
Exogenous Stilbenes Improved Tolerance of Arabidopsis thaliana to a Shock of Ultraviolet B Radiation


 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. The Effect of Exogenous Stilbenes, Stilbene Precursors and Octocrylene on A. thaliana Growth
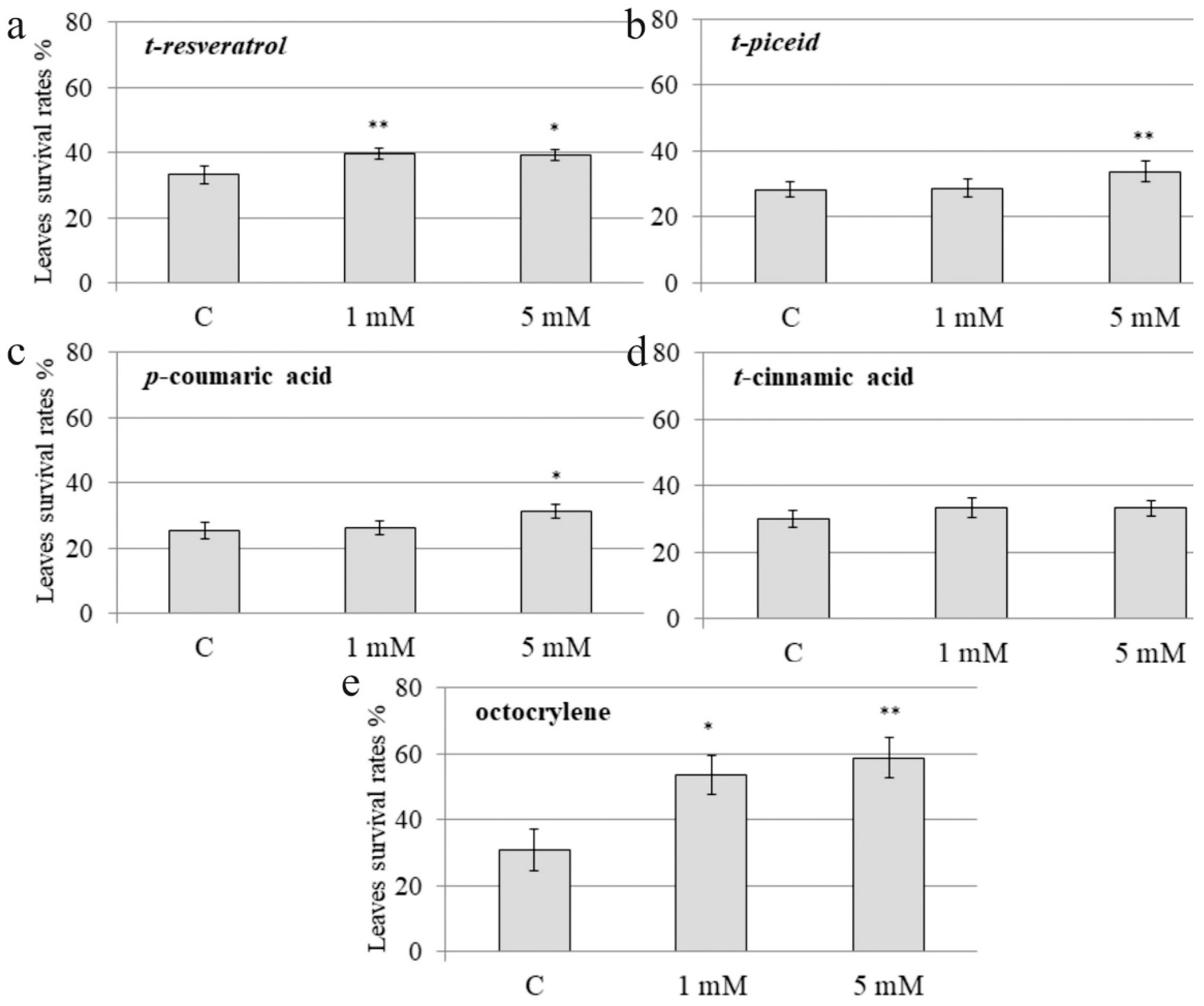
2.2. The Effect of Exogenous Stilbenes, Stilbene Precursors and Octocrylene on A. thaliana Leaf Survival after the UV-B Treatment
2.3. Detection of t-Resveratrol on the A. thaliana Leaves after UV-B Treatment
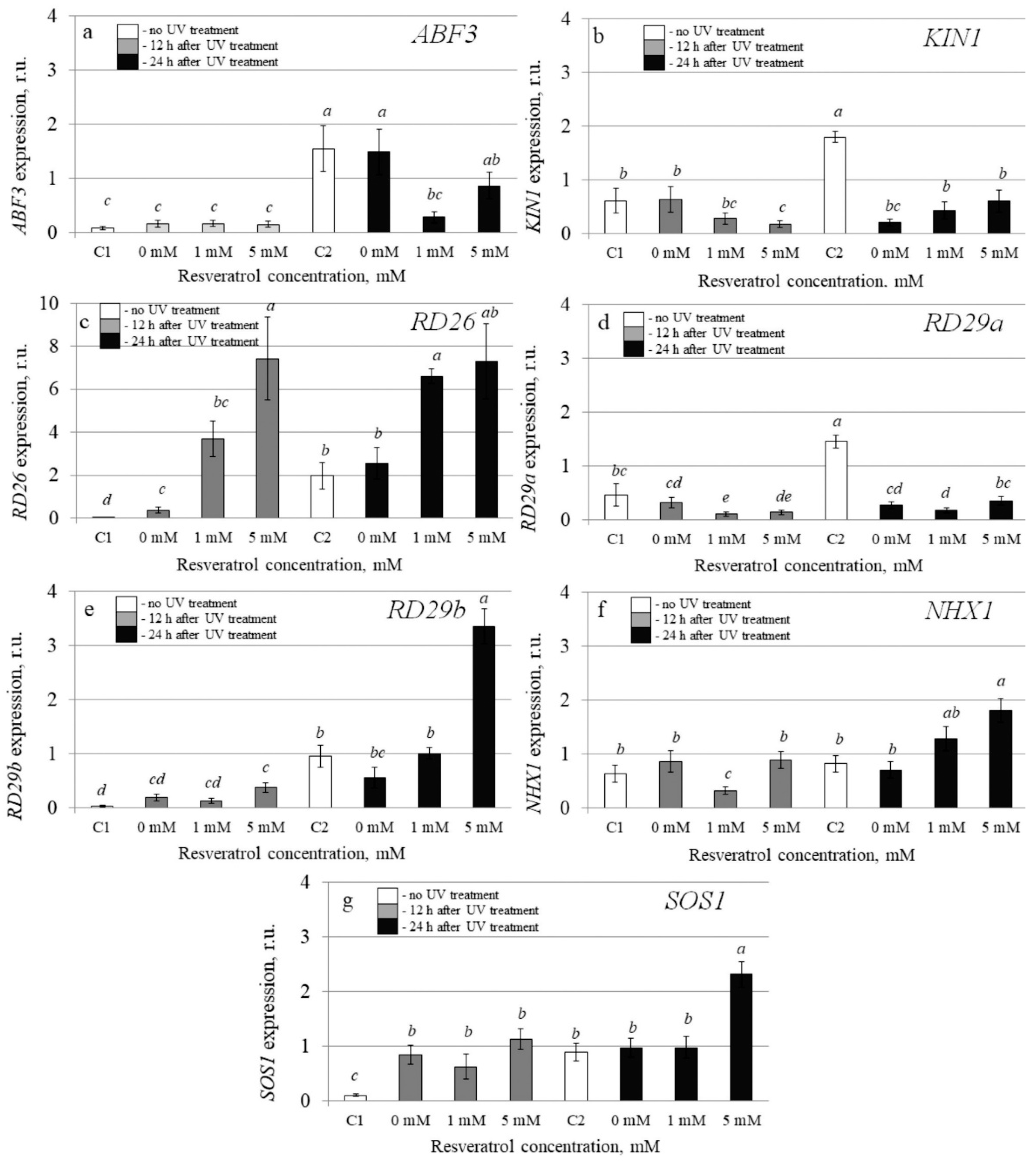
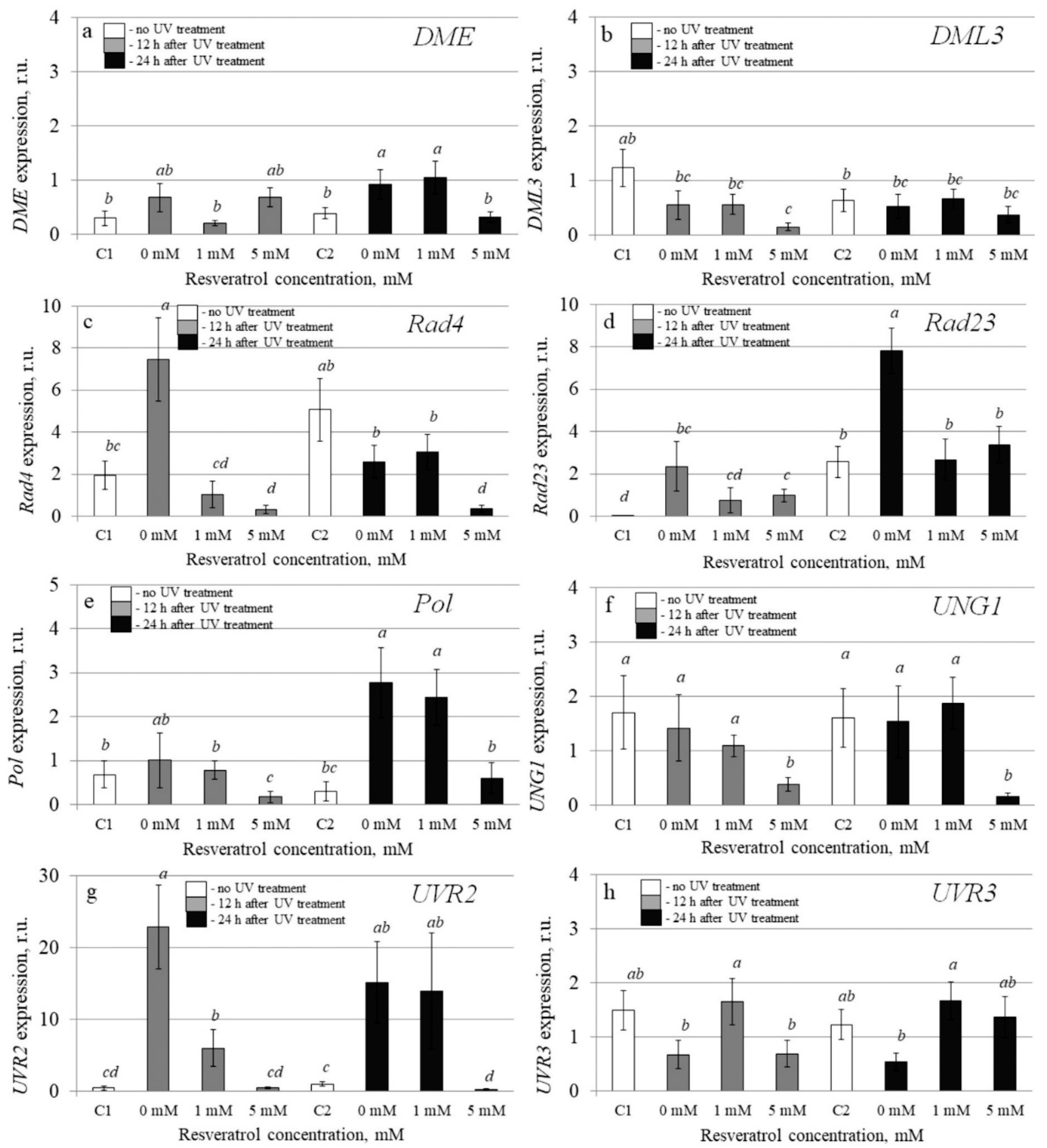
2.4. The Effect of t-Resveratrol and UV-B Treatments on the Expression of Selected Stress-Inducible and DNA Repair Genes
2.5. The Effect of Exogenous p-Coumaric Acid on the Content of Phenolic Secondary Metabolites in A. thaliana
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Foliar Plant Chemical Treatments and UV-B Stress Tolerance Assays
4.3. High Performance Liquid Chromatography with Diode Array Detection (HPLC-DAD)
4.4. Total RNA Isolation and Real-Time Quantitative RT-PCR
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hollósy, F. Effects of ultraviolet radiation on plant cells. Micron 2002, 33, 179–197. [Google Scholar] [CrossRef]
- Kakani, V.G.; Reddy, K.R.; Zhao, D.; Sailaja, K. Field crop responses to ultraviolet-B radiation: A review. Agric. For. Meteorol. 2003, 120, 191–218. [Google Scholar] [CrossRef]
- Strid, A.; Chow, W.S.; Anderson, J.M. UV-B damage and protection at the molecular level in plants. Photosynth. Res. 1994, 39, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Teramura, A.H.; Sullivan, J.H. Effects of UV-B radiation on photosynthesis and growth of terrestrial plants. Photosynth. Res. 1994, 39, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Zuk-Golaszewska, K.; Upadhyaya, M.K.; Golaszewski, J. The effect of UV-B radiation on plant growth and development. Plant Soil Environ. 2003, 49, 135–140. [Google Scholar] [CrossRef] [Green Version]
- Wargent, J.J.; Jordan, B.R. From ozone depletion to agriculture: Understanding the role of UV radiation in sustainable crop production. New Phytol. 2013, 197, 1058–1076. [Google Scholar] [CrossRef]
- Jansen, M.A.K. Ultraviolet-B radiation effects on plants: Induction of morphogenic responses. Physiol. Plant. 2002, 116, 423–429. [Google Scholar] [CrossRef]
- Lois, R.; Buchanan, B.B. Severe sensitivity to ultraviolet radiation in an Arabidopsis mutant deficient in flavonoid accumulation. Planta 1994, 194, 504–509. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T. Phytochemicals as protectors against ultraviolet radiation: Versatility of effects and mechanisms. Planta Med. 2008, 74, 1548–1559. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, L.B.D.; Leal-Costa, M.V.; Menezes, E.A.; Lopes, V.R.; Muzitano, M.F.; Costa, S.S.; Tavares, E.S. Ultraviolet-B radiation effects on phenolic profile and flavonoid content of Kalanchoe pinnata. J. Photochem. Photobiol. B 2015, 148, 73–81. [Google Scholar] [CrossRef]
- Duan, D.; Halter, D.; Baltenweck, R.; Tisch, C.; Tröster, V.; Kortekamp, A.; Hugueney, P.; Nick, P. Genetic diversity of stilbene metabolism in Vitis sylvestris. J. Exp. Bot. 2015, 66, 3243–3257. [Google Scholar] [CrossRef]
- Ahn, S.Y.; Kim, S.A.; Choi, S.J.; Yun, H.K. Comparison of accumulation of stilbene compounds and stilbene related gene expression in two grape berries irradiated with different light sources. Hortic. Environ. Biotechnol. 2015, 56, 36–43. [Google Scholar] [CrossRef]
- Julkunen-Tiitto, R.; Nenadis, N.; Neugart, S.; Robson, M.; Agati, G.; Vepsäläinen, J.; Zipoli, G.; Nybakken, L.; Winkler, B.; Jansen, M.A.K. Assessing the response of plant flavonoids to UV radiation: An overview of appropriate techniques. Phytochem. Rev. 2015, 14, 273–297. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Ogneva, Z.V.; Suprun, A.R.; Grigorchuk, V.P.; Dubrovina, A.S. Action of ultraviolet-C radiation and p-coumaric acid on stilbene accumulation and expression of stilbene biosynthesis-related genes in the grapevine Vitis amurensis Rupr. Acta Physiol. Plant. 2019, 41, 28. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Di Ferdinando, M.; Ferrini, F.; Pollastri, S.; Tattini, M. Functional roles of flavonoids in photoprotection: New evidence, lessons from the past. Plant Physiol. Biochem. 2013, 72, 35–45. [Google Scholar] [CrossRef]
- Csepregi, K.; Hideg, É. Phenolic compound diversity explored in the context of photo-oxidative stress protection. Phytochem. Anal. 2018, 29, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Landry, L.G.; Chapple, C.C.S.; Last, R.L. Arabidopsis mutants lacking phenolic sunscreens exhibit enhanced ultraviolet-B injury and oxidative damage. Plant Physiol. 1995, 109, 1159–1166. [Google Scholar] [CrossRef] [Green Version]
- Kiselev, K.V. Perspectives for production and application of resveratrol. Appl. Microbiol. Biotechnol. 2011, 90, 417–425. [Google Scholar] [CrossRef]
- El Khawand, T.; Courtois, A.; Valls, J.; Richard, T.; Krisa, S. A review of dietary stilbenes: Sources and bioavailability. Phytochem. Rev. 2018, 17, 1007–1029. [Google Scholar] [CrossRef]
- Chong, J.; Poutaraud, A.; Hugueney, P. Metabolism and roles of stilbenes in plants. Plant Sci. 2009, 177, 143–155. [Google Scholar] [CrossRef]
- Berman, A.Y.; Motechin, R.A.; Wiesenfeld, M.Y.; Holz, M.K. The therapeutic potential of resveratrol: A review of clinical trials. NPJ Precis. Oncol. 2017, 1, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubrovina, A.S.; Kiselev, K.V. Regulation of stilbene biosynthesis in plants. Planta 2017, 346, 597–623. [Google Scholar] [CrossRef] [PubMed]
- Valletta, A.; Iozia, L.M.; Leonelli, F. Impact of environmental factors on stilbene biosynthesis. Plants 2021, 10, 90. [Google Scholar] [CrossRef] [PubMed]
- Cantos, E.; Garcia-Viguera, C.; de Pascual-Teresa, S.; Tomas-Berberan, F.A. Effect of postharvest ultraviolet irradiation on resveratrol and other phenolics of cv. Napoleon table grapes. J. Agric. Food Chem. 2000, 48, 4606–4612. [Google Scholar] [CrossRef]
- Li, X.; Zheng, X.; Yan, S.; Li, S. Effects of salicylic acid (SA), ultraviolet radiation (UV-B and UV-C) on trans-resveratrol inducement in the skin of harvested grape berries. Front. Agric. China 2008, 2, 77–81. [Google Scholar] [CrossRef]
- Sheng, K.; Zheng, H.; Shui, S.S.; Yan, L.; Liu, C.; Zheng, L. Comparison of postharvest UV-B and UV-C treatments on table grape: Changes in phenolic compounds and their transcription of biosynthetic genes during storage. Postharvest Biol. Technol. 2018, 138, 74–81. [Google Scholar] [CrossRef]
- Kong, Q.; Deng, R.; Li, X.; Zeng, Q.; Zhang, X.; Yu, X.; Ren, X. Based on RNA-Seq analysis identification and expression analysis of trans-scripusin. A synthesize-related genes of UV-treatment in postharvest grape fruit. Arch. Biochem. Biophys. 2020, 690, 108471. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, D.; Wang, L.; Jiang, C.; Wang, Y. VqMAPKKK38 is essential for stilbene accumulation in grapevine. Hortic. Res. 2017, 4, 17058. [Google Scholar] [CrossRef] [Green Version]
- Ogneva, Z.V.; Aleynova, O.A.; Suprun, A.R.; Karetin, Y.A.; Dubrovina, A.S.; Kiselev, K.V. Tolerance of Arabidopsis thaliana plants overexpressing grapevine VaSTS1 or VaSTS7 genes to cold, heat, drought, salinity, and ultraviolet irradiation. Biol. Plant. 2021, 65, 111–117. [Google Scholar] [CrossRef]
- Ma, F.; Yao, W.; Wang, L.; Wang, Y. Dynamic translocation of stilbene synthase VpSTS29 from a Chinese wild Vitis species upon UV irradiation. Phytochemistry 2019, 159, 137–147. [Google Scholar] [CrossRef]
- Nagapan, T.S.; Ghazali, A.R.; Basri, D.F.; Lim, W.N. Photoprotective effect of stilbenes and its derivatives against ultraviolet radiation-induced skin disorders. Biomed. Pharmacol. J. 2018, 11, 3. [Google Scholar] [CrossRef]
- Svobodová, A.; Psotová, J.; Walterová, D. Natural phenolics in the prevention of UV-induced skin damage. A review. Biomed. Pap. 2003, 147, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Tyunin, A.P.; Kiselev, K.V. Alternations in VaSTS gene cytosine methylation and t-resveratrol production in response to UV-C irradiation in Vitis amurensis Rupr. cells. Plant Cell Tiss. Organ Cult. 2016, 124, 33–45. [Google Scholar] [CrossRef]
- D’Auria, J.C.; Gershenzon, J. The secondary metabolism of Arabidopsis thaliana: Growing like a weed. Curr. Opin. Plant Biol. 2005, 8, 308–316. [Google Scholar] [CrossRef]
- Kraker, J.W.; Jonathan, G. From amino acid to glucosinolate biosynthesis: Protein sequence changes in the evolution of methylthioalkylmalate synthase in Arabidopsis. Plant Cell 2011, 23, 38–53. [Google Scholar] [CrossRef] [Green Version]
- Kolb, C.A.; Kaser, M.A.; Kopecky, J.; Zotz, G.; Riederer, M.; Pfundel, E.E. Effects of natural intensities of visible and ultraviolet radiation on epidermal ultraviolet screening and photosynthesis in grape leaves. Plant Physiol. 2001, 127, 863–875. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Grigorchuk, V.P.; Ogneva, Z.V.; Suprun, A.R.; Dubrovina, A.S. Stilbene biosynthesis in the needles of spruce Picea jezoensis. Phytochemistry 2016, 131, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Romero-Pérez, A.I.; Ibern-Gómez, M.; Lamuela-Raventós, R.M.; de la Torre-Boronat, M.C. Piceid, the major resveratrol derivative in grape juices. J. Agric. Food Chem. 1999, 47, 1533–1536. [Google Scholar] [CrossRef]
- Chelikani, P.; Fita, I.; Loewen, P.C. Diversity of structures and properties among catalases. Cell. Mol. Life Sci. 2004, 61, 192–208. [Google Scholar] [CrossRef] [PubMed]
- Takshak, S.; Agrawal, S.B. Effect of ultraviolet-B radiation on biomass production, lipid peroxidation, reactive oxygen species, and antioxidants in Withania somnifera. Biol. Plant. 2014, 58, 328–334. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Maruyama, K.; Seki, M.; Hiratsu, K.; Ohme-Takagi, M.; Tran, L.S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A dehydration-induced NAC protein, RD26, is involved in a novel ABA-dependent stress-signaling pathway. Plant J. 2004, 39, 863–876. [Google Scholar] [CrossRef] [PubMed]
- Apse, M.P.; Aharon, G.S.; Snedden, W.A.; Blumwald, E. Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 1999, 20, 1256–1258. [Google Scholar] [CrossRef]
- Liu, P.; Yang, G.D.; Li, H.; Wu, C.A. Overexpression of NHX1s in transgenic Arabidopsis enhances photoprotection capacity in high salinity and drought conditions. Acta Physiol. Plant. 2010, 32, 81–90. [Google Scholar] [CrossRef]
- Msanne, J.; Lin, J.; Stone, J.M.; Awada, T. Characterization of abiotic stress-responsive Arabidopsis thaliana RD29A and RD29B genes and evaluation of transgenes. Planta 2011, 234, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Modarresi, M.; Eyvazi, A. Bioinformatical evaluation of desiccation-responsive rd29A gene in Arabidopsis thaliana. Pak. J. Biol. Sci. 2014, 1, 80–85. [Google Scholar] [CrossRef] [Green Version]
- Leonard, E.; Yan, Y.; Lim, K.H.; Koffas, M.A. Investigation of two distinct flavone synthases for plant-specific flavone biosynthesis in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2005, 71, 8241–8248. [Google Scholar] [CrossRef] [Green Version]
- Shumakova, O.A.; Manyakhin, A.Y.; Kiselev, K.V. Resveratrol content and expression of phenylalanine ammonia-lyase and stilbene synthase genes in cell cultures of Vitis amurensis treated with coumaric acid. Appl. Biochem. Biotechnol. 2011, 165, 1427–1436. [Google Scholar] [CrossRef]
- Aleynova, O.A.; Grigorchuk, V.P.; Dubrovina, A.S.; Rybin, V.G.; Kiselev, K.V. Stilbene accumulation in cell cultures of Vitis amurensis Rupr. overexpressing VaSTS1, VaSTS2, and VaSTS7 genes. Plant Cell Tiss. Organ Cult. 2016, 125, 329–339. [Google Scholar] [CrossRef]
- Ryan, K.G.; Swinny, E.E.; Winefield, C.; Markham, K.R. Flavonoids and UV photoprotection in Arabidopsis mutants. Z. Naturforsch. C 2001, 56, 745–754. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Tyunin, A.P.; Ogneva, Z.V.; Dubrovina, A.S. Age-associated alterations in the somatic mutation level in Arabidopsis thaliana. Plant Growth Regul. 2015, 75, 493–501. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Aleynova, O.A.; Grigorchuk, V.P.; Dubrovina, A.S. Stilbene accumulation and expression of stilbene biosynthesis pathway genes in wild grapevine Vitis amurensis Rupr. Planta 2017, 245, 151–159. [Google Scholar] [CrossRef]
- Kiselev, K.V.; Shumakova, O.A.; Manyakhin, A.Y.; Mazeika, A.N. Influence of calcium influx induced by the calcium ionophore, A23187, on resveratrol content and the expression of CDPK and STS genes in the cell cultures of Vitis amurensis. Plant Growth Regul. 2012, 68, 371–381. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Number of Leaves in the Rosette | Fresh weight of the Rosette, FW |
|---|---|---|
| Control, H20 | 13.96 ± 0.21 | 0.22 ± 0.02 |
| DMSO | 13.37 ± 0.42 | 0.22 ± 0.04 |
| 1 mM t-resveratrol | 13.88 ± 0.61 | 0.22 ± 0.04 |
| 5 mM t-resveratrol | 13.19 ± 0.45 | 0.19 ± 0.02 |
| 1 mM t-piceid | 12.65 ± 0.67 | 0.22 ± 0.02 |
| 5 mM t-piceid | 13.31 ± 0.51 | 0.27 ± 0.05 |
| 1 mM p-coumaric acid | 14.33 ± 0.83 | 0.33 ± 0.05 * |
| 5 mM p-coumaric acid | 15.67 ± 1.02 | 0.29 ± 0.04 |
| 1 mM t-cinnamic acid | 13.39 ± 0.45 | 0.23 ± 0.05 |
| 5 mM t-cinnamic acid | 13.41 ± 0.46 | 0.24 ± 0.04 |
| 1 mM octocrylene | 13.27 ± 0.43 | 0.21 ± 0.04 |
| 5 mM octocrylene | 13.16 ± 0.39 | 0.21 ± 0.05 |
| Peak Numbers a | Compounds | Control, before Treatment, mg/g FW | 5 mM p-Coumaric Acid, mg/g FW |
|---|---|---|---|
| 1, 3 | Coumaroyl hexoside (isomer 1 and 2) | 0.0 ± 0.0 | 2.64 ± 0.64 ** |
| 2 | Glucohirsutin | 0.05 ± 0.03 | 0.57 ± 0.20 * |
| 4 | Sinapoyl hexoside | 0.06 ± 0.03 | 0.17 ± 0.06 * |
| 5 | Robinin, kaempferol 3-O-rhamnosyl-glucoside 7-O-rhamnoside | 0.32 ± 0.07 | 0.57 ± 0.13 * |
| 6 | p-coumaric acid | 0.0 ± 0.0 | 10.32 ± 1.45 ** |
| 7 | Kaempferol-3-O-glucosid-7-O-ramnoside | 0.21 ± 0.07 | 5.26 ± 1.12 ** |
| 8 | Sinapoyl malate | 2.29 ± 0.47 | 3.96 ± 0.79 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogneva, Z.V.; Volkonskaia, V.V.; Dubrovina, A.S.; Suprun, A.R.; Aleynova, O.A.; Kiselev, K.V. Exogenous Stilbenes Improved Tolerance of Arabidopsis thaliana to a Shock of Ultraviolet B Radiation. Plants 2021, 10, 1282. https://doi.org/10.3390/plants10071282
Ogneva ZV, Volkonskaia VV, Dubrovina AS, Suprun AR, Aleynova OA, Kiselev KV. Exogenous Stilbenes Improved Tolerance of Arabidopsis thaliana to a Shock of Ultraviolet B Radiation. Plants. 2021; 10(7):1282. https://doi.org/10.3390/plants10071282
Chicago/Turabian StyleOgneva, Zlata V., Vlada V. Volkonskaia, Alexandra S. Dubrovina, Andrey R. Suprun, Olga A. Aleynova, and Konstantin V. Kiselev. 2021. "Exogenous Stilbenes Improved Tolerance of Arabidopsis thaliana to a Shock of Ultraviolet B Radiation" Plants 10, no. 7: 1282. https://doi.org/10.3390/plants10071282
APA StyleOgneva, Z. V., Volkonskaia, V. V., Dubrovina, A. S., Suprun, A. R., Aleynova, O. A., & Kiselev, K. V. (2021). Exogenous Stilbenes Improved Tolerance of Arabidopsis thaliana to a Shock of Ultraviolet B Radiation. Plants, 10(7), 1282. https://doi.org/10.3390/plants10071282