Abstract
In vegetables of Brassica rapa L., Fusarium oxysporum f. sp. rapae (For) or F. oxysporum f. sp. conglutinans (Foc) cause Fusarium yellows. A resistance gene against Foc (FocBr1) has been identified, and deletion of this gene results in susceptibility (focbr1-1). In contrast, a resistance gene against For has not been identified. Inoculation tests showed that lines resistant to Foc were also resistant to For, and lines susceptible to Foc were susceptible to For. However, prediction of disease resistance by a dominant DNA marker on FocBr1 (Bra012688m) was not associated with disease resistance of For in some komatsuna lines using an inoculation test. QTL-seq using four F2 populations derived from For susceptible and resistant lines showed one causative locus on chromosome A03, which covers FocBr1. Comparison of the amino acid sequence of FocBr1 between susceptible and resistant alleles (FocBr1 and FocBo1) showed that six amino acid differences were specific to susceptible lines. The presence and absence of FocBr1 is consistent with For resistance in F2 populations. These results indicate that FocBr1 is essential for For resistance, and changed amino acid sequences result in susceptibility to For. This susceptible allele is termed focbr1-2, and a new DNA marker (focbr1-2m) for detection of the focbr1-2 allele was developed.
1. Introduction
Brassica rapa L. comprises a variety of vegetables that are rich sources of nutrients including vitamins, minerals, dietary fiber, and phytochemicals [1,2]. In leafy vegetables of B. rapa, there are two morphotypes, heading types such as Chinese cabbage (var. pekinensis) and non-heading type such as pak choi (var. chinensis), komatsuna (var. perviridis) or chijimina (var. narinosa). Root vegetables such as turnip (var. rapa) also belong to B. rapa [1,2]. Most commercial cultivars of these vegetables are F1 hybrids, and hybrid vigor, disease resistance, and late bolting are important breeding traits [3,4,5]. In particular, disease resistance is demanded by farmers, especially for soil-borne diseases that are difficult to control with chemicals [5,6].
Plants have evolved their immunity to pathogens via two mechanisms [7,8]. Pattern recognition receptors (PRRs) located in the plant cell membrane recognize pathogen-associated molecular patterns (PAMPs), which activate PAMP-triggered immunity (PTI) and restrict pathogen development. Most pathogens secrete effectors (avirulence (AVR) proteins) into plant cells to suppress PTI, while plants have various resistance (R) genes, which mainly encode Toll/Interleukin-1 receptor (TIR) or coiled-coil (CC), nucleotide-binding site (NBS), and leucine-rich repeat (LRR) domains, to detect effectors. Recognition of effectors by R proteins induces effector-triggered immunity (ETI), and recognition of specific effectors by R proteins is termed “gene-for-gene resistance” [7].
Fusarium oxysporum is a soil-borne fungus and comprises 150 host-specific formae speciales. F. oxysporum causes yellows in a wide range of host plants [5,6]. In B. rapa vegetables, two formae speciales of F. oxysporum f. sp. conglutinans (Foc) and f. sp. rapae (For) have been identified as causing Fusarium yellows [9]. Foc was first reported as a causal agent of yellowing in cabbage (Brassica oleracea L. var. capitata) in 1913 [5,6] and causes Fusarium yellows not only in B. oleracea vegetables including cabbage or broccoli but also in B. rapa vegetables including turnip, komatsuna, and pak choi [9,10,11,12,13]. In contrast, For causes yellowing in B. rapa vegetables, but not in B. oleracea vegetables [9].
The Fusarium yellows R gene against Foc has been identified in B. rapa (FocBr1) and B. oleracea (FocBo1) [11,12,13,14]. FocBr1 and FocBo1 are orthologs and encode a TIR-NBS-LRR protein. In B. rapa, an approximately 35-kb deletion including FocBr1 results in susceptibility (focbr1-1) and there are no reports of other causative mutations for susceptibility [12,15]. In contrast, there are three different susceptible alleles of FocBo1 in B. oleracea (focbo1-1, focbo1-2 and focbo1-3), but a 35-kb deletion similar to that in Fusarium yellows susceptible lines of B. rapa has not been identified [11,13,16,17].
The Fusarium yellows R gene against For (ForBr1) has not been identified. In this study, we performed QTL-seq to isolate ForBr1. We developed a DNA marker that can identify the susceptible alleles of Fusarium yellows and tested this marker in cultivars of B. rapa vegetables.
2. Results
2.1. Screening of Lines for Resistance to F. oxysporum f. sp. Rapae
We have shown that FocBr1 (Bra012688) is a resistance gene to F. oxysporum f. sp. conglutinans (Foc), and deletion of this gene results in susceptibility to Foc [12]; this susceptible allele is termed focbr1-1 [14]. We made a DNA marker (Bra012688m) to detect the deletion of FocBr1 that is homozygous for the focbr1-1 allele [12,15]. In this study, we performed inoculation tests using F. oxysporum f. sp. rapae (For) for screening for resistant lines. Three Chinese cabbage (var. pekinensis), three turnip (var. rapa) and 22 komatsuna (var. perviridis) lines were tested of which 18 lines were resistant and 10 lines were susceptible (Table 1). We also inoculated these 28 lines with Foc, and resistance to For and Foc was identical (Table 1). We examined whether the results of the inoculation test were consistent with the prediction by FocBr1 DNA marker (Bra012688m). In all lines of Chinese cabbage and turnip, the prediction by the DNA marker was identical to the resistance determined by the inoculation test, while in seven of 22 komatsuna lines (“Zaoh”, YBCG-12, YBCG-13, YBCG-14, YBCG-15, YBCG-TC02, and YBCG-TC05) the DNA marker prediction was not consistent with the results of the inoculation test (Table 1). We tested an additional 15 lines of B. rapa; three lines (“Chijimikomatsuna”, “Tsunashima”, and “Hirose”) were not consistent between the DNA marker prediction and the results of the inoculation test using For (Table S1).

Table 1.
Assessment of Fusarium yellows resistance by inoculation test.
2.2. Identification of the Causative Region of Resistance for F. oxysporum f. sp. Rapae
We performed linkage analysis using three individual F2 populations derived from hybrids between For susceptible lines not containing the FocBr1 deletion and resistant lines. In the 200 plants of the F2 population derived from YBCG-11 (resistant) × YBCG-12 (susceptible) hybrid, 169 plants were resistant and 31 plants were susceptible to For. The number of susceptible plants was too small to be explained by a single gene dominance (chi-squared test, p < 0.05) (Table 2). This was also the case for the other two populations derived from YBCG-11 × YBCG-13 (susceptible) and YBCG-11 × YBCG-14 (susceptible) hybrids (Table 2). To identify the region covering the R gene for For (ForBr1), we performed QTL-seq analysis using bulked DNAs derived from about 20 resistant and susceptible individual plants derived from YBCG-11 × YBCG-12, YBCG-11 × YBCG-13, or YBCG-11 × YBCG-14 hybrids and found one similar locus on chromosome A03 in all three populations (Figure 1, Figures S1–S3). 22.0–33.5 Mb, 22.9–35.5 Mb, and 22.5–33.6 Mb region was detected as the QTL by 95% significance in the F2 population derived from YBCG-11 × YBCG-12, YBCG-11 × YBCG-13, and YBCG-11 × YBCG-14 hybrid, respectively (Figure 1). 1824, 1778, and 1734 genes were located in three QTLs, and 1655 genes overlapped (Figure S4, Table S2). A domain search using HMMSCAN with Pfam database and NCBI conserved domain search found nine genes encoding NBS-LRR proteins, including FocBr1 (BraA03g047240.3C or Bra012688) (Table S3).
Table 2.
Linkage analysis using six individual F2 populations.
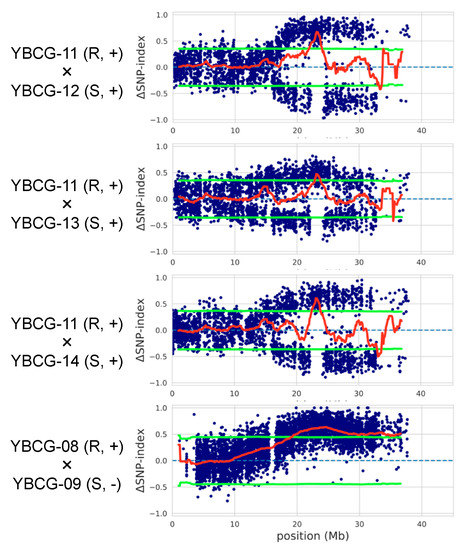
Figure 1.
QTL-seq results on chromosome A03. F2 populations derived from YBCG-11 × YBCG-12, YBCG-11 × YBCG-13, YBCG-11 × YBCG-14, and YBCG-08 × YBCG-09 hybrids were used. Blue dots indicate ∆SNP-index, and the red line indicates the sliding window average of ∆SNP-index. Light green lines represent p < 0.05. R and S represent resistant and susceptible, respectively. + and - represent the presence and absence of PCR amplification of Bra012688m marker, respectively.
2.3. A New Susceptible Allele of FocBr1 Was Identified
Because FocBr1 was included in three QTLs, we focused on FocBr1 for further analysis. The expression level of FocBr1 in three susceptible lines (“Zaoh”, YBCG-12, and YBCG-15) was similar to that of resistant lines (YBCG-11 and YBCG-16) (Figure 2), indicating that expression levels are not related to susceptibility. Next, we compared the amino acid sequences of FocBr1 in resistant and susceptible lines. The amino acid sequence of FocBr1 in the resistant line, YBCG-11, was 100% identical to FocBr1 in the resistant line, RJKB-T23 [12]. Amino acid sequences of FocBr1 were 100% identical among bulked DNAs of susceptible plants derived from F2 populations of YBCG-11 × YBCG-12, YBCG-11 × YBCG-13 and YBCG-11 × YBCG-14 hybrids, but there were some substitutions of amino acid sequences compared with FocBr1 in YBCG-11 (Figure 3). There were eleven amino acid sequence differences in FocBr1 between resistant and susceptible lines; five (A546T, N721D, T803K, V805E, and K862N) of which were identical between FocBr1 in the susceptible lines and the resistant allele of FocBo1 (B. oleracea) (Figure 3). Both FocBr1 and FocBo1 are resistance genes to Foc, indicating that the difference of amino acid sequences between FocBr1 and FocBo1 might not relate to the Fusarium yellows resistance, and the identical amino acid sequences between susceptible lines and FocBo1 might not lead to its susceptibility. The remaining six amino acid changes (Q859W, M869K, L1060F, V1148L, K1212T, and Q1395L) were differed between susceptible lines (YBCG-12, YBCG-13, and YBCG-14) and resistance lines of B. rapa and B. oleracea (Figure 3), which are susceptible line specific, suggesting that some of these amino acid sequences specific to susceptible lines result in susceptibility to For; some mutations may result in loss of function.
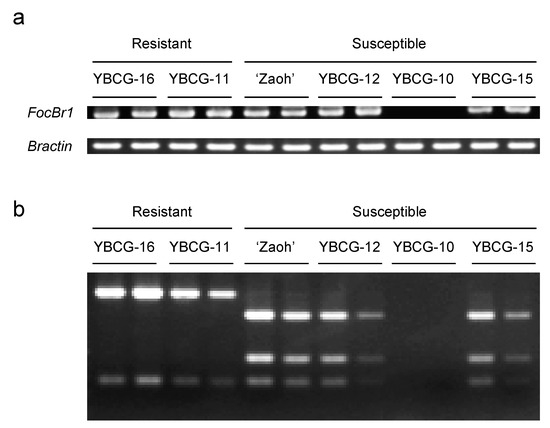
Figure 2.
Expression and genotype of FocBr1 in For resistant and susceptible lines. (a) Expression of FocBr1 and Bractin (control) was confirmed by RT-PCR. (b) DNA fragments of RT-PCR products digested by Hind III. YBCG-16 and YBCG-11 have FocBr1/FocBr1 homozygous or FocBr1/focbr1-1 heterozygous alleles. “Zaoh”, YBCG-12 and YBCG-15 have focbr1-2/focbr1-2 homozygous or focbr1-2/focbr1-1 heterozygous alleles, and YBCG-10 has focbr1-1/focbr1-1 homozygous allele.
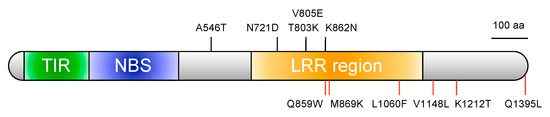
Figure 3.
Protein structure of FocBr1 in the resistant line of B. rapa. TIR (green box), NBS (blue box), and LRR region (orange box) were identified. Black lines represent the position of difference of amino acid sequences between resistant and susceptible lines in B. rapa, while amino acid sequences of FocBr1 in the susceptible lines were identical to the FocBo1 (Foc resistance gene in B. oleracea). Red lines represent the position of susceptible line-specific amino acid substitutions. Domains were predicted using HMMSCAN with Pfam database. (https://www.ebi.ac.uk/Tools/hmmer/, accessed on 1 April 2021) and NCBI conserved domain search (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi, accessed on 1 April 2021).
We examined whether FocBr1 deletion (focbr1-1) causes susceptibility to For. Linkage analysis using three individual F2 populations derived from hybrids between For susceptible lines (focbr1-1) and resistant lines (YBCG-08 (resistant) × YBCG-09 (susceptible), YBCG-TC01 (resistant) × YBCG-10 (susceptible), and YBCG-11 × YBCG-10) showed that the number of resistant and susceptible plants segregated as 3:1 ratio (chi-squared test, p > 0.05) (Table 2). QTL-seq analysis using bulked DNAs derived from about 20 resistant and susceptible plants of the F2 population derived from YBCG-08 × YBCG-09 hybrid found one causative locus (19.0–36.6 Mb) on chromosome A03, which covers the FocBr1 locus (Figure 1 and Figure S5). We tested a DNA marker (Bra012688m) in 12 resistant and susceptible plants from these three F2 populations, and the presence and absence of FocBr1 was consistent with the inoculation test (Figure S6). These results indicate that FocBr1 is essential for resistance to not only Foc but also For, supporting the suggestion that mutations cause susceptibility to Foc and For. This susceptible allele was termed focbr1-2.
2.4. Development of a New DNA Marker for Fusarium Yellows Resistance
Using sequence polymorphism between FocBr1 and focbr1-2, a new cleaved amplified polymorphic sequence (CAPS) DNA marker (focbr1-2m) was developed. Using this DNA marker, genotypes of 12 resistant and susceptible plants in three F2 populations derived from YBCG-11 × YBCG-12, YBCG-11 × YBCG-13, and YBCG-11 × YBCG-14 hybrids were confirmed, and genotypes were identical to the resistance determined by the inoculation test (Table S4). All ten lines that were not consistent between For inoculation test and the DNA marker (Bra012688m) prediction had homozygous focbr1-2 or heterozygous focbr1-2 and focbr1-1 alleles (Table 1 and Table 3, and Table S1). Another 33 B. rapa lines were consistent between the inoculation test using For and prediction by DNA marker (focbr1-2m) (Table 3). These results indicate that this new DNA marker can detect not only the focbr1-1 susceptible allele but also the focbr1-2 allele.
Table 3.
F. oxysporum f. sp. rapae resistance and FocBr1 genotype determined by focbr1-2m marker.
2.5. Prediction of Fusarium Yellows Resistance in Commercial B. rapa Vegetables by DNA Marker
Using the focbr1-2m marker, we predicted the resistance to For in 157 cultivars of Chinese cabbage, 35 cultivars of turnip, 40 cultivars of pak choi, and 73 cultivars of komatsuna. Of 157 cultivars of Chinese cabbage, six cultivars (3.8%) were heterozygous for FocBr1/focbr1-2, and there were no cultivars homozygous for focbr1-2/focbr1-2 or heterozygous for focbr1-2/focbr1-1 (Table 4). There were six Chinese cabbage cultivars (3.8%) homozygous for focbr1-1/focbr1-1 (Table 4), which could be susceptible to either For or Foc. Of 35 cultivars of turnip, five cultivars (14.3 %) were heterozygous for FocBr1/focbr1-2, and there were no cultivars homozygous for focbr1-2/focbr1-2, focbr1-1/focbr1-1, or heterozygous for focbr1-2/focbr1-1 (Table 4). Of 40 cultivars of pak choi, 16 cultivars (40.0%) were heterozygous for FocBr1/focbr1-2, and there were no cultivars homozygous for focbr1-2/focbr1-2, focbr1-1/focbr1-1, or heterozygous for focbr1-2/focbr1-1 (Table 4). Of 73 cultivars of komatsuna, 21 cultivars (28.8%) were heterozygous for FocBr1/focbr1-2, and five cultivars (6.8%) were homozygous for focbr1-2/focbr1-2 or heterozygous for focbr1-2/focbr1-1. There were three cultivars (4.1%) homozygous for focbr1-1/focbr1-1 (Table 4). Cultivars with focbr1-2/focbr1-2 or focbr1-2/focbr1-1 were found only in komatsuna.
Table 4.
Genotype distribution of FocBr1 in B. rapa subspecies.
3. Discussion
Previously, we identified a resistance gene to Foc (FocBr1), which is a single dominant gene. In susceptible lines, a 35 kb deletion, which includes FocBr1, was found [12], and the susceptible allele was termed focbr1-1 [14]. A dominant DNA marker (Bra012688m) has been made and the prediction of Foc resistance using Bra012688m was consistent with phenotypes of Foc resistance confirmed by an inoculation test in inbred lines of Chinese cabbage [12,15]. In this study, we inoculated Foc and other formae speciales, For, to Chinese cabbage, turnip, pak choi, komatsuna, and chijimina lines, and we found the prediction using Bra012688m was not consistent with phenotype using inoculation test in some lines, especially in komatsuna. To clarify the inheritance pattern of the R gene to For, we performed linkage analysis. Three F2 populations derived from crosses between For resistant and susceptible lines (focbr1-1) showed a 3:1 ratio of resistant to susceptible plants. In contrast, the other three F2 populations derived from crosses between For resistant and susceptible lines (focbr1-2) did not show a 3:1 ratio of resistant to susceptible plants; the number of susceptible plants of the F2 population is smaller than the expected number. However, QTL-seq using these populations identified one causative locus. This could be a difference in the detail of the loss of function; FocBr1 of focbr1-2 allele might have a weak function against For or be susceptible to environmental effects, although the focbr1-1 allele has completely lost its function. However, as both alleles showed a strong susceptible phenotype and we could not identify any significant difference between these two alleles in B. rapa lines, further analysis will be needed to identify this minor difference between focbr1-1 and focbr1-2 alleles.
QTL-seq analysis using F2 populations derived from crosses between For resistant and susceptible lines with the FocBr1 deletion (focbr1-1) or without the FocBr1 deletion (focbr1-2) identified the same single causative locus for For resistance, which covered FocBr1. There was no difference in expression levels of FocBr1 between resistant and susceptible lines, but there were some amino acid sequence differences between For susceptible allele (focbr1-2) and resistant alleles (FocBr1 and FocBo1), suggesting that changes of amino acid sequence result in loss of function. Some substitutions were in the LRR region, and these susceptible line-specific amino acid changes may lead to loss of recognition of AVR. This new allele might be useful for identifying the sequence that is important for the interaction between R and AVR proteins. To prove this amino acid sequences change results in loss of function, which might be due to loss of recognition to AVR, further experiments such as making transgenic plants for complementation or loss of function by CRISPR-Cas9 system will be required.
Alternatively, another gene(s) linked to the FocBr1 locus may work together with FocBr1 for For resistance, because the peak detected by QTL-seq is upstream from the FocBr1 position in three F2 populations derived from crosses between For resistant and susceptible lines (focbr1-2). In Arabidopsis thaliana, TIR-NBS-LRR type Resistance to Ralstonia solanacearum 1 (RRS1) and Resistance to Pseudomonas syringae 4 (RPS4) are neighboring genes and both are required for resistance to Colletotrichum higginsianum, Ralstonia solanacearum, and Pseudomonas syringae pv. tomato strain DC3000 expressing avrRps4 [18]. RRS1 encodes a WRKY domain protein as well as a TIR-NBS-LRR protein and works as a “sensor” to detect the effector, and RPS4 works as a “helper” to activate cell death [19,20]. If a similar function is applied to Fusarium yellows resistance, FocBr1 will work as a “sensor” NBS-LRR with other “helper” gene(s), which may be located on a region upstream from FocBr1. Resistant and susceptible alleles of “sensor”, FocBr1 and focbr1-2, might be able to recognize AVR to greater or lesser degrees, respectively, and other “helper” gene(s) might have different functions between resistant and susceptible lines, resulting in a shift of QTL peak to upstream. Further analyses using plants recombined between QTL peak locus and FocBr1 gene are required to clarify whether other factor(s) are important for For resistance and the For infection mechanisms. There is also another possibility that minor QTLs not linked to FocBr1 locus are important for resistance to For.
Using a new DNA marker, focbr1-2m, we screened genotypes of B. rapa breeding lines and cultivars. There were six lines that showed weak resistance against For and Foc, and three of the six lines (“CR-Taiga”, “Natsurakuten”, and YBCG-TC04) showed heterozygosity of FocBr1 and focbr1-2. However, in the remaining three lines (“CR-Yukiakari”, NSI-01, and YBCG-18), we cannot distinguish between the homozygosity of FocBr1 and the heterozygosity of FocBr1 and focbr1-1, because focbr1-1 results in deletion of FocBr1 and focbr1-2m cannot amplify focbr1-1 allele. In B. rapa, plants having a homozygous clubroot resistance gene show more stable clubroot resistance than plants having a heterozygous resistance gene [21,22,23]. Three lines may be heterozygous for FocBr1 and focbr1-1. In the case of focbr1-1, it is desirable to develop a DNA marker to distinguish between FocBr1/focbr1-1 heterozygosity and FocBr1/FocBr1 homozygosity, i.e., using a linked marker close to the 35-kb deletion [14]. In the case of focbr1-2, codominant DNA marker, focbr1-2m, can distinguish the heterozygosity of FocBr1 and focbr1-2 alleles, which will be useful for breeding stable Fusarium yellows resistant cultivars.
In Chinese cabbage cultivars, there were no cultivars homozygous for focbr1-2, and a few lines heterozygous for FocBr1/focbr1-2. In our previous study using the Bra012688m marker, there was complete agreement between the DNA marker-based prediction and the inoculation test in Chinese cabbage lines [15]. Thus, there is little risk that the presence of the focbr1-2 allele leads to susceptibility during the breeding of Fusarium yellows resistant cultivars in Chinese cabbage. Like in Chinese cabbage, most turnip cultivars (about 85%) did not have focbr1-2 alleles, so this allele will not be a problem for breeding. However, in pak choi, 40% of cultivars were heterozygous for FocBr1 and focbr1-2 alleles. In komatsuna, about 30% of cultivars were heterozygous for FocBr1 and focbr1-2 alleles and about 7% of cultivars were homozygous for focbr1-2 allele or heterozygous for focbr1-2 and focbr1-1 alleles. For the breeding of Fusarium yellows resistant cultivars in pak choi or komatsuna, the presence of the focbr1-2 allele should be mapped in breeding lines, and the DNA marker, focbr1-2m, developed in this study will be useful for DNA marker-assisted selection.
4. Materials and Methods
4.1. Plant Materials and DNA and RNA Extraction
The breeding lines and commercial F1 hybrid cultivars of B. rapa vegetables were used as plant materials (Table S5). F2 populations were produced by bud pollination of F1 hybrid crossing YBCG-11 × YBCG-12, YBCG-11 × YBCG-13, YBCG-11 × YBCG-14, YBCG-08 × YBCG-09, YBCG-TC01 × YBCG-10, and YBCG-11 × YBCG-10. Genomic DNA was isolated from leaves by the CTAB (cetyl trimethyl ammonium bromide) method [24]. Total RNA was isolated from noninoculated leaves of ten-days-old seedlings by the SV Total RNA Isolation System (Promega Co., Madison, WI, USA).
4.2. Inoculation Test
A strain of F. oxysporum f. sp. rapae (isolated from komatsuna) (provided by NARO, MAFF 240322) or F. oxysporum f. sp. conglutinans (isolated from cabbage) was used to prepare inocula. Liquid inocula were obtained by inoculating potato sucrose broth medium (200 g/L potato extract and 20 g/L sucrose in distilled water) with the isolate and shaking at 130 rpm on a rotary shaker for 1 week. Roots of ten-days-old seedlings were dipped in fungal spore suspension (fungal titer of ~5 × 106) for 5 h and then transplanted into a cell tray filled with soil. Plants were grown in the greenhouse, and two or three weeks after inoculation, individual plants were scored for interaction phenotype (IP) based on six categories that are 0 (no symptoms in tops and roots), 3 (darkening of roots, slight top stunting, and no chlorosis), 5 (dark stunted roots, tops stunted, and slight chlorosis of cotyledons), 7 (severe stunting of roots and tops and strong chlorosis) and 9 (severe stunting, necrosis, and death). To show the phenotype of breeding lines and cultivars, the average IP among 25 seedlings were categorized into resistant (IP = 0) or susceptible (IP = 3–9). Average IP of most resistant lines/cultivars was around 0 and average IP of most susceptible lines/cultivars was 9. However, some of the 25 seedlings of the line showed IP = 0 while others showed IP ≥ 3, and these exceptions are represented by R*. In the linkage analysis, F2 seedlings were used for inoculation test, and phenotypes of individual seedlings were resistant (IP = 0) or susceptible (IP = 3–9). Chinese cabbage inbred lines RJKB-T23 and RJKB-T24 were used as a resistant and susceptible control [12].
4.3. QTL-Seq
QTL-seq was performed following the method described in [25]. From F2 populations derived from YBCG-11 × YBCG-12, YBCG-11 × YBCG-13, YBCG-11 × YBCG-14, and YBCG-08 × YBCG-09 hybrids, about 20 plants were selected from resistant (IP = 0) and susceptible (IP = 9) plants based on their perfectly resistance or susceptible phenotype. The equal amount of DNA from each sample was bulked by resistant and susceptible phenotypes, and named R-bulk and S-bulk, respectively. Eight sequence libraries were prepared for DNA sequencing using TruSeq DNA PCR-Free kit (Illumina, Inc., San Diego, CA, USA), and sequenced by Illumina Hiseq 4000 (paired end, 150 bp). For detecting the parental SNPs, DNA from parental line (YBCG-08) was also sequenced.
Sequence reads were quality trimmed by FaQCs. Trimmed reads of R-bulk and S-bulk were aligned to the B. rapa reference genome version 3.0 (https://brassicadb.cn, accessed on 1 April 2021), and SNP-index was calculated at all SNPs in R-bulk and S-bulk compared with resistant parental sequences, then the subtracted value of SNP-index of R-bulk from SNP-index of S-bulk was calculated as ∆SNP-index using QTL-seq pipeline.
4.4. Prediction of Fusarium Yellows Resistance by DNA Markers
To predict the Fusarium yellows resistance, the dominant marker Bra012688m [15] and codominant CAPS marker, focbr1-2m, were used. PCR was performed using QuickTaq®HS DyeMix (TOYOBO Co., Ltd., Osaka, Japan). The reaction mixture was incubated in the thermal cycler (TaKaRa PCR Thermal Cycler Dice® Gradient, Takara Bio Inc., Kusatsu, Japan) at 94 °C for 2 min following by 35 cycles of 94 °C for 30 s, 55 °C for 30 s, and 68 °C for 1 min. PCR products were detected by electrophoresis (i-MyRunII, COSMO BIO CO., LTD., Tokyo, Japan) using 1.0% agarose gel (Bra012688m). To distinguish the FocBr1 and focbr1-2 alleles, amplified DNA digested by Hind III restriction enzyme were electrophoresed on 1.5% agarose gel. Two or more independent individual plants in each cultivar were tested for genotyping. The primer sets of the DNA markers are listed in Table S6.
4.5. Gene Expression Analysis
cDNA was synthesized from 500 ng total RNA using ReverTra Ace qPCR RT Master Mix with gDNA Remover (TOYOBO Co., Ltd.). The specificity of the primer set of FocBr1 was first tested by electrophoresis of RT-PCR amplified products using QuickTaq®HS DyeMix on 1.5% agarose gel in which a single product was observed. RT-PCR conditions were 94 °C for 2 min followed by 35 cycles of 94 °C for 30 s, 55 °C for 30 s, and 68 °C for 30 s. The absence of genomic DNA contamination was confirmed by the PCR of the no RT control. To distinguish the FocBr1 and focbr1-2 alleles, amplified DNA by RT-PCR digested by Hind III restriction enzyme were electrophoresed on 1.5% agarose gel. The primer sets for RT-PCR are listed in Table S6.
5. Conclusions
In this study, we identified FocBr1 as a For resistance gene, and a new susceptible allele of FocBr1, focbr1-2, was identified in B. rapa. Furthermore, a new DNA marker, which can distinguish between FocBr1, focbr1-1, and focbr1-2, was developed.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/plants10061082/s1, Figure S1: QTL-seq results of all chromosomes of F2 populations derived from YBCG-11 × YBCG-12 hybrid, Figure S2: QTL-seq results of all chromosomes of F2 populations derived from YBCG-11 × YBCG-13 hybrid, Figure S3: QTL-seq results of all chromosomes of F2 populations derived from YBCG-11 × YBCG-14 hybrid, Figure S4: Overlapped candidate genes in three F2 populations, Figure S5: QTL-seq results of all chromosomes of F2 populations derived from YBCG-08 × YBCG-09 hybrid, Table S1: Assessment of Fusarium yellows resistance by inoculation test, Table S2: Genes located on candidate locus detected by QTL-seq in four populations by 95% significance, Table S3: NBS-LRR encoded genes in significant QTLs, Table S4: Genotype using focbr1-2m marker in resistant and susceptible plants derived from F2 population, Table S5: List of cultivars used in this study, Table S6: List of primer sequences.
Author Contributions
Conceptualization, N.M., C.S., T.I., K.O., M.S., M.K. (Makoto Kaji), M.K. (Masahiko Katsumata), and R.F.; methodology, validation and formal analysis, N.M., M.A.A., C.S., H.M., T.S., T.I., M.S. and R.F.; writing—original draft preparation, N.M., M.A.A., M.S., E.S.D. and R.F.; writing—review and editing, N.M., E.S.D. and R.F.; visualization, N.M. and M.S.; supervision, M.K. (Masahiko Katsumata) and R.F.; funding acquisition, T.I., M.K. (Makoto Kaji), M.K. (Masahiko Katsumata) and R.F. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by grants from the Project of the NARO Bio-oriented Technology Research Advancement Institution (Research program on Development of Innovation Technology) (30029C).
Acknowledgments
We are grateful to Yumiko Arai for her technical assistance throughout this project. We thank Kenji Osabe for his helpful comments and manuscript editing.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cheng, F.; Sun, R.; Hou, X.; Zheng, H.; Zhang, F.; Zhang, Y.; Liu, B.; Liang, J.; Zhuang, M.; Liu, Y.; et al. Subgenome parallel selection is associated with morphotype diversification and convergent crop domestication in Brassica rapa and Brassica oleracea. Nat. Genet. 2016, 48, 1218–1224. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Miyaji, N.; Osabe, K.; Akter, A.; Mehraj, H.; Shea, D.J.; Fujimoto, R. The importance of genetic and epigenetic research in the Brassica vegetables in the face of climate change. In Genomic Designing of Climate-Smart Vegetable Crops; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 161–255. [Google Scholar]
- Fujimoto, R.; Uezono, K.; Ishikura, S.; Osabe, K.; Peacock, W.J.; Dennis, E.S. Recent research on the mechanism of heterosis is important for crop and vegetable breeding systems. Breed. Sci. 2018, 68, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Akter, A.; Itabashi, E.; Kakizaki, T.; Okazaki, K.; Dennis, E.S.; Fujimoto, R. Genome triplication leads to transcriptional divergence of FLOWERING LOCUS C genes during vernalization in the genus Brassica. Front. Plant Sci. 2021, 11, 619417. [Google Scholar] [CrossRef] [PubMed]
- Mehraj, H.; Akter, A.; Miyaji, N.; Miyazaki, J.; Shea, D.J.; Fujimoto, R.; Doullah, M.A.U. Genetics of clubroot and Fusarium wilt disease resistance in Brassica vegetables: The application of marker assisted breeding for disease resistance. Plants 2020, 9, 726. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Y. An update on the arsenal: Mining resistance genes for disease management of Brassica crops in the genomic era. Hortic. Res. 2020, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Dodds, P.N.; Rathjen, J.P. Plant immunity: Towards an integrated view of plant-pathogen interactions. Nat. Rev. Genet. 2010, 11, 539–548. [Google Scholar] [CrossRef]
- Enya, J.; Togawa, M.; Takeuchi, T.; Yoshida, S.; Tsushima, S.; Arie, T.; Sakai, T. Biological and phylogenetic characterization of Fusarium oxysporum complex, which causes yellows on Brassica spp., and proposal of F. oxysporum f. sp. rapae, a novel forma specialis pathogenic on B. rapa in Japan. Phytopathology 2008, 98, 475–483. [Google Scholar] [CrossRef]
- Pu, Z.; Shimizu, M.; Zhang, Y.; Nagaoka, T.; Hayashi, T.; Hori, H.; Matsumoto, S.; Fujimoto, R.; Okazaki, K. Genetic mapping of a fusarium wilt resistance gene in Brassica oleracea. Mol. Breed. 2012, 30, 809–818. [Google Scholar] [CrossRef]
- Lv, H.; Fang, Z.; Yang, L.; Zhang, Y.; Wang, Q.; Liu, Y.; Zhuang, M.; Yang, Y.; Xie, B.; Liu, B.; et al. Mapping and analysis of a novel candidate Fusarium wilt resistance gene FOC1 in Brassica oleracea. BMC Genom. 2014, 15, 1094. [Google Scholar] [CrossRef]
- Shimizu, M.; Fujimoto, R.; Ying, H.; Pu, Z.; Ebe, Y.; Kawanabe, T.; Saeki, N.; Taylor, J.M.; Kaji, M.; Dennis, E.S.; et al. Identification of candidate genes for fusarium yellows resistance in Chinese cabbage by differential expression analysis. Plant Mol. Biol. 2014, 85, 247–257. [Google Scholar] [CrossRef]
- Shimizu, M.; Pu, Z.; Kawanabe, T.; Kitashiba, H.; Matsumoto, S.; Ebe, Y.; Sano, M.; Funaki, T.; Fukai, E.; Fujimoto, R.; et al. Map-based cloning of a candidate gene conferring Fusarium yellows resistance in Brassica oleracea. Theor. Appl. Genet. 2015, 128, 119–130. [Google Scholar] [CrossRef]
- Akter, M.A.; Mehraj, H.; Itabashi, T.; Shindo, T.; Osaka, M.; Akter, A.; Miyaji, N.; Chiba, N.; Miyazaki, J.; Fujimoto, R. Breeding for disease resistance in Brassica vegetables using DNA marker selection. In Brassica Breeding and Biotechnology; IntechOpen: London, UK, 2021. [Google Scholar] [CrossRef]
- Kawamura, K.; Kawanabe, T.; Shimizu, M.; Okazaki, K.; Kaji, M.; Dennis, E.S.; Osabe, K.; Fujimoto, R. Genetic characterization of inbred lines of Chinese cabbage by DNA markers: Towards the application of DNA markers to breeding of F1 hybrid cultivars. Data Brief 2016, 6, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, K.; Shimizu, M.; Kawanabe, T.; Pu, Z.; Kodama, T.; Kaji, M.; Osabe, K.; Fujimoto, R.; Okazaki, K. Assessment of DNA markers for seed contamination testing and selection of disease resistance in cabbage. Euphytica 2017, 213, 28. [Google Scholar] [CrossRef]
- Sato, M.; Shimizu, M.; Shea, D.J.; Hoque, M.; Kawanabe, T.; Miyaji, N.; Fujimoto, R.; Fukai, E.; Okazaki, K. Allele specific DNA marker for fusarium resistance gene FocBo1 in Brassica oleracea. Breed. Sci. 2019, 69, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Narusaka, M.; Shirasu, K.; Noutoshi, Y.; Kubo, Y.; Shiraishi, T.; Iwabuchi, M.; Narusaka, Y. RRS1 and RPS4 provide a dual Resistance-gene system against fungal and bacterial pathogens. Plant J. 2009, 60, 218–226. [Google Scholar] [CrossRef]
- Huh, S.U.; Cevik, V.; Ding, P.; Duxbury, Z.; Ma, Y.; Tomlinson, L.; Sarris, P.F.; Jones, J.D.G. Protein-protein interactions in the RPS4/RRS1 immune receptor complex. PLoS Pathog. 2017, 13, e1006376. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, H.; Hu, L.; Martinez, P.P.; Moschou, P.N.; Cevik, V.; Ding, P.; Duxbury, Z.; Sarris, P.F.; Jones, J.D.G. Distinct modes of derepression of an Arabidopsis immune receptor complex by two different bacterial effectors. Proc. Natl. Acad. Sci. USA 2018, 115, 10218–10227. [Google Scholar] [CrossRef]
- Suwabe, K.; Tsukazaki, H.; Iketani, H.; Hatakeyama, K.; Fujimura, M.; Nunome, T.; Fukuoka, H.; Matsumoto, S.; Hirai, M. Identification of two loci for resistance to clubroot (Plasmodiophora brassicae Woronin) in Brassica rapa L. Theor. Appl. Genet. 2003, 107, 997–1002. [Google Scholar] [CrossRef]
- Nomura, K.; Minegishi, Y.; Kimizuka-Takagi, C.; Fujioka, T.; Moriguchi, K.; Shishido, R.; Ikehashi, H. Evaluation of F2 and F3 plants introgressed with QTLs for clubroot resistance in cabbage developed by using SCAR markers. Plant Breed. 2005, 124, 371–375. [Google Scholar] [CrossRef]
- Nagaoka, T.; Doullah, M.A.U.; Matsumoto, S.; Kawasaki, S.; Ishikawa, T.; Hori, H.; Okazaki, K. Identification of QTLs that control clubroot resistance in Brassica oleracea and comparative analysis of clubroot resistance genes between B. rapa and B. oleracea. Theor. Appl. Genet. 2010, 120, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed]
- Takagi, H.; Abe, A.; Yoshida, K.; Kosugi, S.; Natsume, S.; Mitsuoka, C.; Uemura, A.; Utsushi, H.; Tamiru, M.; Takuno, S.; et al. QTL-seq: Rapid mapping of quantitative trait loci in rice by whole genome resequencing of DNA from two bulked populations. Plant J. 2013, 74, 174–183. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).