
Metabolite and Elastase Activity Changes in Beach Rose (Rosa rugosa) Fruit and Seeds at Various Stages of Ripeness



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
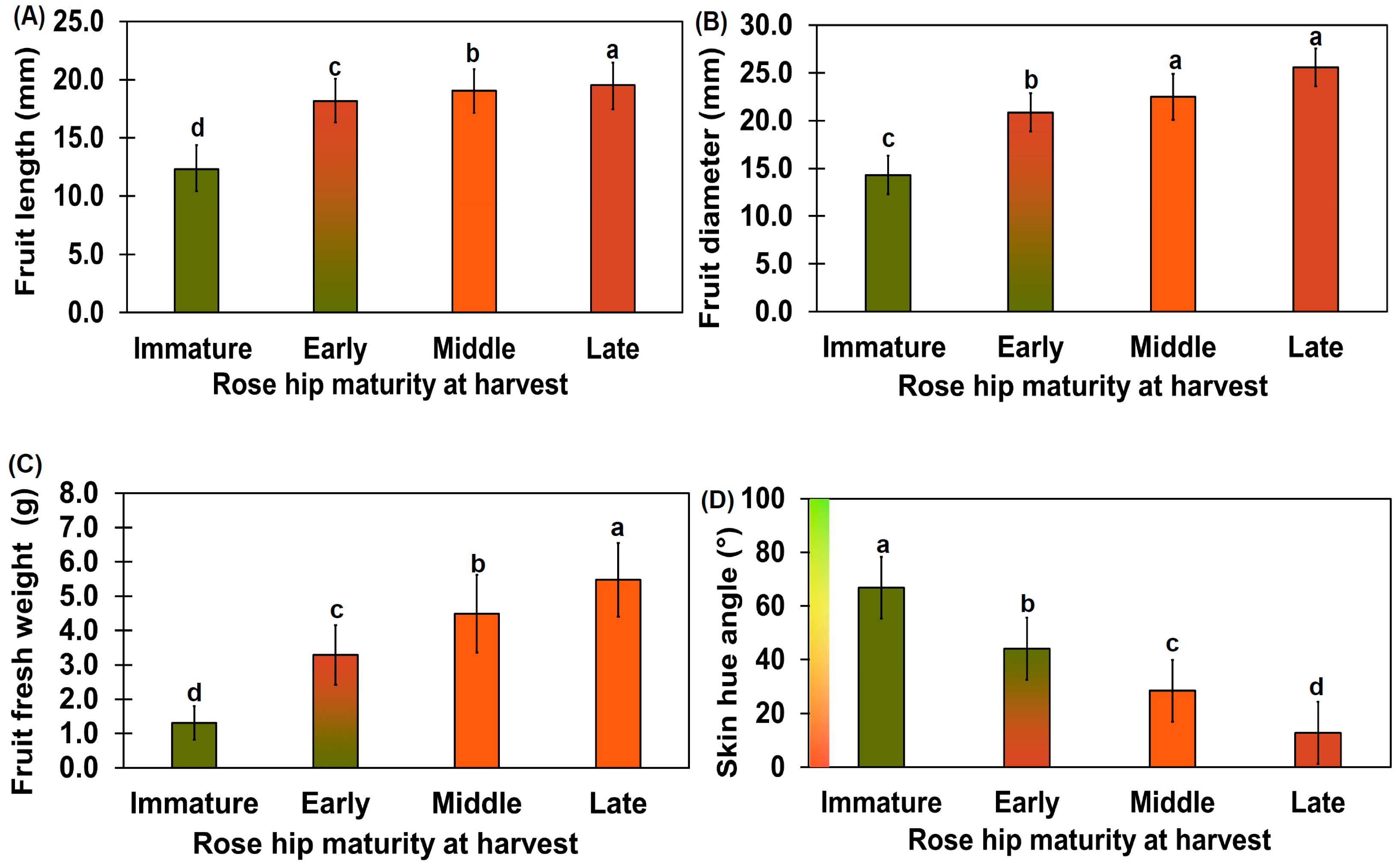
2.1. Characteristics of Each Growth Stage
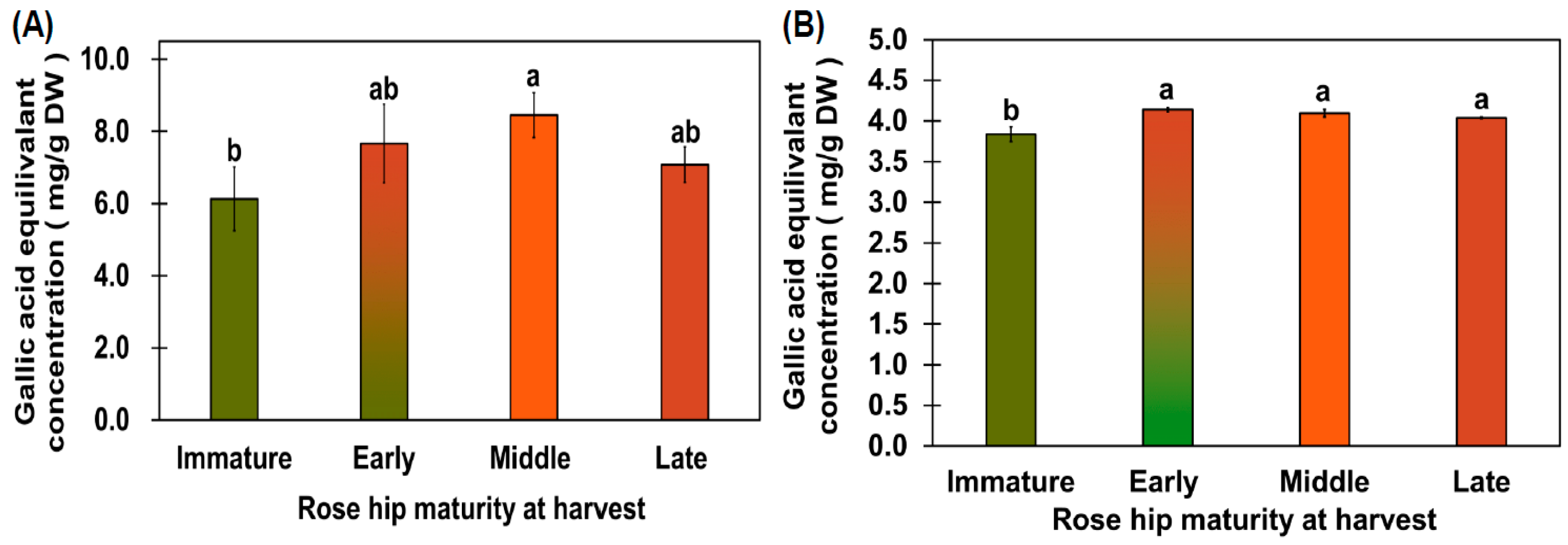
2.2. Total Phenolic Content (TPC) and Antioxidant Capacity of Rose Hip Flesh from Maturity
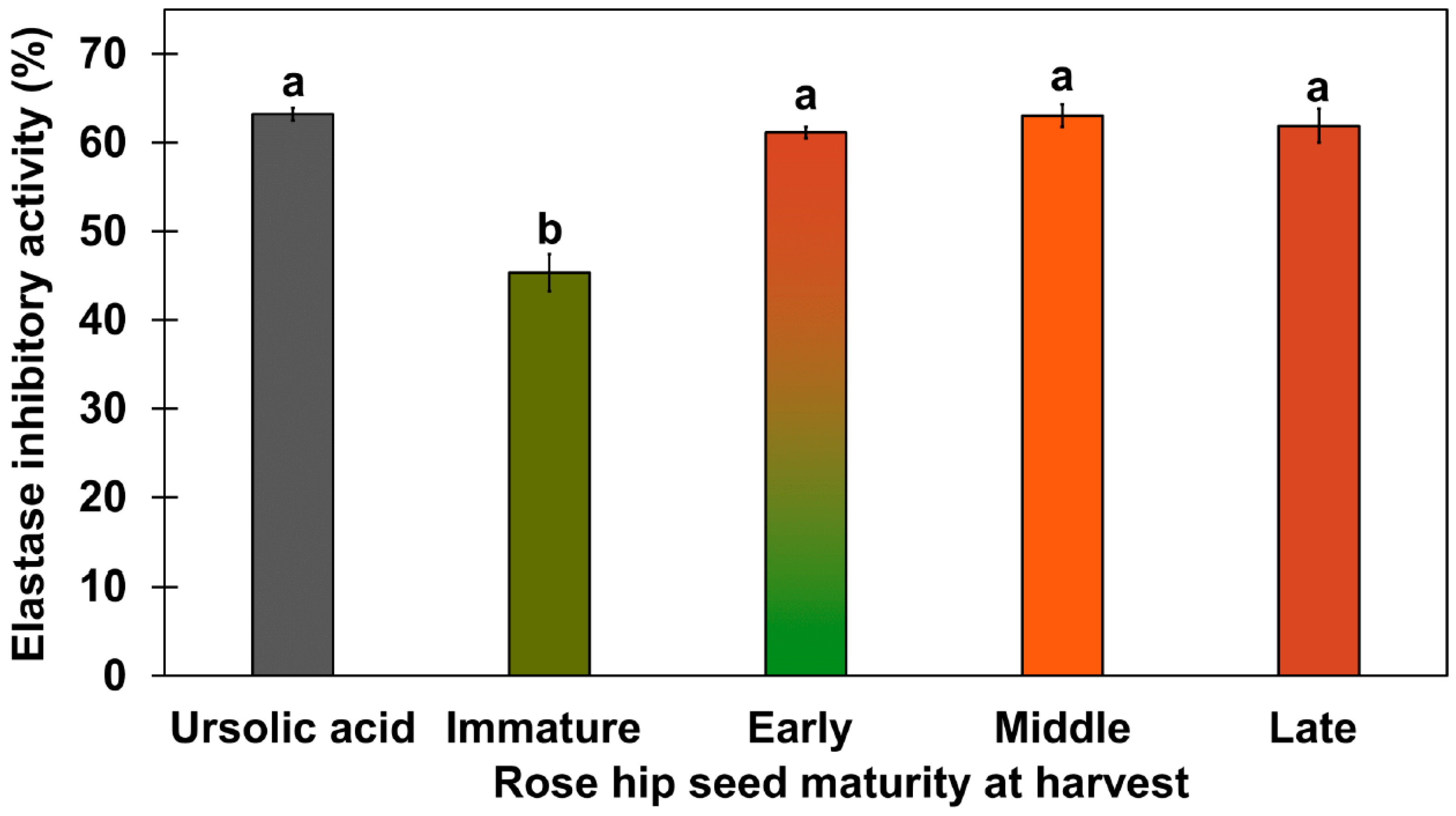
2.3. Elastase Inhibitory Activity
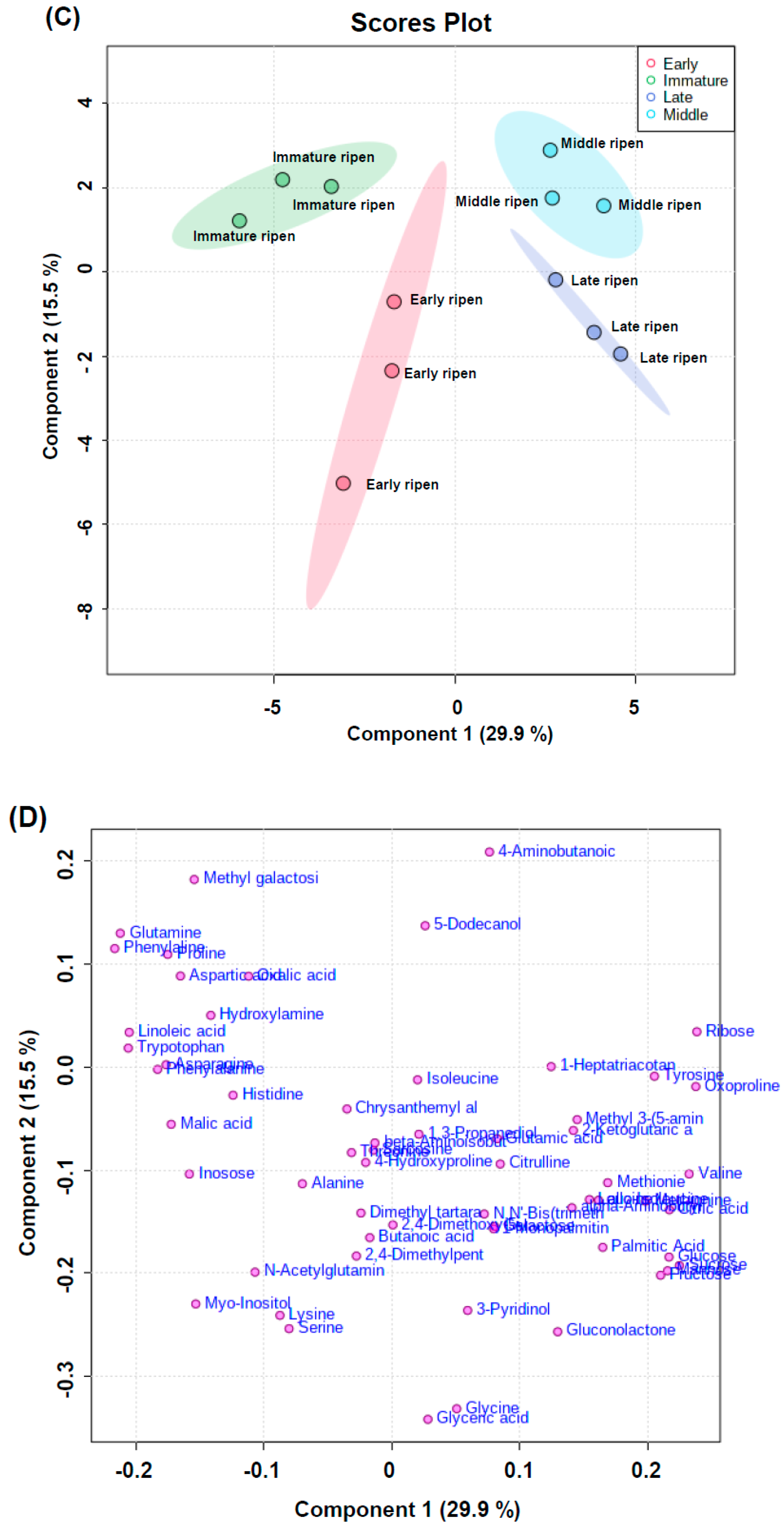
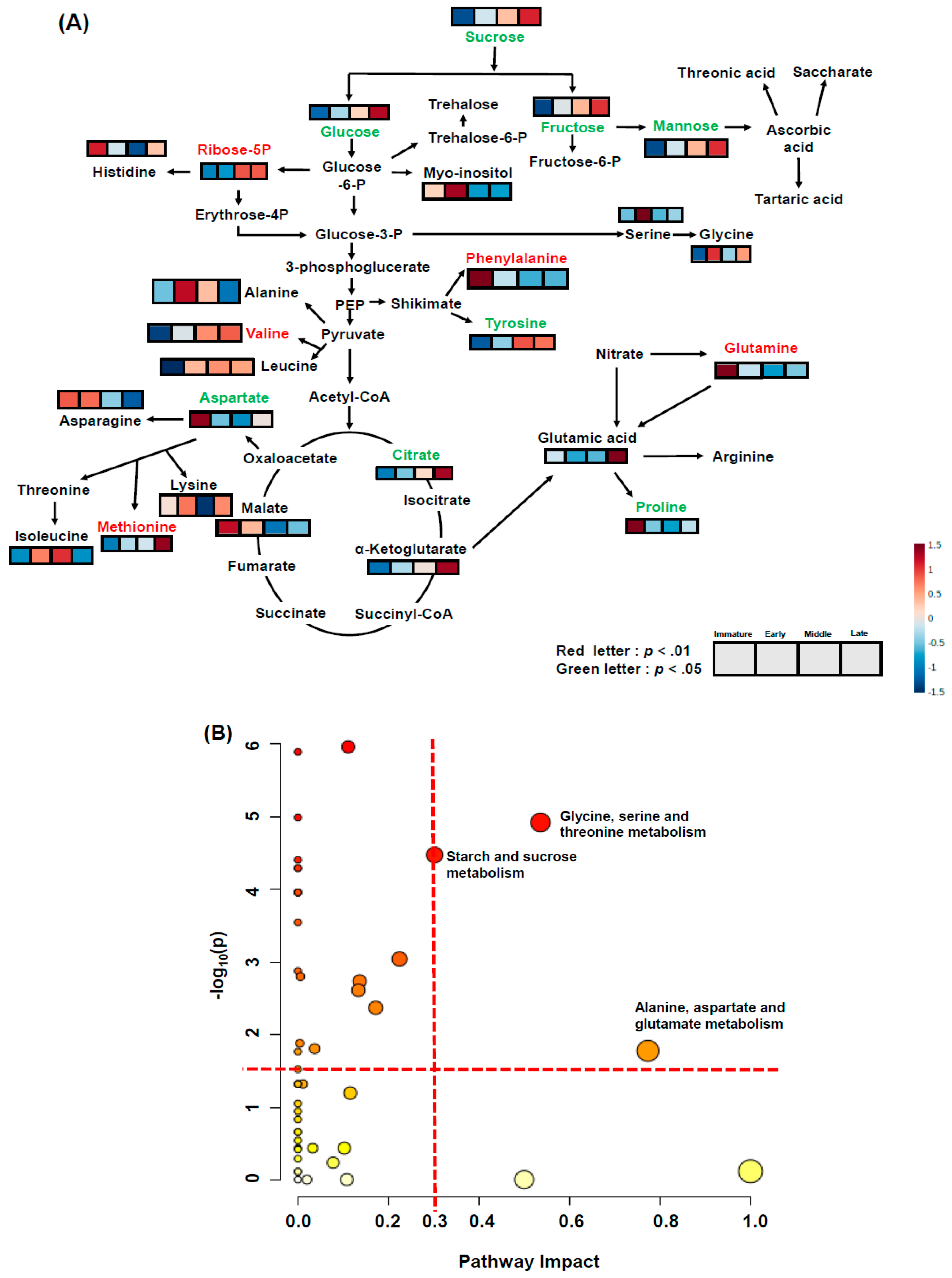
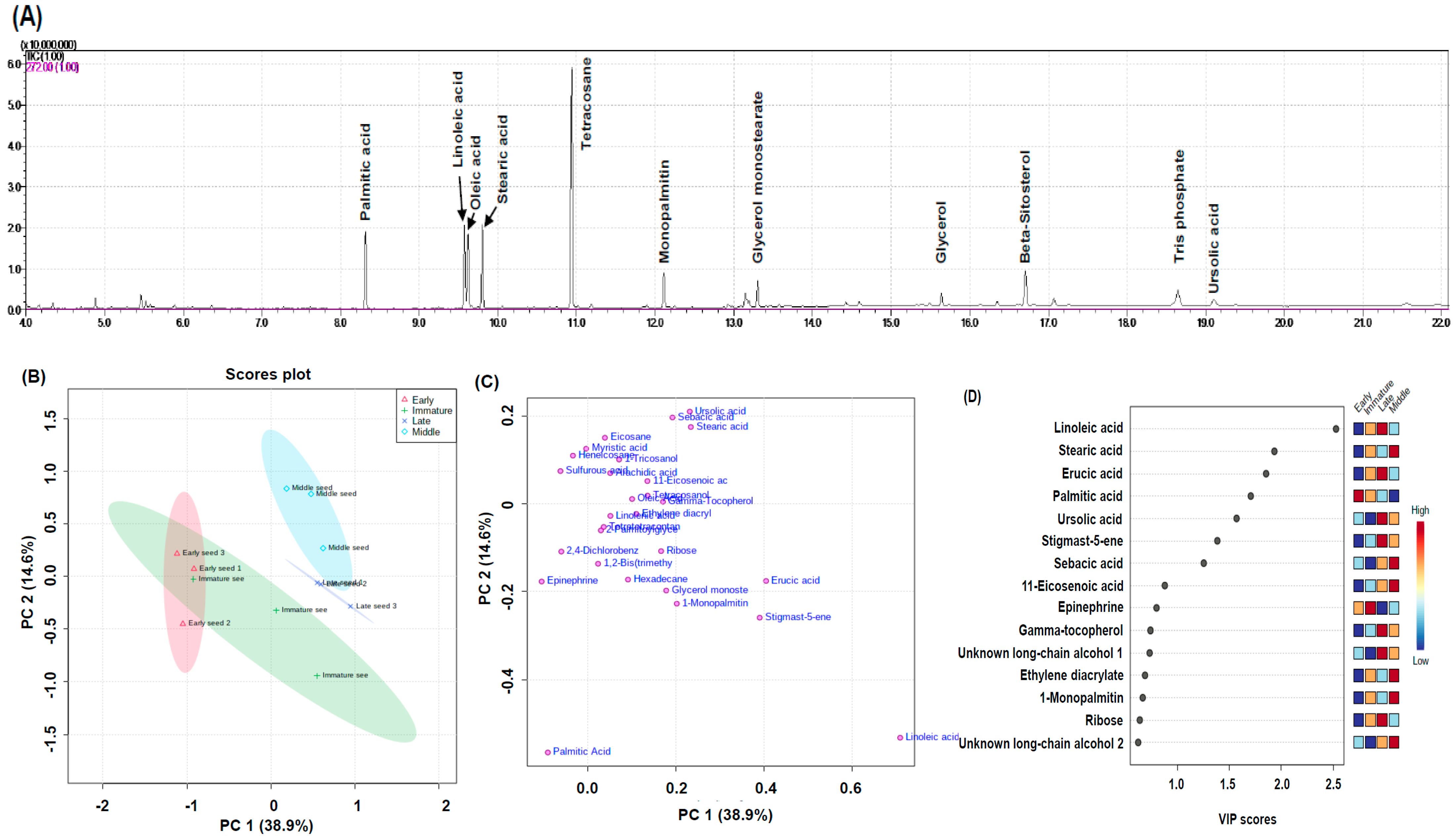
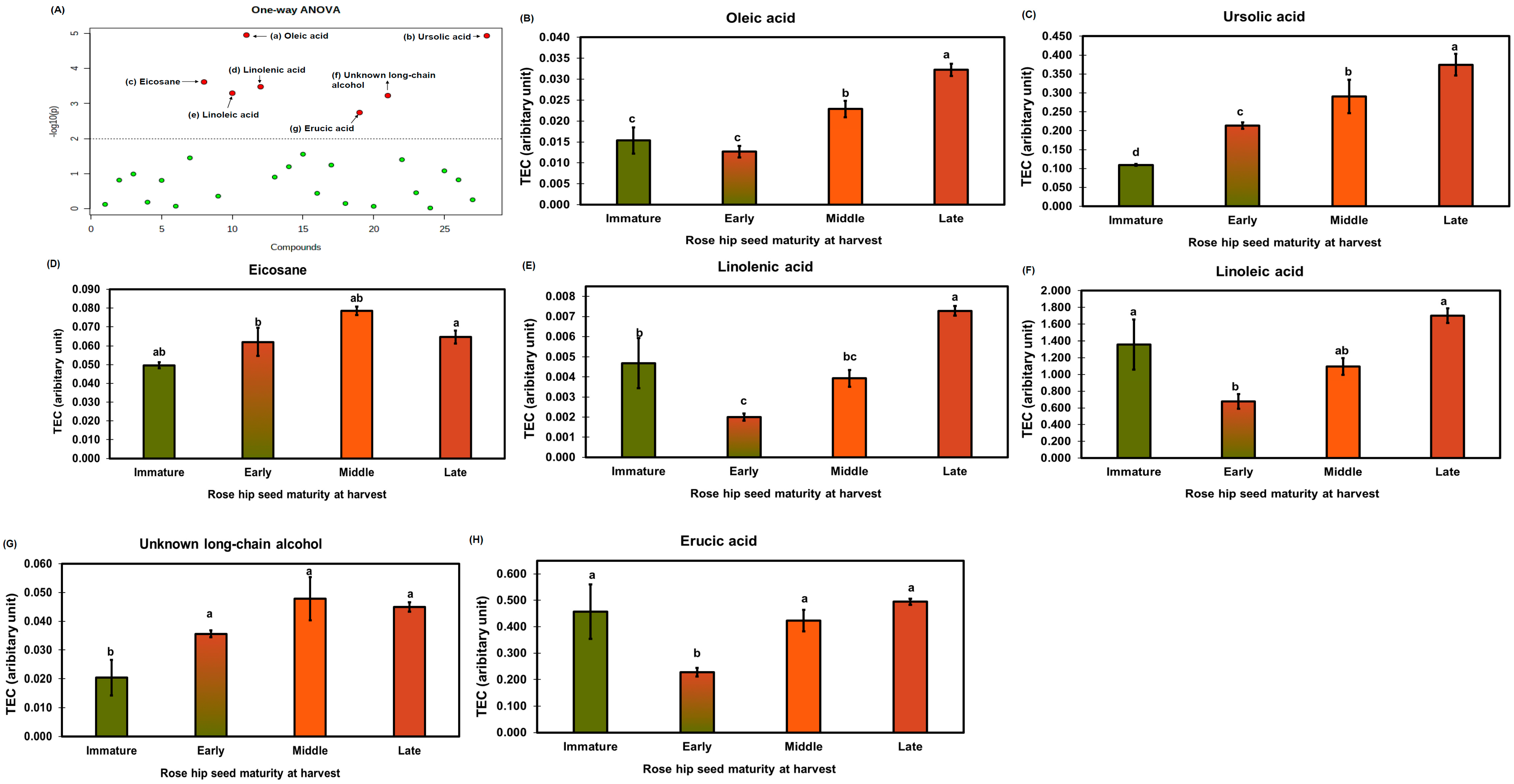
2.4. Water-Soluble Metabolite Profiling for Pathway Analysis
3. Materials and Methods
3.1. Plant Materials and Preparation
3.2. Fruit and Seed Color, Diameter, Weight, and Length Measurements
3.3. TPC Determination
3.4. The 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Antioxidant Capacity
3.5. Elastase Inhibitory Activity
3.6. Water- and Lipid-Soluble Primary Metabolite Profiling Using Gas Chromatography–Mass Spectrometry (GC-MS)
3.7. Amino Acid Quantification
3.8. Data Processing and Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, S.P.; Lahtinen, M.; Sillanpää, M. Green Synthesis and Characterizations of Silver and Gold Nanoparticles Using Leaf Extract of Rosa Rugosa. Colloids Surf. A Physicochem. Eng. Asp. 2010, 364, 34–41. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M.; Byng, J.W. The Number of Known Plants Species in the World and Its Annual Increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Park, B.-J. Isolation of Main Component and Antioxidant Activities on the Stem and Root of Rosa Rugosa. Korean J. Plant Resour. 2008, 21, 402–407. [Google Scholar]
- Gu, D.; Yang, Y.; Bakri, M.; Chen, Q.; Xin, X.; Aisa, H.A. A Lc/Qtof-Ms/Ms Application to Investigate Chemical Compositions in a Fraction with Protein Tyrosine Phosphatase 1b Inhibitory Activity from Rosa Rugosa Flowers. Phytochem. Anal. 2013, 24, 661–670. [Google Scholar] [CrossRef]
- Chrubasik, C.; Duke, R.K.; Chrubasik, S. The evidence for clinical efficacy of rose hip and seed: A systematic review. Phytother. Res. 2006, 20, 1–3. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, F.; Li, R.; Wu, Y.; Liu, S.; Liang, Q. Purification, Characterization, Antioxidant and Moisture-Preserving Activities of Polysaccharides from Rosa Rugosa Petals. Int. J. Biol. Macromol. 2019, 124, 938–945. [Google Scholar] [CrossRef]
- Park, J.C.; Ok, K.D. Phenolic Compounds Isolated from Rosa Rugosa Thunb. In Republic of Korea. Yakhak Hoeji 1993, 37, 365–369. [Google Scholar]
- Demir, F.; Özcan, M. Chemical and technological properties of rose (Rosa canina L.) fruits grown wild in Turkey. J. Food Eng. 2001, 47, 333–336. [Google Scholar] [CrossRef]
- Christensen, R.; Bartels, E.M.; Altman, R.D.; Astrup, A.; Bliddal, H. Does the Hip Powder of Rosa Canina (Rosehip) Reduce Pain in Osteoarthritis Patients?—A Meta-Analysis of Randomized Controlled Trials. Osteoarthr. Cartil. 2008, 16, 965–972. [Google Scholar] [CrossRef] [Green Version]
- Lattanzio, F.; Greco, E.; Carretta, D.; Cervellati, R.; Govoni, P.; Speroni, E. In Vivo Anti-Inflammatory Effect of Rosa canina L. Extract. J. Ethnopharmacol. 2011, 137, 880–885. [Google Scholar] [CrossRef]
- Miraj, S. Phytochemical Composition and in Vitro Pharmacological Activity of Rose Hip (Rosa canina L.). Pharma Chem. 2016, 8, 117–122. [Google Scholar]
- Phetcharat, L.; Wongsuphasawat, K.; Winther, K. The Effectiveness of a Standardized Rose Hip Powder, Containing Seeds and Shells of Rosa Canina, on Cell Longevity, Skin Wrinkles, Moisture, and Elasticity. Clin. Interv. Aging 2015, 10, 1849–1856. [Google Scholar]
- Szentmihályi, K.; Vinkler, P.; Lakatos, B.; Illés, V.; Then, M. Rose Hip (Rosa canina L.) Oil Obtained from Waste Hip Seeds by Different Extraction Methods. Bioresour. Technol. 2002, 82, 195–201. [Google Scholar] [CrossRef]
- Matthaus, B.; Ozcan, M.M. Fatty Acid, Tocopherol and Squalene Contents of Rosaceae Seed Oils. Bot. Stud. 2014, 55, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Daamen, W.F.; Veerkamp, J.H.; van Hest, J.C.M.; van Kuppevelt, T.H. Elastin as a Biomaterial for Tissue Engineering. Biomaterials 2007, 28, 4378–4398. [Google Scholar] [CrossRef] [PubMed]
- Braverman, I.M.; Fonferko, E. Studies in Cutaneous Aging: I. The Elastic Fiber Network. J. Investig. Dermatol. 1982, 78, 434–443. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, H.; Gomi, T.; Shishido, M.; Watanabe, H.; Suenobu, N. Neutrophil Elastase Contributes to Extracellular Matrix Damage Induced by Chronic Low-Dose Uv Irradiation in a Hairless Mouse Photoaging Model. J. Dermatol. Sci. 2010, 60, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Akazaki, S.; Nakagawa, H.; Kazama, H.; Osanai, O.; Kawai, M.; Takema, Y.; Imokawa, G. Age-Related Changes in Skin Wrinkles Assessed by a Novel Three-Dimensional Morphometric Analysis. Br. J. Dermatol. 2002, 147, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Smith, S. Skin Care Products Market Size, Share & Trends Analysis Report, by Product, by Region and Segment Forecasts. Available online: https://www.reportbuyer.com/product/5763749/skin-care-products-market-size-share-and-trends-analysis-report-by-product-by-region-and-segment-forecasts-2019-2025.html?utm_source=PRN#free-sample (accessed on 21 June 2021).
- Ercişli, S.; Ahmet, E. Fruit Characteristics of Native Rose Hip (Rosa Spp.) Selections from the Erzurum Province of Turkey. N. Z. J. Crop Hortic. Sci. 2004, 32, 51–53. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Romero, D.; Castillo, S.; Guillén, F.; Paladine, D.; Zapata, P.J.; Valero, D.; Serrano, M. Rosehip Oil Coating Delays Postharvest Ripening and Maintains Quality of European and Japanese Plum Cultivars. Postharvest Biol. Technol. 2019, 155, 29–36. [Google Scholar] [CrossRef]
- Uggla, M.; Gustavsson, K.-E.; Olsson, M.E.; Nybom, H. Changes in Colour and Sugar Content in Rose Hips (Rosa dumalis L. and R. rubiginosa L.) During Ripening. J. Hortic. Sci. Biotechnol. 2005, 80, 204–208. [Google Scholar] [CrossRef]
- Zhong, L.; Gustavsson, K.-E.; Oredsson, S.; Głąb, B.; Yilmaz, J.L.; Olsson, M.E. Determination of Free and Esterified Carotenoid Composition in Rose Hip Fruit by Hplc-Dad-Apci+-Ms. Food Chem. 2016, 210, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Dolek, U.; Gunes, M.; Genc, N.; Elmastas, M. Total Phenolic Compound and Antioxidant Activity Changes in Rosehip (Rosa sp.) During Ripening. J. Agric. Sci. Technol. 2018, 20, 817–828. [Google Scholar]
- Koczka, N.; Stefanovits-Bányai, É.; Ombódi, A. Total Polyphenol Content and Antioxidant Capacity of Rosehips of Some Rosa Species. Medicines 2018, 5, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, E.-K.; Guo, H.; Choi, J.-K.; Jang, S.-K.; Shin, K.; Cha, Y.-S.; Choi, Y.; Seo, D.-W.; Lee, Y.-B.; Joo, S.-S.; et al. Extraction conditions of white rose petals for the inhibition of enzymes related to skin aging. Lab. Anim. Res. 2015, 31, 148–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thring, T.S.A.; Hili, P.; Naughton, D.P. Anti-Collagenase, Anti-Elastase and Anti-Oxidant Activities of Extracts from 21 Plants. BMC Complement. Altern. Med. 2009, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Simón-Manso, Y.; Lowenthal, M.S.; Kilpatrick, L.E.; Sampson, M.L.; Telu, K.H.; Rudnick, P.A.; Mallard, W.G.; Bearden, D.W.; Schock, T.B.; Tchekhovskoi, D.V.; et al. Metabolite Profiling of a Nist Standard Reference Material for Human Plasma (Srm 1950): Gc-Ms, Lc-Ms, Nmr, and Clinical Laboratory Analyses, Libraries, and Web-Based Resources. Anal. Chem. 2013, 85, 11725–11731. [Google Scholar] [CrossRef]
- Ying, Q.-L.; Rinehart, A.R.; Simon, S.R.; Cheronis, J.C. Inhibition of Human Leucocyte Elastase by Ursolic Acid. Evidence for a Binding Site for Pentacyclic Triterpenes. Biochem. J. 1991, 277, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Castrillo, M.; Kruger, N.J.; Whatley, F.R. Sucrose Metabolism in Mango Fruit during Ripening. Plant Sci. 1992, 84, 45–51. [Google Scholar] [CrossRef]
- Lee, M.Y.; Seo, H.S.; Singh, D.; Lee, S.J.; Lee, C.H. Unraveling Dynamic Metabolomes Underlying Different Maturation Stages of Berries Harvested from Panax Ginseng. J. Ginseng Res. 2020, 44, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Hwang, T.-L.; Su, Y.-C.; Chang, H.-L.; Leu, Y.-L.; Chung, P.-J.; Kuo, L.-M.; Chang, Y.-J. Suppression of Superoxide Anion and Elastase Release by C18 Unsaturated Fatty Acids in Human Neutrophils. J. Lipid Res. 2009, 50, 1395–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cha, B.C.; Lee, E.H. Ingredients of Tyrosinase and Elastase Inhibitory Activity from Calyx of Diospyros Kaki Thunberg. Repub. Korean J. Pharmacogn. 2013, 44, 182–187. [Google Scholar]
- Wenzig, E.M.; Widowitz, U.; Kunert, O.; Chrubasik, S.; Bucar, F.; Knauder, E.; Bauer, R. Phytochemical Composition and in Vitro Pharmacological Activity of Two Rose Hip (Rosa canina L.) Preparations. Phytomedicine 2008, 15, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Ku, K.M.; Choi, J.N.; Kim, J.; Kim, J.K.; Yoo, L.G.; Lee, S.J.; Hong, Y.S.; Lee, C.H. Metabolomics Analysis Reveals the Compositional Differences of Shade Grown Tea (Camellia sinensis L.). J. Agric. Food Chem. 2010, 58, 418–426. [Google Scholar] [CrossRef]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas Chromatography Mass Spectrometry–Based Metabolite Profiling in Plants. Nat. Protoc. 2006, 1, 387. [Google Scholar] [CrossRef]
- Kim, M.J.; Lee, M.Y.; Shon, J.C.; Kwon, Y.S.; Liu, K.-H.; Lee, C.H.; Ku, K.-M. Untargeted and Targeted Metabolomics Analyses of Blackberries–Understanding Postharvest Red Drupelet Disorder. Food Chem. 2019, 300, 125169. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Wishart, D.S.; Xia, J. Using Metaboanalyst 4.0 for Comprehensive and Integrative Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2019, 68, e86. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chae, S.-H.; Lee, Y.-S.; Kim, J.-H.; Han, T.-H.; Ku, K.-M. Metabolite and Elastase Activity Changes in Beach Rose (Rosa rugosa) Fruit and Seeds at Various Stages of Ripeness. Plants 2021, 10, 1283. https://doi.org/10.3390/plants10071283
Chae S-H, Lee Y-S, Kim J-H, Han T-H, Ku K-M. Metabolite and Elastase Activity Changes in Beach Rose (Rosa rugosa) Fruit and Seeds at Various Stages of Ripeness. Plants. 2021; 10(7):1283. https://doi.org/10.3390/plants10071283
Chicago/Turabian StyleChae, Seung-Hun, Young-Sang Lee, Jin-Hee Kim, Tae-Ho Han, and Kang-Mo Ku. 2021. "Metabolite and Elastase Activity Changes in Beach Rose (Rosa rugosa) Fruit and Seeds at Various Stages of Ripeness" Plants 10, no. 7: 1283. https://doi.org/10.3390/plants10071283
APA StyleChae, S.-H., Lee, Y.-S., Kim, J.-H., Han, T.-H., & Ku, K.-M. (2021). Metabolite and Elastase Activity Changes in Beach Rose (Rosa rugosa) Fruit and Seeds at Various Stages of Ripeness. Plants, 10(7), 1283. https://doi.org/10.3390/plants10071283