
Evaluating the Endophytic Activities of Beauveria bassiana on the Physiology, Growth, and Antioxidant Activities of Extracts of Lettuce (Lactuca sativa L.)


Abstract

1. Introduction
2. Results
2.1. Pathogenicity Assessment
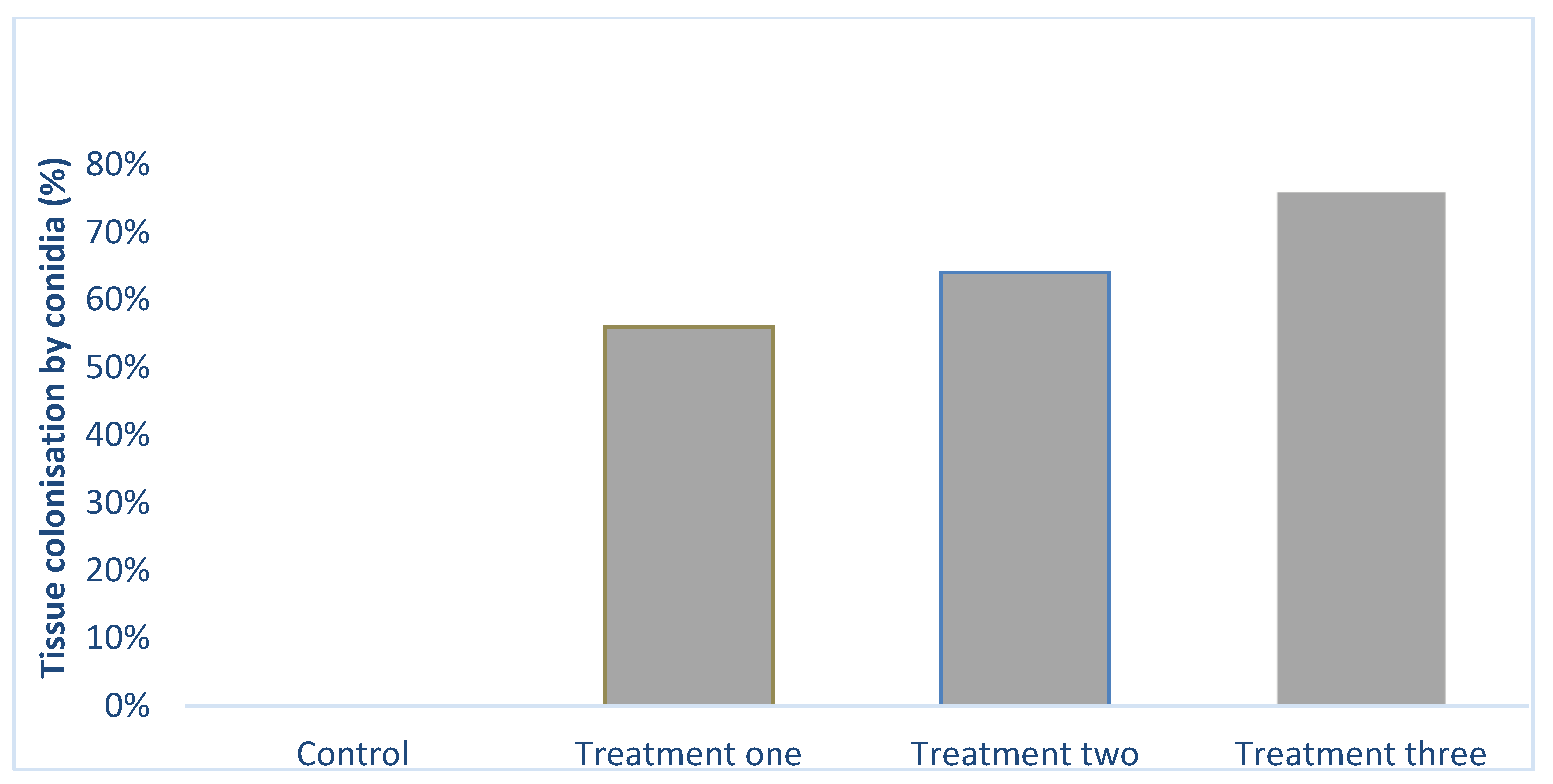
2.2. Colonization of Tissues by Fungus
2.3. Effect of Fungus on Plant Height, Crown Size and Roots Length
Effect of Fungus on Fresh Weight and Dry Weight of the Root and Aerial Parts of Lettuce Plants
2.4. Effect of Fungus (Beauveria bassiana) on Antioxidant Capacity: Ferric Reducing Antioxidant Power (FRAP) and Trolox Equivalent Antioxidant Capacity (TEAC)
2.5. Effect of Beauveria bassiana on Tissue Analysis
2.5.1. Macronutrients
2.5.2. Micronutrients
2.6. Proximate Results
2.6.1. Protein
2.6.2. Fatty Acids
2.7. Chlorophyll
3. Discussion
4. Materials and Methods
4.1. Research Design
4.2. Plants Material
4.3. Fungus Preparation
4.4. Pathogenicity Against Myzus Persicae
4.5. Greenhouse Study
4.6. Antioxidants
4.6.1. Sample Material
4.6.2. Ferric Reducing Antioxidant Power (FRAP)
4.6.3. Trolox Equivalent Antioxidant Capacity (TEAC)
4.7. Tissue Nutrient Content Analysis
4.8. Proximate Analysis
4.8.1. Sample Preparation
4.8.2. Protein Analysis
4.8.3. Fatty Acid Analysis
4.9. Chlorophyll Content Analysis
Estimation of Chlorophyll Content
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrière, V.; Lecompte, F.; Lescourret, F. Efficacy of pest and pathogen control, yield and quality of winter lettuce crops managed with reduced pesticide applications. Eur. J. Agron. 2015, 71, 34–43. [Google Scholar] [CrossRef]
- Chandi, R.S.; Kataria, S.K.; Kaur, J. Arthropods as Vector of Plant Pathogens Viz-a-Viz Their Management. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 4006–4023. [Google Scholar] [CrossRef]
- Koch, R.L.; Hodgson, E.W.; Knodel, J.J.; Varenhorst, A.J.; Potter, B.D. Management of Insecticide-Resistant Soybean Aphids in the Upper Midwest of the United States. J. Integr. Pest Manag. 2018, 9, 1–7. [Google Scholar] [CrossRef]
- Ricupero, M.; Desneux, N.; Zappalà, L.; Biondi, A. Target and non-target impact of systemic insecticides on a polyphagous aphid pest and its parasitoid. Chemosphere 2020, 247, 125728. [Google Scholar] [CrossRef] [PubMed]
- Furlan, L.; Pozzebon, A.; Duso, C.; Simon-Delso, N.; Sánchez-Bayo, F.; Marchand, P.A.; Codato, F.; van Lexmond, M.B.; Bonmatin, J.-M. An update of the Worldwide Integrated Assessment (WIA) on systemic insecticides. Part 3: Alternatives to systemic insecticides. Environ. Sci. Pollut. Res. 2021, 28, 11798–11820. [Google Scholar] [CrossRef]
- Nair, D.N.; Padmavathy, S. Impact of Endophytic Microorganisms on Plants, Environment and Humans. Sci. World J. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.E.; Posada, F.; Aime, M.C.; Pava-Ripoll, M.; Infante, F.; Rehner, S.A. Entomopathogenic fungal endophytes. Biol. Control. 2008, 46, 72–82. [Google Scholar] [CrossRef]
- Shahid, A.; Rao, Q.; Bakhsh, A.; Husnain, T. Entomopathogenic fungi as biological controllers: New insights into their virulence and pathogenicity. Arch. Biol. Sci. 2012, 64, 21–42. [Google Scholar] [CrossRef]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Shah, S.; Shrestha, R.; Maharjan, S.; Selosse, M.A.; Pant, B. Isolation and characterization of plant growth-promoting endophytic fungi from the roots of Dendrobium moniliforme. Plants 2019, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Chand, K.; Shah, S.; Sharma, J.; Paudel, M.R.; Pant, B. Isolation, characterization, and plant growth-promoting activities of endophytic fungi from a wild orchid Vanda cristata. Plant Signal. Behav. 2020, 15, 1744294. [Google Scholar] [CrossRef]
- Omomowo, O.I.; Babalola, O.O. Bacterial and Fungal Endophytes: Tiny Giants with Immense Beneficial Potential for Plant Growth and Sustainable Agricultural Productivity. Microorganisms 2019, 7, 481. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Lata, C. Heavy Metal Stress, Signaling, and Tolerance Due to Plant-Associated Microbes: An Overview. Front. Plant Sci. 2018, 9, 452. [Google Scholar] [CrossRef] [PubMed]
- Lugtenberg, B.J.; Caradus, J.R.; Johnson, L.J. Fungal endophytes for sustainable crop production. FEMS Microbiol. Ecol. 2016, 92. [Google Scholar] [CrossRef]
- Hu, S.; Bidochka, M.J. Root colonization by endophytic insect-pathogenic fungi. J. Appl. Microbiol. 2021, 130, 570–581. [Google Scholar] [CrossRef]
- Afandhi, A.; Widjayanti, T.; Emi, A.A.L.; Tarno, H.; Afiyanti, M.; Handoko, R.N.S. Endophytic fungi Beauveria bassiana Balsamo accelerates growth of common bean (Phaeseolus vulgaris L.). Chem. Biol. Technol. Agric. 2019, 6, 11. [Google Scholar] [CrossRef]
- Bamisile, B.S.; Dash, C.K.; Akutse, K.S.; Keppanan, R.; Wang, L. Fungal Endophytes: Beyond Herbivore Management. Front. Microbiol. 2018, 9, 544. [Google Scholar] [CrossRef] [PubMed]
- Saia, S.; Colla, G.; Raimondi, G.; Di Stasio, E.; Cardarelli, M.; Bonini, P.; Vitaglione, P.; De Pascale, S.; Rouphael, Y. An endophytic fungi-based biostimulant modulated lettuce yield, physiological and functional quality responses to both moderate and severe water limitation. Sci. Hortic. 2019, 256, 108595. [Google Scholar] [CrossRef]
- Ren, A.Z.; Li, X.; Han, R.; Yin, L.J.; Wei, M.Y.; Gao, Y.B. Benefits of a symbiotic association with endophytic fungi are subject to water and nutrient availability in Achnatherum sibiricum. Plant Soil 2011, 346, 363–373. [Google Scholar] [CrossRef]
- White, J.F.; Torres, M.S. Is plant endophyte-mediated defensive mutualism the result of oxidative stress protection? Physiol. Plant. 2010, 138, 440–446. [Google Scholar] [CrossRef]
- Vinale, F.; Nicoletti, R.; Lacatena, F.; Marra, R.; Sacco, A.; Lombardi, N.; D’Errico, G.; Digilio, M.C.; Lorito, M.; Woo, S.L. Secondary metabolites from the endophytic fungus Talaromyces pinophilus. Nat. Prod. Res. 2017, 31, 1778–1785. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Su, T.-J.; Cai, S.-M.; Wu, W. Fungal endophyte-derived Fritillaria unibracteata var. wabuensis: diversity, antioxidant capacities in vitro and relations to phenolic, flavonoid or saponin compounds. Sci. Rep. 2017, 7, srep42008. [Google Scholar] [CrossRef] [PubMed]
- Eotieno, N.; Lally, R.D.; Ekiwanuka, S.; Elloyd, A.; Eryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef]
- Waqas, M.; Khan, A.L.; Kamran, M.; Hamayun, M.; Kang, S.-M.; Kim, Y.-H.; Lee, I.-J. Endophytic Fungi Produce Gibberellins and Indoleacetic Acid and Promotes Host-Plant Growth during Stress. Molecules 2012, 17, 10754–10773. [Google Scholar] [CrossRef]
- Ismail, I.; Hamayun, M.; Sayyed, A.; Din, I.U.; Gul, H.; Hussain, A. Gibberellin and indole acetic acid production capacity of endophytic fungi isolated from Zea mays L. Int. J. Biosci. 2016, 8, 35–43. [Google Scholar] [CrossRef]
- Jaber, L.R.; Enkerli, J. Fungal entomopathogens as endophytes: Can they promote plant growth? Biocontrol Sci. Technol. 2017, 27, 28–41. [Google Scholar] [CrossRef]
- Lata, R.; Chowdhury, S.; Gond, S.K.; White, J.F. Induction of abiotic stress tolerance in plants by endophytic microbes. Lett. Appl. Microbiol. 2018, 66, 268–276. [Google Scholar] [CrossRef]
- Ikram, M.; Ali, N.; Jan, G.; Jan, F.G.; Rahman, I.U.; Iqbal, A.; Hamayun, M. IAA producing fungal endophyte Penicillium roqueforti Thom., enhances stress tolerance and nutrients uptake in wheat plants grown on heavy metal contaminated soils. PLoS ONE 2018, 13, e0208150. [Google Scholar] [CrossRef]
- Ismail, I.; Hamayun, M.; Hussain, A.; Iqbal, A.; Khan, S.A.; Lee, I.-J. Endophytic Fungus Aspergillus japonicus Mediates Host Plant Growth under Normal and Heat Stress Conditions. BioMed Res. Int. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Branine, M.; Bazzicalupo, A.; Branco, S. Biology and applications of endophytic insect-pathogenic fungi. PLoS Pathog. 2019, 15, e1007831. [Google Scholar] [CrossRef]
- Rajab, L.; Ahmad, M.; Gazal, I. Endophytic establishment of the fungal entomopathogen, Beauveria bassiana (Bals.) Vuil., in cucumber plants. Egypt. J. Biol. Pest Control. 2020, 30, 1–7. [Google Scholar] [CrossRef]
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monzón, A.; Ownley, B.H.; et al. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef]
- Nishi, O.; Sushida, H.; Higashi, Y.; Iida, Y. Epiphytic and endophytic colonisation of tomato plants by the entomopathogenic fungus Beauveria bassiana strain GHA. Mycology 2021, 12, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Heviefo, A.G.; Nyamador, S.W.; Glitho, I.A.; Tamo, M. Establishment of the Fungal Entomopathogen Beauveria bassiana as Endophyte in Cabbage Plant for Disease And Lepidopteran Larvae Pest Control. Bulletin de la Recherche Agronomique du Bénin (BRAB), Numéro Spécial Développement Agricole Durable (DAD)-Décembre 2017, 1–12. Available online: https://www.researchgate.net/publication/323734527_Establishment_of_the_fungal_entomopathogen_Beauveria_bassiana_as_endophyte_in_cabbage_plant_Brassica_oleracea_for_disease_and_lepidopteran_larvae_pest_control (accessed on 30 March 2021).
- Ai, Z.; Wang, G.; Liang, C.; Liu, H.; Zhang, J.; Xue, S.; Liu, G. The Effects of nitrogen addition on the uptake and allocation of macro- and micronutrients in Bothriochloa ischaemum on Loess Plateau in China. Front. Plant Sci. 2017, 8, 1476. [Google Scholar] [CrossRef] [PubMed]
- Ucar, E.; Ozyigit, Y.; Demirbas, A.; Guven, D.Y.; Turgut, K. Effect of different nitrogen doses on dry matter ratio, chlorophyll and macro/micro nutrient content in sweet herb (Stevia rebaudiana Bertoni). Commun. Soil Sci. Plant Anal. 2017, 48, 1231–1239. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.; Rahmat, A.; Rahman, Z.A. Effects of Nitrogen Fertilization on Synthesis of Primary and Secondary Metabolites in Three Varieties of Kacip fatimah (Labisia, Pumila, and Blume). Int. J. Mol. Sci. 2011, 12, 5238–5254. [Google Scholar] [CrossRef]
- Radušienė, J.; Marksa, M.; Ivanauskas, L.; Jakštas, V.; Çalişkan, Ö.; Kurt, D.; Odabaş, M.S.; Çirak, C. Effect of nitrogen on herb production, secondary metabolites and antioxidant activities of Hypericum pruinatum under nitrogen application. Ind. Crop. Prod. 2019, 139, 111519. [Google Scholar] [CrossRef]
- Chen, Z.; Jin, Y.; Yao, X.; Chen, T.; Wei, X.; Li, C.; White, J.F.; Nan, Z. Fungal endophyte improves survival of Lolium perenne in low fertility soils by increasing root growth, metabolic activity and absorption of nutrients. Plant Soil 2020, 452, 185–206. [Google Scholar] [CrossRef]
- Jurić, S.; Stracenski, K.S.; Król-Kilińska, Ż.; Žutić, I.; Uher, S.F.; Đermić, E.; Topolovec-Pintarić, S.; Vinceković, M. The enhancement of plant secondary metabolites content in Lactuca sativa L. by encapsulated bioactive agents. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Yang, L.; Zou, Y.-N.; Tian, Z.-H.; Wu, Q.-S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Javed, K.; Javed, H.; Mukhtar, T.; Qiu, D. Pathogenicity of some entomopathogenic fungal strains to green peach aphid, Myzus persicae Sulzer (Homoptera: Aphididae). Egypt. J. Biol. Pest Control. 2019, 29, 1–7. [Google Scholar] [CrossRef]
- Motholo, L.; Booyse, M.; Hatting, J.; Tsilo, T.; Thekisoe, O. Pathogenicity of Beauveria bassiana (Hypocreales: Cordycipitaceae) against the Russian Wheat Aphid, Diuraphis noxia (Hemiptera: Aphididae). Afr. Èntomol. 2020, 28, 455–461. [Google Scholar] [CrossRef]
- Cheong, P.C.; Glare, T.R.; Rostás, M.; Haines, S.; Brookes, J.J.; Ford, S. Lack of involvement of chitinase in direct toxicity of Beauveria bassiana cultures to the aphid Myzus persicae. J. Invertebr. Pathol. 2020, 169, 107276. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Feng, J.; Fan, Y.; Zhang, Y.; Bidochka, M.J.; Leger, R.J.S.; Pei, Y. Expressing a fusion protein with protease and chitinase activities increases the virulence of the insect pathogen Beauveria bassiana. J. Invertebr. Pathol. 2009, 102, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Bayissa, W.; Ekesi, S.; Mohamed, S.A.; Kaaya, G.P.; Wagacha, J.M.; Hanna, R.; Maniania, N.K. Selection of fungal isolates for virulence against three aphid pest species of crucifers and okra. Anz. Schädlingskunde Pflanz. Umweltschutz 2017, 90, 355–368. [Google Scholar] [CrossRef]
- Jaber, L.R.; Ownley, B.H. Can we use entomopathogenic fungi as endophytes for dual biological control of insect pests and plant pathogens? Biol. Control. 2018, 116, 36–45. [Google Scholar] [CrossRef]
- Moloinyane, S.; Nchu, F. The effects of endophytic Beauveria bassiana inoculation on infestation level of Planococcus ficus, growth and volatile constituents of potted greenhouse grapevine (Vitis vinifera L.). Toxins 2019, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Muvea, A.M.; Meyhöfer, R.; Subramanian, S.; Poehling, H.-M.; Ekesi, S.; Maniania, N.K. Colonization of onions by endophytic fungi and their impacts on the biology of Thrips tabaci. PLoS ONE 2014, 9, e108242. [Google Scholar] [CrossRef]
- Dash, C.K.; Bamisile, B.S.; Keppanan, R.; Qasim, M.; Lin, Y.; Islam, S.U.; Hussain, M.; Wang, L. Endophytic entomopathogenic fungi enhance the growth of Phaseolus vulgaris L. (Fabaceae) and negatively affect the development and reproduction of Tetranychus urticae Koch (Acari: Tetranychidae). Microb. Pathog. 2018, 125, 385–392. [Google Scholar] [CrossRef]
- Gayler, S.; Grams, T.E.E.; Heller, W.; Treutter, D.; Priesack, E. A dynamical model of environmental effects on allocation to carbon-based secondary compounds in juvenile trees. Ann. Bot. 2007, 101, 1089–1098. [Google Scholar] [CrossRef]
- Araya, T.; Noguchi, K.; Terashima, I. Effect of nitrogen nutrition on the carbohydrate repression of photosynthesis in leaves of Phaseolus vulgaris L. J. Plant Res. 2009, 123, 371–379. [Google Scholar] [CrossRef]
- Rozpądek, P.; Wężowicz, K.; Nosek, M.; Ważny, R.; Tokarz, K.; Lembicz, M.; Miszalski, Z.; Turnau, K. The fungal endophyte Epichloë typhina improves photosynthesis efficiency of its host orchard grass (Dactylis glomerata). Planta 2015, 242, 1025–1035. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Bush, D.R. Carbohydrate export from the leaf: a highly regulated process and target to enhance photosynthesis and productivity. Plant Physiol. 2011, 155, 64–69. [Google Scholar] [CrossRef]
- Smith, A.M.; Stitt, M. Coordination of carbon supply and plant growth. Plant Cell Environ. 2007, 30, 1126–1149. [Google Scholar] [CrossRef]
- Sun, M.; Liu, X. Carbon requirements of some nematophagous, entomopathogenic and mycoparasitic hyphomycetes as fungal biocontrol agents. Mycopathology 2006, 161, 295–305. [Google Scholar] [CrossRef] [PubMed]
- White, J.F.; Kingsley, K.L.; Zhang, Q.; Verma, R.; Obi, N.; Dvinskikh, S.; Elmore, M.T.; Verma, S.K.; Gond, S.K.; Kowalski, K.P. Review: Endophytic microbes and their potential applications in crop management. Pest Manag. Sci. 2019, 75, 2558–2565. [Google Scholar] [CrossRef] [PubMed]
- Safavi, S.A.; Shah, F.A.; Pakdel, A.K.; Rasoulian, G.R.; Bandani, A.R.; Butt, T.M. Effect of nutrition on growth and virulence of the entomopathogenic fungus Beauveria bassiana. FEMS Microbiol. Lett. 2007, 270, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Vergara, C.; Araujo, K.E.C.; Sperandio, M.V.L.; Santos, L.A.; Urquiaga, S.; Zilli, J. Édson Dark septate endophytic fungi increase the activity of proton pumps, efficiency of 15N recovery from ammonium sulphate, N content, and micronutrient levels in rice plants. Braz. J. Microbiol. 2019, 50, 825–838. [Google Scholar] [CrossRef]
- Rout, G.R.; Sahoo, S. Role of iron in plant growth and metabolism. Rev. Agric. Sci. 2015, 3, 1–24. [Google Scholar] [CrossRef]
- Uluisik, I.; Karakaya, H.C.; Koc, A. The importance of boron in biological systems. J. Trace Elements Med. Biol. 2018, 45, 156–162. [Google Scholar] [CrossRef]
- Ruscitti, M.; Arango, M.; Beltrano, J. Improvement of copper stress tolerance in pepper plants (Capsicum annuum L.) by inoculation with arbuscular mycorrhizal fungi. Theor. Exp. Plant Physiol. 2017, 29, 37–49. [Google Scholar] [CrossRef]
- Schmidt, S.B.; Husted, S. The biochemical properties of manganese in plants. Plants 2019, 8, 381. [Google Scholar] [CrossRef] [PubMed]
- Luciano, Á.J.; Irineo, T.P.; Ocampo-Velázquez, R.V.; Feregrino-Pérez, A.A.; Hernández, A.C.; Guevara-González, R.G. Inte-grating plant nutrients and elicitors for production of secondary metabolites, sustainable crop production and human health: A review. Int. J. Agric. Biol. 2017, 19, 391–402. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Hossain, S.; Al Mahmud, J.; Rahman, A.; Inafuku, M.; Oku, H.; Fujita, M. Coordinated actions of glyoxalase and antioxidant defense systems in conferring abiotic stress tolerance in plants. Int. J. Mol. Sci. 2017, 18, 200. [Google Scholar] [CrossRef]
- Carrasco-Gil, S.; Hernandez-Apaolaza, L.; Lucena, J.J. Effect of several commercial seaweed extracts in the mitigation of iron chlorosis of tomato plants (Solanum lycopersicum L.). Plant Growth Regul. 2018, 86, 401–411. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, S.; Gaur, S.; Singh, S.; Yadav, V.; Liu, S.; Singh, V.P.; Sharma, S.; Srivastava, P.; Prasad, S.M.; et al. Acquisition and homeostasis of iron in higher plants and their probable role in abiotic stress tolerance. Front. Environ. Sci. 2018, 5, 1–15. [Google Scholar] [CrossRef]
- Bidart-Bouzat, M.G.; Imeh-Nathaniel, A. Global change effects on plant chemical defenses against insect herbivores. J. Integr. Plant Biol. 2008, 50, 1339–1354. [Google Scholar] [CrossRef]
- Ray, S.; Singh, V.; Singh, S.; Sarma, B.K.; Singh, H.B. Biochemical and histochemical analyses revealing endophytic Alcaligenes faecalis mediated suppression of oxidative stress in Abelmoschus esculentus challenged with Sclerotium rolfsii. Plant Physiol. Biochem. 2016, 109, 430–441. [Google Scholar] [CrossRef]
- Shahabivand, S.; Parvaneh, A.; Aliloo, A.A. Root endophytic fungus Piriformospora indica affected growth, cadmium partitioning and chlorophyll fluorescence of sunflower under cadmium toxicity. Ecotoxicol. Environ. Saf. 2017, 145, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, F.; Markgren, J.; Hedenqvist, M.; Johansson, E. Modeling to understand plant protein structure-function relationships—implications for seed storage proteins. Molecules 2020, 25, 873. [Google Scholar] [CrossRef]
- Idhan, A.; Nadir, M.; Kadir, M. Paddy chlorophyll concentrations in drought stress condition and endophytic fungi application. IOP Conf. Ser. Earth Environ. Sci. 2018, 156, 012040. [Google Scholar] [CrossRef]
- Latifian, M.; Rad, B. Pathogenicity of the entomopathogenic fungi Beauveria bassiana (Balsamo) Vuillmin, Beauveria brongniartii Saccardo and Metarhizium anisopliae Metsch to adult Oryctes elegans Prell and effects on feeding and fecundity. Intl. J. Agri. Crop Sci. 2012, 4, 1026–1032. [Google Scholar]
- Nazir, T.; Basit, A.; Hanan, A.; Majeed, M.; Qiu, D. In vitro pathogenicity of some entomopathogenic fungal strains against green peach aphid Myzus persicae (Homoptera: Aphididae). Agronomy 2018, 9, 7. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Miller, N.J.; Rice-Evans, C.; Davies, M.J. A new method for measuring antioxidant activity. Biochem. Soc. Trans. 1993, 21, 95S. [Google Scholar] [CrossRef]
- Miller, R.O. High-Temperature Oxidation: Dry Ashing. In Handbook of Reference Methods for Plant Analysis; CRC Press: Boca Raton, FL, USA, 1998; pp. 53–56, Chapter 5. [Google Scholar]
- Chikwanha, O.C.; Muchenje, V.; Nolte, J.E.; Dugan, M.E.; Mapiye, C. Grape pomace (Vitis vinifera L. cv. Pinotage) supplementation in lamb diets: Effects on growth performance, carcass and meat quality. Meat Sci. 2019, 147, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Sukhija, P.S.; Palmquist, D.L. Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. J. Agric. Food Chem. 1988, 36, 1202–1206. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Rajalakshmi, K.; Banu, N. Extraction and estimation of chlorophyll from medicinal plants. Int. J. Sci. Res. 2015, 4, 209–212. [Google Scholar]
- Finney, D.J. Probit Analysis, 2nd ed.; Cambridge University Press: Cambridge, UK, 1952; pp. 388–390. [Google Scholar]


{kind=link}
{kind=link}
| Treatments | Mean ± SE Number of Dead Insects and (Abbott-Corrected Percentage Mortality) |
|---|---|
| Control | 0 ± 0 |
| 1 × 106 conidia mL−1 | 48 ± 0.50 |
| 1 × 107 conidia mL−1 | 66 ± 0.31 |
| 1× 108 conidia mL−1 | 78 ± 0.18 |
| LC50 Fiducial limits (95% CI) | 1.2 × 106 conidia mL−1 1.1 × 106–1.6 × 106 conidia mL−1 |
| Treatments | Roots Length (cm) | Plant Height (cm) | Crown Size (cm) |
|---|---|---|---|
| Control | 19.50 ± 0.54 a | 15.52 ± 0.10 a | 25.94 ± 0.19 a |
| 1 × 106 conidia mL−1 | 19.33 ± 0.47 a | 16.02 ± 0.12 b | 27.50 ± 0.27 bc |
| 1 × 107 conidia mL−1 | 20.10 ± 0.51 a | 16.02 ± 0.12 b | 26.96 ± 0.29 b |
| 1 × 108 conidia mL−1 | 20.33 ± 0.38 a | 16.04 ± 0.18 b | 28.14 ± 0.22 c |
| Treatments | Roots Fresh Weight (g) | Plant Fresh Weight (g) | Roots Dry Weight(g) | Plant Dry Weight (g) |
|---|---|---|---|---|
| Control | 23.57 ± 0.62 a | 58.70 ± 1.60 a | 3.18 ± 0.12 a | 4.24 ± 0.11 a |
| 1 × 106 conidia mL−1 | 23.12 ± 1.21 a | 61.70 ± 1.53 a | 3.30 ± 0.06 a | 4.49 ± 0.13 a |
| 1 × 107 conidia mL−1 | 20. 48 ± 1.00 a | 54.17 ± 2.26 a | 3.39 ± 0.11 a | 4.61 ± 0.11 a |
| 1 × 108 conidia mL−1 | 21.28 ± 0.95 a | 57.65 ± 2.43 a | 3.32 ± 0.10 a | 4.49 ± 0.18 a |
| Treatments | Frap (µmol AAE/g) Mean ± SE | TEAC (µmol TE/g) Mean ± SE |
|---|---|---|
| Control | 86.13 ± 6.35 a | 88.92 ± 7.02 a |
| 1 × 106 conidia mL−1 | 46.15 ± 3.61 b | 32.17 ± 6.43 b |
| 1 × 107 conidia mL−1 | 55.81 ± 13.15 b | 43.14 ± 4.40 b |
| 1 × 108 conidia mL−1 | 89.43 ± 9.10 a | 97.97 ± 5.30 a |
| Treatments | C | N | P | K | Ca | Mg | Na |
|---|---|---|---|---|---|---|---|
| Control | 407.36 ± 0.41 a | 20.90 ± 0.60 a | 4.10 ± 0.60 a | 55.33 ± 0.33 a | 10.10 ± 0.45 a | 4.27 ± 0.15 a | 2.43 ± 0.14 a |
| 1 × 106 conidia mL−1 | 365.50 ± 3.36 b | 21.37 ± 0.75 a | 4.67 ± 0.38 a | 60.00 ± 4.16 a | 11.67 ± 0.88 a | 5.27 ± 0.56 a | 2.58 ± 0.19 a |
| 1 × 107 conidia mL−1 | 363.13 ± 1.48 b | 22.13 ± 0.47 a | 4.53 ± 0.18 a | 60.67 ± 0.67 a | 10.33 ± 0.33 a | 5.20 ± 0.23 a | 2.42 ± 0.04 a |
| 1 × 108 conidia mL−1 | 396.33 ± 6.55 a | 21.20 ± 0.81 a | 4.60 ± 0.30 a | 58.33 ± 2.73 a | 10.03 ± 0.48 a | 4.63 ± 0.20 a | 2.44 ± 0.29 a |
| Treatments | Mn | Fe | Cu | Zn | B |
|---|---|---|---|---|---|
| Control | 54.73 ± 5.25 a | 286.00 ± 17.47 a | 2.87 ± 0.33 a | 47.27 ± 7.13 a | 38.70 ± 1.29 a |
| 1 × 106 conidia mL−1 | 70.63 ± 7.58 ab | 439.33 ± 41.91 b | 5.00 ± 0.15 b | 39.77 ± 3.48 a | 45.43 ± 2.57 ab |
| 1 × 107 conidia mL−1 | 81.03 ± 4.39 b | 427.33 ± 2.85 b | 5.90 ± 0.26 b | 38.33 ± 4.99 a | 50.27 ± 1.01 b |
| 1 × 108 conidia mL−1 | 72.60 ± 1.99 ab | 464.67 ± 34.36 b | 3.57 ± 0.35 a | 36.10 ± 2.21 a | 46.27 ± 3.44 ab |
| Treatments | Protein (g kg−1) Mean ± SE |
|---|---|
| Control | 27.87 ± 2.02 ab |
| 1 × 106 conidia mL−1 | 23.20 ± 1.19 a |
| 1 × 107 conidia mL−1 | 32.01 ± 2.25 ab |
| 1 × 108 conidia mL−1 | 34.06 ± 2.26 b |
| Treatments | Palmitic Acid Mean ± SE | Linoleic Acid Mean ± SE | Linolenic Acid Mean ± SE | Total Fatty Acids Mean ± SE |
|---|---|---|---|---|
| Control | 366.67 ± 33.33 a | 266.67 ± 33.33 a | 933.33 ± 166.67 a | 1566.67 ± 233.33 a |
| 1 × 106 conidia mL−1 | 466.67 ± 166.67 a | 200.00 ± 0.00 a | 566.67 ± 33.33 a | 1233.33 ± 185.59 a |
| 1 × 107 conidia mL−1 | 300.00 ± 0.00 a | 200.00 ± 0.00 a | 733.33 ± 33.33 a | 1233.00 ± 33.33 a |
| 1 × 108 conidia mL−1 | 400.00 ± 0.00 a | 200.00 ± 0.00 a | 833.33 ± 33.33 a | 1433.33 ± 33.33 a |
| Treatments | Total Chlorophyll | Chlorophyll a | Chlorophyll b |
|---|---|---|---|
| Control | 503.13 ± 36.73 a | 368.51 ± 28.31 a | 134.75 ± 8.45 a |
| 1 × 106 conidia mL−1 | 488.93 ± 6.71 a | 355.16 ± 5.15 a | 133.89 ± 1.56 a |
| 1 × 107 conidia mL−1 | 520.84 ± 25.97 a | 381.12 ± 17.63 a | 139.85 ± 8.38 a |
| 1 × 108 conidia mL−1 | 586.51 ± 31.21 a | 425.39 ± 22.37 a | 161.26 ± 8.82 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Macuphe, N.; Oguntibeju, O.O.; Nchu, F. Evaluating the Endophytic Activities of Beauveria bassiana on the Physiology, Growth, and Antioxidant Activities of Extracts of Lettuce (Lactuca sativa L.). Plants 2021, 10, 1178. https://doi.org/10.3390/plants10061178
Macuphe N, Oguntibeju OO, Nchu F. Evaluating the Endophytic Activities of Beauveria bassiana on the Physiology, Growth, and Antioxidant Activities of Extracts of Lettuce (Lactuca sativa L.). Plants. 2021; 10(6):1178. https://doi.org/10.3390/plants10061178
Chicago/Turabian StyleMacuphe, Neo, Oluwafemi Omoniyi Oguntibeju, and Felix Nchu. 2021. "Evaluating the Endophytic Activities of Beauveria bassiana on the Physiology, Growth, and Antioxidant Activities of Extracts of Lettuce (Lactuca sativa L.)" Plants 10, no. 6: 1178. https://doi.org/10.3390/plants10061178
APA StyleMacuphe, N., Oguntibeju, O. O., & Nchu, F. (2021). Evaluating the Endophytic Activities of Beauveria bassiana on the Physiology, Growth, and Antioxidant Activities of Extracts of Lettuce (Lactuca sativa L.). Plants, 10(6), 1178. https://doi.org/10.3390/plants10061178