
Induction of Polyploidy and Metabolic Profiling in the Medicinal Herb Wedelia chinensis



Abstract
1. Introduction
2. Results
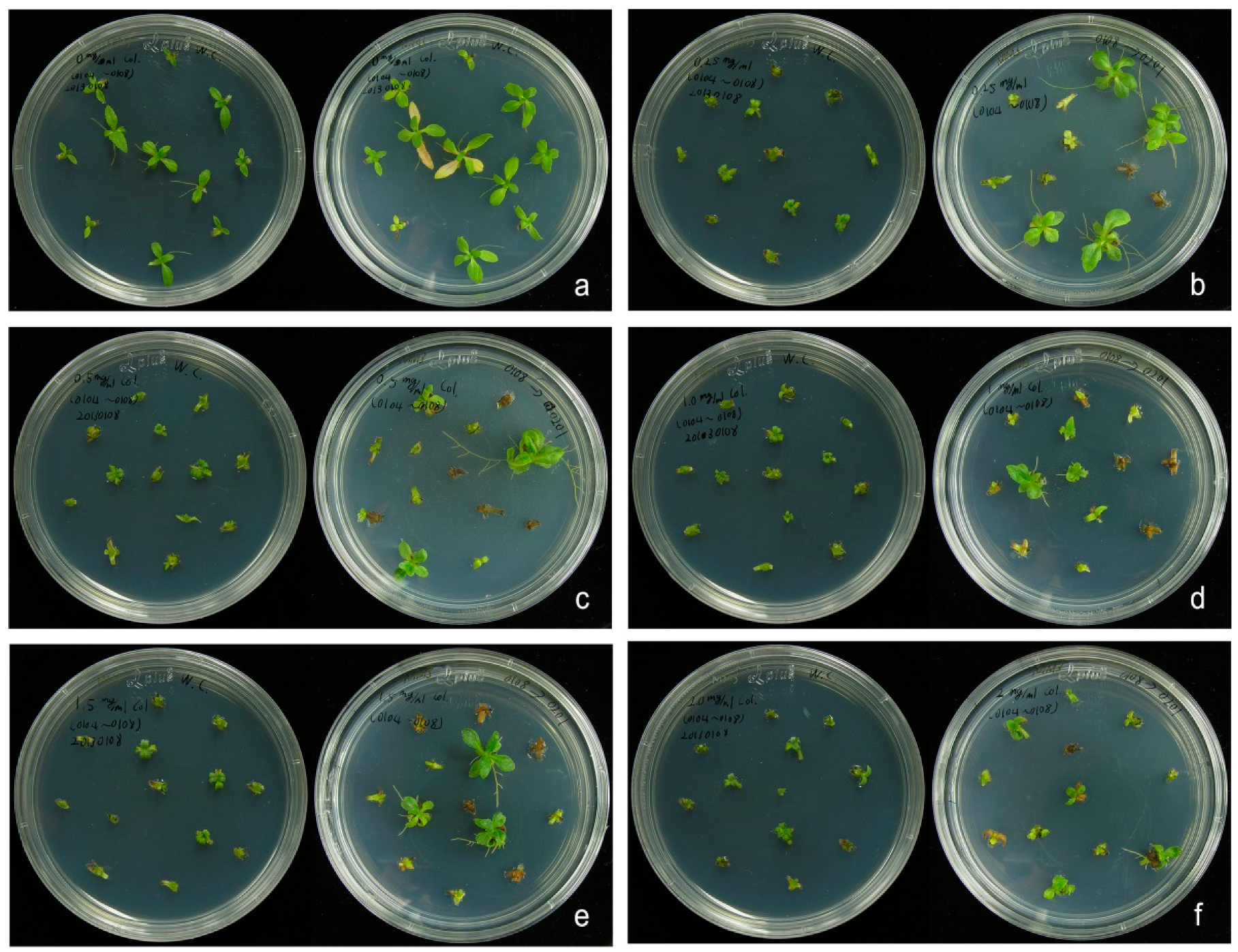
2.1. In Vitro Propagation and Induction of Polyploid Plants
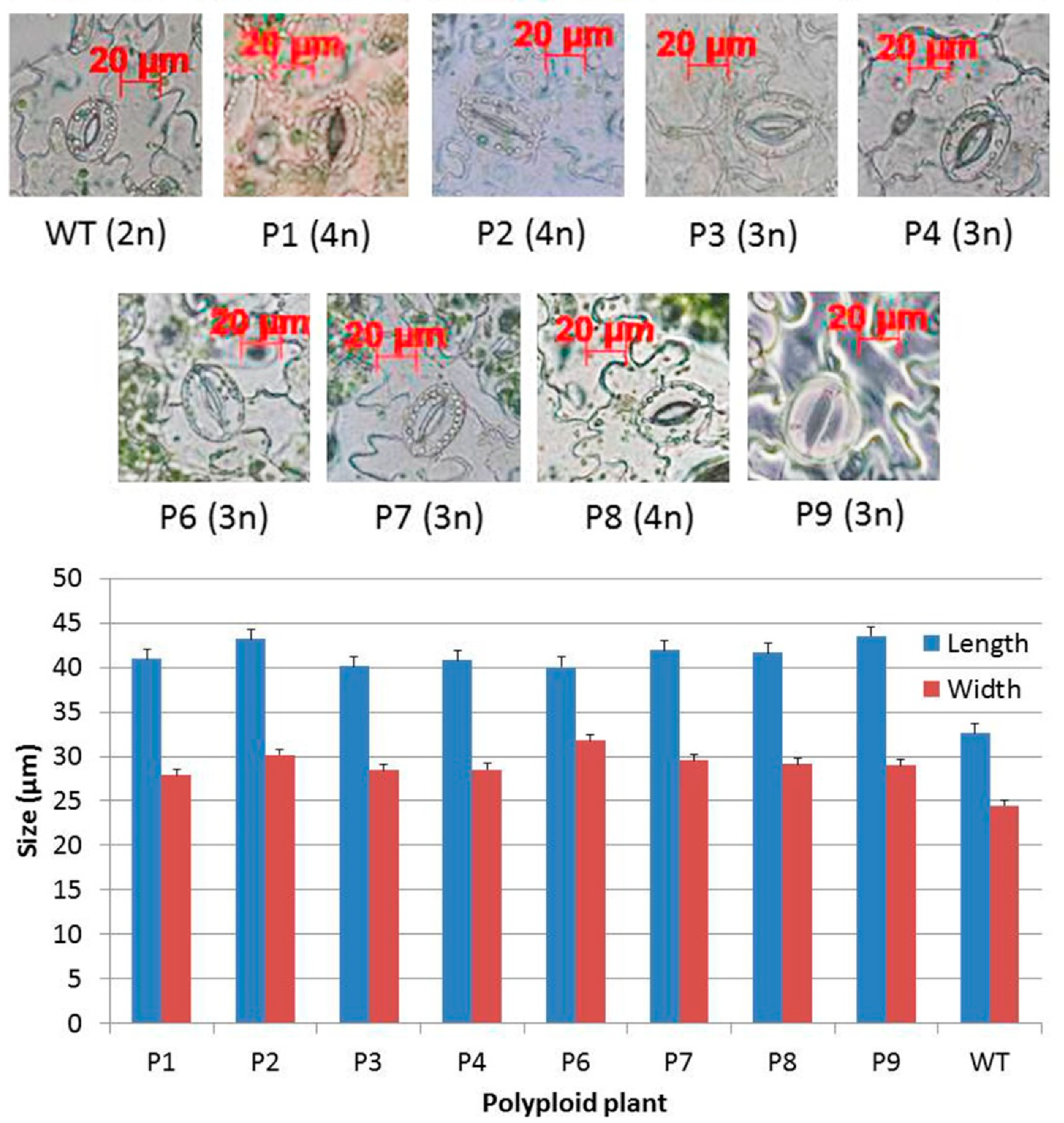
2.2. Characterization of Polyploid Plants
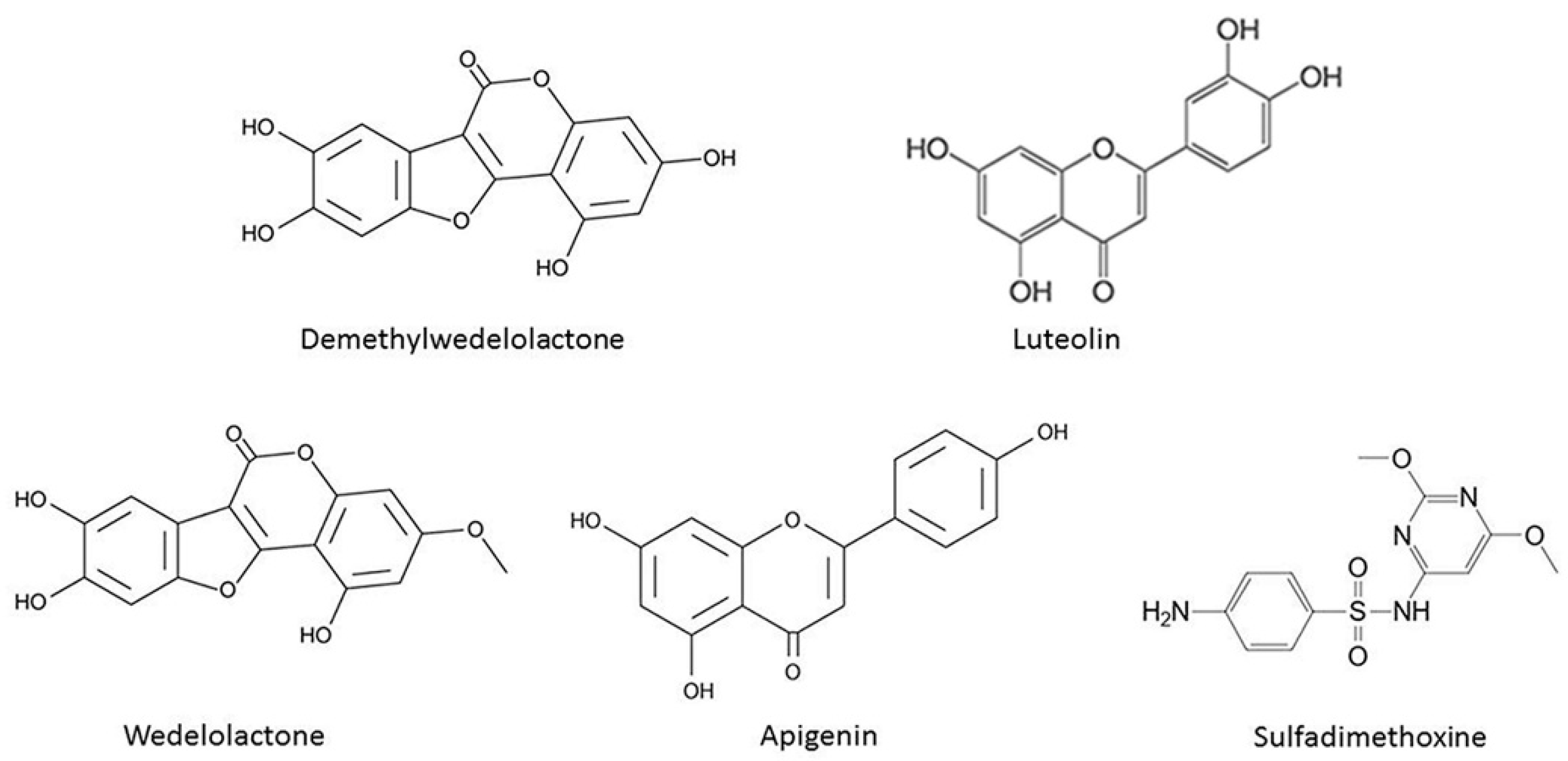
2.3. Identification of Chemical Compounds by UPLC/MS/MS
2.4. Identification and Quantitative Analysis of 4 Metabolites in Wild-Type and Polyploid Plants
3. Discussion
4. Materials and Methods
4.1. Plant Material and Culture Conditions
4.2. Induction of Polyploidy
4.3. Flow Cytometry Analysis
4.4. Measurement of Stomata
4.5. Sample Preparation for UPLC Analysis
4.6. UPLC-DAD-MS/MS Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Lin, S.C.; Lin, C.C.; Lin, Y.H.; Shyuu, S.J. Hepatoprotective effects of Taiwan folk medicine: Wedelia chinensis on three hepatotoxin-induced hepatotoxicity. Am. J. Chin. Med. 1994, 22, 155–168. [Google Scholar] [CrossRef] [PubMed]
- Krishnaraju, A.V.; Tayi, V.N.R.; Sundararaju, D.; Vanisree, M.; Tsay, H.S.; Subbaraju, G.V. Assessment of bioactivity of Indian medicinal plants using brine shrimp (Artemia salina) lethality assay. Intl. J. Appl. Sci. Eng. 2005, 3, 125–134. [Google Scholar]
- Lin, F.M.; Chen, L.R.; Lin, E.H.; Ke, F.C.; Chen, H.Y.; Tsai, M.J.; Hsiao, P.W. Compounds from Wedelia chinensis synergistically suppress androgen activity and growth in prostate cancer cells. Carcinogenesis 2007, 28, 2521–2529. [Google Scholar] [CrossRef]
- Tsai, C.H.; Lin, F.M.; Yang, Y.C.; Lee, M.T.; Cha, T.L.; Wu, G.J.; Hsieh, S.C.; Hsiao, P.W. Herbal extract of Wedelia chinensis attenuates androgen receptor activity and orthotopic growth of prostate cancer in nude mice. Clin. Cancer Res. 2009, 15, 5435–5444. [Google Scholar] [CrossRef] [PubMed]
- Manjamalai, A.; Jiflin, G.J.; Berlin Grace, V.M. Study on the effect of essential oil of Wedelia chinensis (Osbeck) against microbes and inflammation. Asian J. Pharm. Clin. Res. 2012, 5, 155–163. [Google Scholar]
- Manjamalai, A.; Berlin Grace, V.M. Antioxidant activity of essential oils from Wedelia chinensis (Osbeck) in vitro and in vivo lung cancer bearing C57BL/6 mice. Asian Pac. J. Cancer Prev. 2012, 13, 3065–3071. [Google Scholar] [CrossRef]
- Sattler, M.C.; Carvalho, C.R.; Clarindo, W.R. The polyploidy and its key role in plant breeding. Planta 2016, 243, 281–296. [Google Scholar] [CrossRef]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Hafiz, A.; Silvestri, C. Studies on colchicine induced chromosome doubling for enhancement of quality traits in ornamental plants. Plants 2019, 8, 194. [Google Scholar] [CrossRef] [PubMed]
- Niazian, M. Application of genetic and biotechnology for improving medicinal plants. Planta 2019, 249, 953–973. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, B.; Ebrahimzadeh, H. In vitro androgenesis: Spontaneous vs. artificial genome doubling and characterization of regenerants. Plant Cell Rep. 2020, 39, 299–316. [Google Scholar] [CrossRef]
- Niazian, M.; Nalousi, A.M. Artificial polyploidy induction for improvement of ornamental and medicinal plants. Plant Cell Tissue Organ Cult. 2020, 142, 447–469. [Google Scholar] [CrossRef]
- Touchell, D.H.; Palmer, J.E.; Ranney, T.G. In vitro ploidy manipulation for crop improvement. Front. Plant Sci. 2020, 11, 722. [Google Scholar] [CrossRef]
- Karamanou, M.; Tsoucalas, G.; Pantos, K.; Androutsos, G. Isolating colchicine in 19th century: An old drug revisited. Curr. Pharm. Des. 2018, 24, 654–658. [Google Scholar] [CrossRef]
- Salma, U.; Kundu, S.; Mandal, N. Artifical polyploidy in medicinal plants: Advancement in the last two decades and impending prospects. J. Crop Sci. Biotechnol. 2017, 20, 9–19. [Google Scholar] [CrossRef]
- Pradhan, S.K.; Gupta, R.C.; Goel, R.K. Differential content of secondary metabolites in diploid and tetraploid cytotypes of Siegesbeckia orientalis L. Nat. Prod. Res. 2018, 32, 2476–2482. [Google Scholar] [CrossRef]
- Švehlíková, V.; Repčák, M. Variation of apigenin quantity in diploid and tetraploid Chamomilla recutita (L.) Rauschert. Plant Biol. 2000, 2, 403–407. [Google Scholar] [CrossRef]
- De Jesus-Gonzalez, L.; Weathers, P.J. Tetraploid Artemisia annua hairy roots produce more artemisinin than diploids. Plant Cell Rep. 2003, 21, 809–813. [Google Scholar] [CrossRef]
- Majdi, M.; Karimzadeh, G.; Malboobi, M.A.; Omidbaigi, R.; Mirzaghaderi, G. Induction of tetraploidy to feverfew (Tanacetum parthenium Schulz-Bip.): Morphological, physiological, cytological, and phytochemical changes. HortScience 2010, 45, 16–21. [Google Scholar] [CrossRef]
- Xu, C.G.; Tang, T.X.; Chen, R.; Liang, C.H.; Liu, X.Y.; Wu, C.L.; Yang, Y.S.; Yang, D.P.; Wu, H. A comparative study of bioactive secondary metabolite production in diploid and tetraploid Echinacea purpurea (L.) Moench. Plant Cell Tissue Organ Cult. 2014, 116, 323–332. [Google Scholar] [CrossRef]
- Salma, U.; Kundu, S.; Harra, A.K.; Ali, M.N.; Mandal, N. Augmentation of wedelolactone through in vitro tetraploid induction in Eclipta alba (L.) Hassk. Plant Cell Tissue Organ Cult. 2018, 133, 289–298. [Google Scholar] [CrossRef]
- Bhuvaneswari, G.; Thirugnanasampandan, R.; Gogulramnath, M. Effect of colchicine induced tetraploidy on morphology, cytology, essential oil composition, gene expression and antioxidant activity of Citrus limon (L.) Osbeck. Physiol. Mol. Biol. Plant 2020, 26, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Noori, S.A.S.; Norouzi, M.; Karimzadeh, G.; Shirkool, K.; Niazian, M. Effect of colchicine-induced polyploidy on morphological characteristics and essential oil composition of ajowan (Trachyspermum ammi L.). Plant Cell Tissue Organ Cult. 2017, 130, 543–551. [Google Scholar] [CrossRef]
- Julião, S.A.; Ribeiro, C.D.V.; Lopes, J.M.L.; Matos, E.M.D.; Reis, A.C.; Peixoto, P.H.P.; Machado, M.A.; Azevedo, A.L.S.; Grazul, R.M.; Campos, J.M.S.D.; et al. Induction of synthetic polyploids and assessment of genome stability in Lippia alba. Front. Plant Sci. 2020, 11, 292. [Google Scholar] [CrossRef]
- Hannweg, K.; Visser, G.; de Jager, K.; Bertling, I. In vitro-induced polyploidy and its effect on horticultural characteristics, essential oil composition and bioactivity of Tetradenia riparia. S. Afr. J. Bot. 2016, 106, 186–191. [Google Scholar] [CrossRef]
- Nalawade, S.M.; Tsay, H.S. In vitro propagation of some important Chinese medicinal plants and their sustainable usage. In Vitro Cell. Dev. Biol. Plant 2004, 40, 143–154. [Google Scholar] [CrossRef]
- Gómez-Galera, S.; Pelacho, A.M.; Gené, A.; Capell, T.; Christou, P. The genetic manipulation of medicinal and aromatic plants. Plant Cell Rep. 2007, 26, 1689–1715. [Google Scholar] [CrossRef] [PubMed]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Tsai, C.H.; Tzeng, S.F.; Hsieh, S.C.; Lin, C.Y.; Tsai, C.J.; Chen, Y.R.; Yang, Y.C.; Chou, Y.W.; Lee, M.T.; Hsiao, P.W. Development of a standardized and effect-optimized herbal extract of Wedelia chinensis for prostate cancer. Phytomedicine 2015, 22, 406–414. [Google Scholar] [CrossRef]
- MassBank Europe. MassBank Record EA018355. Available online: https://www.massbank.eu (accessed on 18 May 2021).
- Li, M.; Si, D.; Fu, Z.; Sang, M.; Zhang, Z.; Liu, E.; Yang, W.; Gao, X.; Han, L. Enhanced identification of the in vivo metabolites of Ecliptae herba in rat plasma by integrating untargeted data-dependent MS2 and predictive multiple reaction monitoring-information dependent acquisition-enhanced product ion scan. J. Chromatogr. B 2019, 1109, 99–111. [Google Scholar] [CrossRef]
- Cheruvu, H.S.; Yadav, N.K.; Valicherla, G.R.; Arya, R.K.; Hussain, Z.; Sharma, C.; Arya, K.R.; Singh, R.K.; Datta, D.; Gayen, J.R. LC-MS/MS method for the simultaneous quantification of luteolin, wedelolactone and apigenin in mice plasma using hansen solubility parameters for liquid-liquid extraction: Application to pharmacokinetics of Eclipta alba chloroform fraction. J. Chromatogr. B 2018, 1081–1082, 76–86. [Google Scholar] [CrossRef]
- Zhou, K.; Fleet, P.; Nevo, E.; Zhang, X.; Sun, G. Transcriptome analysis reveals plant response to colchicine treatment during on chromosome doubling. Sci. Rep. 2017, 7, 8503. [Google Scholar] [CrossRef] [PubMed]
- Kaensaksiri, T.; Soontornchainaksaeng, P.; Soonthornchareonnon, N.; Prathanturarug, S. In vitro induction of polyploidy in Centella asiatica (L.) Urban. Plant Cell Tissue Organ Cult. 2011, 107, 187–194. [Google Scholar] [CrossRef]
- Chen, E.G.; Tsai, K.L.; Chung, H.H.; Chen, J.T. Chromosome doubling-enhanced biomass and dihydrotanshinone I production in Salvia miltiorrhiza, a traditional chinese medicinal plant. Molecules 2018, 23, 3106. [Google Scholar] [CrossRef] [PubMed]
- Zafar, R.; Sagar, B.P.S. In vitro plant regeneration of Eclipta alba and increased production of coumestans. Fitoterapia 1999, 70, 348–358. [Google Scholar] [CrossRef]
- Sagar, B.P.S.; Panwar, R.; Goswami, A.; Kadian, K.; Tyagi, K.; Chugh, M.; Dalal, S.; Zafar, R. Pharmacokinetic interactions of antihepatotoxic wedelolactone with paracetamol in wistar albino rats. Pharm. Biol. 2006, 44, 554–561. [Google Scholar] [CrossRef]
- Lee, Y.J.; Lin, W.L.; Chen, N.F.; Chuang, S.K.; Tseng, T.H. Demethylwedelolactone derivatives inhibit invasive growth in vitro and lung metastasis of MDA-MB-231 breast cancer cells in nude mice. Eur. J. Med. Chem. 2012, 56, 361–367. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid biosynthesis: A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef]
- To, K.Y.; Wang, C.K. Molecular breeding of flower color. In Floriculture, Ornamental and Plant Biotechnology, Volume I; Teixeira da Silva, J.A., Ed.; Global Science Books: Middlesex, UK, 2006; pp. 300–310. [Google Scholar]
- Martin, Y.P.; Beena, M.R.; Joseph, D. High frequency axillary bud multiplication and ex vitro rooting of Wedelia chinensis (Osbeck) Merr.—A medicinal plant. Indian J. Exp. Biol. 2003, 41, 262–266. [Google Scholar] [PubMed]
- Rahman, M.M.; Bhadra, S.K. Development of protocol for in vitro culture and rapid propagation of Wedelia chinensis (Osbeek) Merr. J. Med. Plant Res. 2011, 5, 2387–2392. [Google Scholar]
- Animal Genome Size Database. Available online: http://www.genomesize.com (accessed on 18 May 2021).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Formula | Molecular Weight | Retention Time (min) | Precursor Ion Spectra (m/z) | Product Ion Spectra (m/z) | Reference |
|---|---|---|---|---|---|---|
| Sulfadimethoxine | C12H13N4S | 309.07 | 3.55 | 309 | 230 | [29] |
| Demethylwedelolactone | C15H8O7 | 300.22 | 2.57 | 299 | 227 | [30] |
| Luteolin | C15H10O6 | 285.04 | 3.61 | 285 | 133 | [31] |
| Wedelolactone | C16H10O7 | 314.25 | 3.80 | 313 | 298 | [31] |
| Apigenin | C15H10O5 | 269.05 | 4.27 | 269 | 117 | [31] |
| Sample | Demethylwedelolactone | Luteolin | Wedelolactone | Apigenin |
|---|---|---|---|---|
| Root | n.d. | 2.06 ± 0.17 | n.d. | n.d. |
| Stem | n.d. | n.d. | n.d. | n.d. |
| Leaf | 145.34 ± 6.95 | 85.85 ± 5.54 | 18.51 ± 3.27 | 34.13 ± 4.98 |
| Plant No. | Polyploidy | Demethylwedelolactone | Luteolin | Wedelolactone | Apigenin |
|---|---|---|---|---|---|
| WT | 2n | 183.00 ± 6.44 | 104.80 ± 7.37 | 18.19 ± 8.10 | 21.21 ± 4.39 |
| P1 | 4n | 16.48 ± 8.10 | 87.44 ± 11.23 | 4.55 ± 1.55 | 29.58 ± 4.96 |
| P2 | 4n | 261.67 ± 15.63 | 76.86 ± 10.55 | 12.85 ± 4.79 | 20.16 ± 4.67 |
| P3 | 3n | 44.24 ± 11.57 | 65.68 ± 12.12 | 6.29 ± 1.48 | 20.09 ± 3.85 |
| P4 | 3n | 189.90 ± 11.54 | 107.07 ± 10.16 | 13.24 ± 4.12 | 26.03 ± 6.61 |
| P6 | 3n | 274.65 ± 13.67 | 91.61 ± 17.77 | 22.74 ± 4.19 | 15.53 ± 3.59 |
| P7 | 3n | 143.83 ± 14.71 | 90.44 ± 8.31 | 14.59 ± 2.58 | 20.57 ± 2.61 |
| P8 | 4n | 15.61 ± 3.72 | 50.67 ± 6.08 | 11.50 ± 2.23 | 17.54 ± 3.24 |
| P9 | 3n | 124.69 ± 6.26 | 70.38 ± 7.90 | 16.43 ± 2.64 | 23.50 ± 4.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, Y.-T.; Chen, P.-Y.; To, K.-Y. Induction of Polyploidy and Metabolic Profiling in the Medicinal Herb Wedelia chinensis. Plants 2021, 10, 1232. https://doi.org/10.3390/plants10061232
Tsai Y-T, Chen P-Y, To K-Y. Induction of Polyploidy and Metabolic Profiling in the Medicinal Herb Wedelia chinensis. Plants. 2021; 10(6):1232. https://doi.org/10.3390/plants10061232
Chicago/Turabian StyleTsai, Yung-Ting, Po-Yen Chen, and Kin-Ying To. 2021. "Induction of Polyploidy and Metabolic Profiling in the Medicinal Herb Wedelia chinensis" Plants 10, no. 6: 1232. https://doi.org/10.3390/plants10061232
APA StyleTsai, Y.-T., Chen, P.-Y., & To, K.-Y. (2021). Induction of Polyploidy and Metabolic Profiling in the Medicinal Herb Wedelia chinensis. Plants, 10(6), 1232. https://doi.org/10.3390/plants10061232