrossRef]
- Melcher, R.L.J.; Neumann, M.; Fuenzalida Werner, J.P.; Gröhn, F.; Moerschbacher, B.M. Revised domain structure of ulvan lyase and characterization of the first ulvan binding domain. Sci. Rep. 2017, 7, 44115. [Google Scholar] [CrossRef] [Green Version]
- Cluzet, S.; Torregrosa, C.; Jacquet, C.; Lafitte, C.; Fournier, J.; Mercier, L.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.-T.; Dumas, B. Gene expression profiling and protection of Medicago truncatula against a fungal infection in response to an elicitor from green algae Ulva spp. Plant Cell Environ. 2004, 27, 917–928. [Google Scholar] [CrossRef]
- Paulert, R.; Talamini, V.; Cassolato, J.E.F.; Duarte, M.E.R.; Noseda, M.D.; Smania, A.; Stadnik, M.J. Effects of sulfated polysaccharide and alcoholic extracts from green seaweed Ulva fasciata on anthracnose severity and growth of common bean (Phaseolus vulgaris L.). J. Plant Dis. Prot. 2009, 116, 263–270. [Google Scholar] [CrossRef]
- Jaulneau, V.; Lafitte, C.; Corio-Costet, M.-F.; Stadnik, M.J.; Salamagne, S.; Briand, X.; Esquerré-Tugayé, M.-T.; Dumas, B. An Ulva armoricana extract protects plants against three powdery mildew pathogens. Eur. J. Plant Pathol. 2011, 131, 393–401. [Google Scholar] [CrossRef]
- Paulert, R.; Ebbinghaus, D.; Urlass, C.; Moerschbacher, B.M. Priming of the oxidative burst in rice and wheat cell cultures by ulvan, a polysaccharide from green macroalgae, and enhanced resistance against powdery mildew in wheat and barley plants. Plant Pathol. 2010, 59, 634–642. [Google Scholar] [CrossRef]
- de Freitas, M.B.; Stadnik, M.J. Ulvan-induced resistance in Arabidopsis thaliana against Alternaria brassicicola requires reactive oxygen species derived from NADPH oxidase. Physiol. Mol. Plant Pathol. 2015, 90, 49–56. [Google Scholar] [CrossRef]
- El Modafar, C.; Elgadda, M.; El Boutachfaiti, R.; Abouraicha, E.; Zehhar, N.; Petit, E.; El Alaoui-Talibi, Z.; Courtois, B.; Courtois, J. Induction of natural defence accompanied by salicylic acid-dependant systemic acquired resistance in tomato seedlings in response to bioelicitors isolated from green algae. Sci. Hortic. 2012, 138, 55–63. [Google Scholar] [CrossRef]
- Hidangmayum, A.; Dwivedi, P.; Katiyar, D.; Hemantaranjan, A. Application of chitosan on plant responses with special reference to abiotic stress. Physiol. Mol. Biol. Plants 2019, 25, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Ajmera, P.; Kalani, S.; Sharma, L. Parsley-benefits & side effects on health. Int. J. Physiol. Nutr. Phys. Educ. 2019, 4, 1236–1242. [Google Scholar]
- Makri, O.; Kintzios, S. Ocimum sp. (Basil): Botany, Cultivation, Pharmaceutical Properties, and Biotechnology. J. Herbs. Spices Med. Plants 2008, 13, 123–150. [Google Scholar] [CrossRef]
- Marthe, F.; Bruchmüller, T.; Börner, A.; Lohwasser, U. Variability in parsley (Petroselinum crispum [Mill.] Nyman) for reaction to Septoria petroselini Desm., Plasmopara petroselini Săvul. et O. Săvul. and Erysiphe heraclei DC. ex Saint-Aman causing Septoria blight, downy mildew an. Genet. Resour. Crop Evol. 2013, 60, 1007–1020. [Google Scholar] [CrossRef]
- Topolovec-Pintarić, S.; Martinko, K. Downy Mildew of Basil: A New Destructive Disease Worldwide. In Plant Diseases-Current Threats and Management Trends; IntechOpen: London, UK, 2020. [Google Scholar]
- Lahaye, M.; Inizan, F.; Vigoureux, J. NMR analysis of the chemical structure of ulvan and of ulvan-boron complex formation. Carbohydr. Polym. 1998, 36, 239–249. [Google Scholar] [CrossRef]
- de Carvalho, M.M.; de Freitas, R.A.; Ducatti, D.R.B.; Ferreira, L.G.; Gonçalves, A.G.; Colodi, F.G.; Mazepa, E.; Aranha, E.M.; Noseda, M.D.; Duarte, M.E.R. Modification of ulvans via periodate-chlorite oxidation: Chemical characterization and anticoagulant activity. Carbohydr. Polym. 2018, 197, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Robic, A.; Bertrand, D.; Sassi, J.-F.; Lerat, Y.; Lahaye, M. Determination of the chemical composition of ulvan, a cell wall polysaccharide from Ulva spp. (Ulvales, Chlorophyta) by FT-IR and chemometrics. J. Appl. Phycol. 2009, 21, 451–456. [Google Scholar] [CrossRef]
- Masanetz, C.; Grosch, W. Key odorants of parsley leaves (Petroselinum crispum [Mill.] Nym. ssp. crispum) by Odour-activity values. Flavour Fragr. J. 1998, 13, 115–124. [Google Scholar] [CrossRef]
- Harper, C.B.; Johnson, A.J.; Meyerson, E.; Savas, T.L.; Miikkulainen, R.; de la Parra, J.; Savas, T.L.; Miikkulainen, R.; Harper, C.B. Flavor-cyber-agriculture: Optimization of plant metabolites in an open-source control environment through surrogate modeling. PLoS ONE 2019, 14, e0213918. [Google Scholar] [CrossRef] [Green Version]
- Shukla, P.S.; Borza, T.; Critchley, A.T.; Prithiviraj, B. Seaweed-Based Compounds and Products for Sustainable Protection against Plant Pathogens. Mar. Drugs 2021, 19, 59. [Google Scholar] [CrossRef]
- Abouraïcha, E.; El Alaoui-Talibi, Z.; El Boutachfaiti, R.; Petit, E.; Courtois, B.; Courtois, J.; El Modafar, C. Induction of natural defense and protection against Penicillium expansum and Botrytis cinerea in apple fruit in response to bioelicitors isolated from green algae. Sci. Hortic. 2015, 181, 121–128. [Google Scholar] [CrossRef]
- Arif, Y.; Sami, F.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salicylic acid in relation to other phytohormones in plant: A study towards physiology and signal transduction under challenging environment. Environ. Exp. Bot. 2020, 175, 104040. [Google Scholar] [CrossRef]
- Robert-Seilaniantz, A.; Grant, M.; Jones, J.D.G. Hormone Crosstalk in Plant Disease and Defense: More Than Just Jasmonate-Salicylate Antagonism. Annu. Rev. Phytopathol. 2011, 49, 317–343. [Google Scholar] [CrossRef]
- Thulke, O.; Conrath, U. Salicylic acid has a dual role in the activation of defence-related genes in parsley. Plant J. 1998, 14, 35–42. [Google Scholar] [CrossRef]
- Gamir, J.; Sánchez-Bel, P.; Flors, V. Molecular and physiological stages of priming: How plants prepare for environmental challenges. Plant Cell Rep. 2014, 33, 1935–1949. [Google Scholar] [CrossRef] [PubMed]
- Prodhan, M.Y.; Munemasa, S.; Nahar, M.N.-E.-N.; Nakamura, Y.; Murata, Y. Guard Cell Salicylic Acid Signaling Is Integrated into Abscisic Acid Signaling via the Ca2+/CPK-Dependent Pathway. Plant Physiol. 2018, 178, 441–450. [Google Scholar] [CrossRef] [Green Version]
- Koo, Y.M.; Heo, A.Y.; Choi, H.W. Salicylic acid as a safe plant protector and growth regulator. Plant Pathol. J. 2020, 36, 1–10. [Google Scholar] [CrossRef]
- Bittelli, M.; Flury, M.; Campbell, G.S.; Nichols, E.J. Reduction of transpiration through foliar application of chitosan. Agric. For. Meteorol. 2001, 107, 167–175. [Google Scholar] [CrossRef]
- Iriti, M.; Picchi, V.; Rossoni, M.; Gomarasca, S.; Ludwig, N.; Gargano, M.; Faoro, F. Chitosan antitranspirant activity is due to abscisic acid-dependent stomatal closure. Environ. Exp. Bot. 2009, 66, 493–500. [Google Scholar] [CrossRef]
- Jung, H.W.; Tschaplinski, T.J.; Wang, L.; Glazebrook, J.; Greenberg, J.T. Priming in Systemic Plant Immunity. Science 2009, 324, 89–91. [Google Scholar] [CrossRef]
- Farouk, S.; Amany, A. Improving growth and yield of cowpea by foliar application of chitosan under water stress. Egypt. J. Biol. 2012, 14. [Google Scholar] [CrossRef] [Green Version]
- Deschamps, C.; Raskin, I.; Simon, J.E. Regulation of Essential Oil Accumulation in Basil (Ocimum basilicum L.) in Response to Elicitation. Int. J. Plant Sci. 2008, 169, 981–986. [Google Scholar] [CrossRef]
- Kim, H.-J.; Chen, F.; Wang, X.; Rajapakse, N.C. Effect of Chitosan on the Biological Properties of Sweet Basil (Ocimum basilicum L.). J. Agric. Food Chem. 2005, 53, 3696–3701. [Google Scholar] [CrossRef]
- Pirbalouti, A.G.; Malekpoor, F.; Salimi, A.; Golparvar, A.; Hamedi, B. Effects of Foliar of the Application Chitosan and Reduced Irrigation on Essential Oil Yield, Total Phenol Content and Antioxidant Activity of Extracts from Green and Purple Basil. Acta Sci. Pol. Hortorum Cultus 2017, 16, 177–186. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Filisetti-Cozzi, T.M.C.C.; Carpita, N.C. Measurement of uronic acids without interference from neutral sugars. Anal. Biochem. 1991, 197, 157–162. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Dodgson, K.; Price, R. A note on the determination of the ester sulphate content of sulphated polysaccharides. Biochem. J. 1962, 84, 106–110. [Google Scholar] [CrossRef] [Green Version]
- Ascêncio, S.D.; Orsato, A.; França, R.A.; Duarte, M.E.R.; Noseda, M.D. Complete 1H and 13C NMR assignment of digeneaside, a low-molecular-mass carbohydrate produced by red seaweeds. Carbohydr. Res. 2006, 341, 677–682. [Google Scholar] [CrossRef]
- Vander, P.; Vårum, K.M.; Domard, A.; Eddine El Gueddari, N.; Moerschbacher, B.M. Comparison of the Ability of Partially N-Acetylated Chitosans and Chitooligosaccharides to Elicit Resistance Reactions in Wheat Leaves. Plant Physiol. 1998, 118, 1353–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Pistelli, L.; D’Angiolillo, F.; Morelli, E.; Basso, B.; Rosellini, I.; Posarelli, M.; Barbafieri, M. Response of spontaneous plants from an ex-mining site of Elba island (Tuscany, Italy) to metal(loid) contamination. Environ. Sci. Pollut. Res. 2017, 24, 7809–7820. [Google Scholar] [CrossRef]
- Scartazza, A.; Picciarelli, P.; Mariotti, L.; Curadi, M.; Barsanti, L.; Gualtieri, P. The role of Euglena gracilis paramylon in modulating xylem hormone levels, photosynthesis and water-use efficiency in Solanum lycopersicum L. Physiol. Plant. 2017, 161, 486–501. [Google Scholar] [CrossRef]
- Giuliani, C.; Ascrizzi, R.; Lupi, D.; Tassera, G.; Santagostini, L.; Giovanetti, M.; Flamini, G.; Fico, G. Salvia verticillata: Linking glandular trichomes, volatiles and pollinators. Phytochemistry 2018, 155, 53–60. [Google Scholar] [CrossRef]
- National Institute of Standards and Technology. NIST/EPA/NIH Mass Spectral Library; The NIST Mass Spectrometry Data Center: Gaithersburg, MD, USA, 2014.
- Adams, R.P.R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy; Carol, S., Ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; ISBN 1932633219. [Google Scholar]
- Choi, Y.H.; Kim, H.K.; Hazekamp, A.; Erkelens, C.; Lefeber, A.W.M.; Verpoorte, R. Metabolomic Differentiation of Cannabis sativa Cultivars Using 1H NMR Spectroscopy and Principal Component Analysis. J. Nat. Prod. 2004, 67, 953–957. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Basil | Parsley | |||||
|---|---|---|---|---|---|---|
| Water | Chitosan 100 μg/mL | UIE 1 mg/mL | Water | Chitosan 100 μg/mL | UIE 1 mg/mL | |
| Chlorophyll a (Chla) µg/g FW | 973.37 ± 23.55 a | 898.2 ± 21.74 ab | 857.78 ± 16.71 b | 1112.64 ± 247.03 a | 1424.95 ± 26.83 a | 1294.21 ± 140.59 a |
| Chlorophyll b (Chlb) µg/g FW | 248.86 ± 6.41 a | 229.94 ± 5.56 a | 228.27 ± 2.68 a | 358.75 ± 66.20 a | 433.26 ± 8.97 a | 398.15 ± 41.08 a |
| Total Chlorophylls (TChl) µg/g FW | 1222.23 ± 29.92 a | 1128.26 ± 27.30 ab | 1086.05 ± 19.28 b | 1471.39 ± 312.97 a | 1858.21 ± 35.14 a | 1692.35 ± 181.66 a |
| Ratio Chl a/Chl b | 3.91 a | 3.91 a | 3.76 b | 3.06 a | 3.27 a | 3.25 a |
| Total Carotenoids (TCar) µg/g FW | 248.37 ± 5.46 a | 230.1 ± 6.41 ab | 219.86 ± 4.67 b | 269.71 ± 63.79 a | 342.4 ± 18.13 a | 337.22 ± 32.57 a |
| Ratio TCar/TChl | 0.20 a | 0.20 a | 0.20a | 0.18 a | 0.18 a | 0.20 a |
| Total polyphenols (TP) mg GAE/g FW | 2.63 ± 0.25 a | 2.67 ± 0.14 a | 2.61 ± 0.79 a | 4.52 ± 0.34 a | 5.52 ± 0.24 a | 5.49 ± 0.26 a |
| DPPH assay (IC50 µg/mL) | 1.56 ± 0.29 a | 1.24 ± 0.04 a | 1.13 ± 0.14 a | 10.78± 1.74 a | 6.88 ± 0.41 a | 8.47 ± 0.99 a |
| Basil | Parsley | |||||
|---|---|---|---|---|---|---|
| Water | Chitosan 100 μg/mL | UIE 1 mg/mL | Water | Chitosan 100 μg/mL | UIE 1 mg/mL | |
| Salicylic Acid (SA) ng/g FW | 45.30± 2.68 c | 253.04 ± 14.72 a | 149.68 ± 17.72 b | 225.06 ± 8.40 b | 204.80 ± 5.50 b | 977.98 ± 12.19 a |
| Salicylic Acid Glucoside (SAG) ng/g FW | 1.39 ± 0.18 c | 33.81 ± 2.90 b | 236.19 ± 6.93 a | 113.71 ± 2.71 b | 123.72 ± 8.21 b | 1643.57 ± 42.23 a |
| Jasmonic Acid (JA) ng/g FW | 182.39 ± 5.83 b | 135.27 ± 12.12 c | 350.58 ± 18.12 a | 84.06 ± 3.75 b | 302.31 ± 2.89 a | 65.87 ± 3.37 c |
| Abscisic Acid (ABA) ng/g FW | 71.28 ± 1.36 b | 89.66 ± 2.37 b | 135.54 ± 15.07 a | 62.38 ± 4.18 c | 93.81 ± 9.35 b | 152.42 ± 3.47 a |
| Azelaic Acid (AZA) ng/g FW | 77.19 ± 4.00 b | 101.30 ± 10.19 a | 73.49 ± 5.25 b | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paulert, R.; Ascrizzi, R.; Malatesta, S.; Berni, P.; Noseda, M.D.; Mazetto de Carvalho, M.; Marchioni, I.; Pistelli, L.; Rabello Duarte, M.E.; Mariotti, L.; et al. Ulva intestinalis Extract Acts as Biostimulant and Modulates Metabolites and Hormone Balance in Basil (Ocimum basilicum L.) and Parsley (Petroselinum crispum L.). Plants 2021, 10, 1391. https://doi.org/10.3390/plants10071391
Paulert R, Ascrizzi R, Malatesta S, Berni P, Noseda MD, Mazetto de Carvalho M, Marchioni I, Pistelli L, Rabello Duarte ME, Mariotti L, et al. Ulva intestinalis Extract Acts as Biostimulant and Modulates Metabolites and Hormone Balance in Basil (Ocimum basilicum L.) and Parsley (Petroselinum crispum L.). Plants. 2021; 10(7):1391. https://doi.org/10.3390/plants10071391
Chicago/Turabian StylePaulert, Roberta, Roberta Ascrizzi, Silvia Malatesta, Paolo Berni, Miguel Daniel Noseda, Mariana Mazetto de Carvalho, Ilaria Marchioni, Luisa Pistelli, Maria Eugênia Rabello Duarte, Lorenzo Mariotti, and et al. 2021. "Ulva intestinalis Extract Acts as Biostimulant and Modulates Metabolites and Hormone Balance in Basil (Ocimum basilicum L.) and Parsley (Petroselinum crispum L.)" Plants 10, no. 7: 1391. https://doi.org/10.3390/plants10071391
APA StylePaulert, R., Ascrizzi, R., Malatesta, S., Berni, P., Noseda, M. D., Mazetto de Carvalho, M., Marchioni, I., Pistelli, L., Rabello Duarte, M. E., Mariotti, L., & Pistelli, L. (2021). Ulva intestinalis Extract Acts as Biostimulant and Modulates Metabolites and Hormone Balance in Basil (Ocimum basilicum L.) and Parsley (Petroselinum crispum L.). Plants, 10(7), 1391. https://doi.org/10.3390/plants10071391