
High Throughput Profiling of Flavonoid Abundance in Agave lechuguilla Residue-Valorizing under Explored Mexican Plant



 , , ,
, , ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Agro-Residue Collection
2.2. Storage Conditions
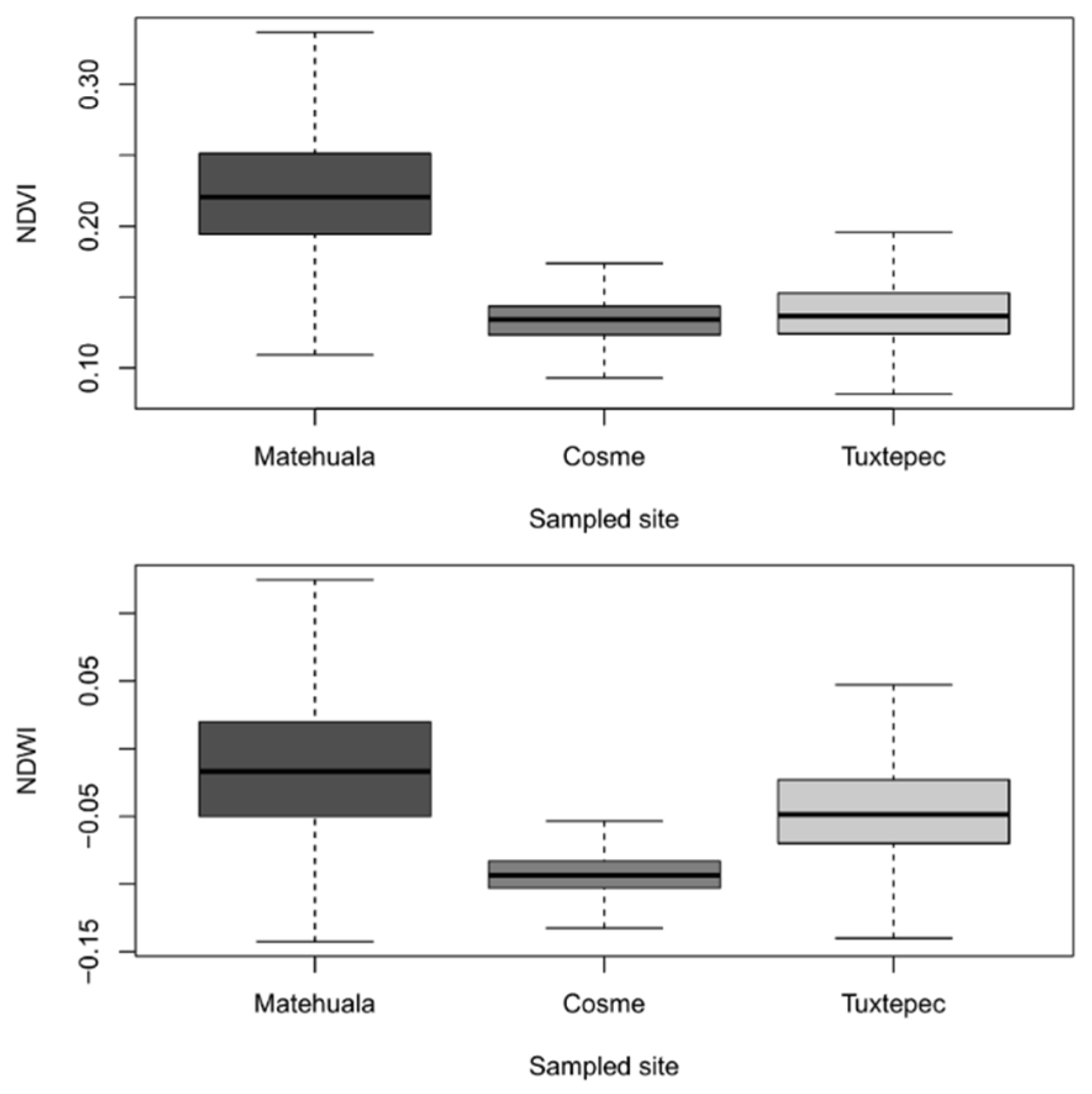
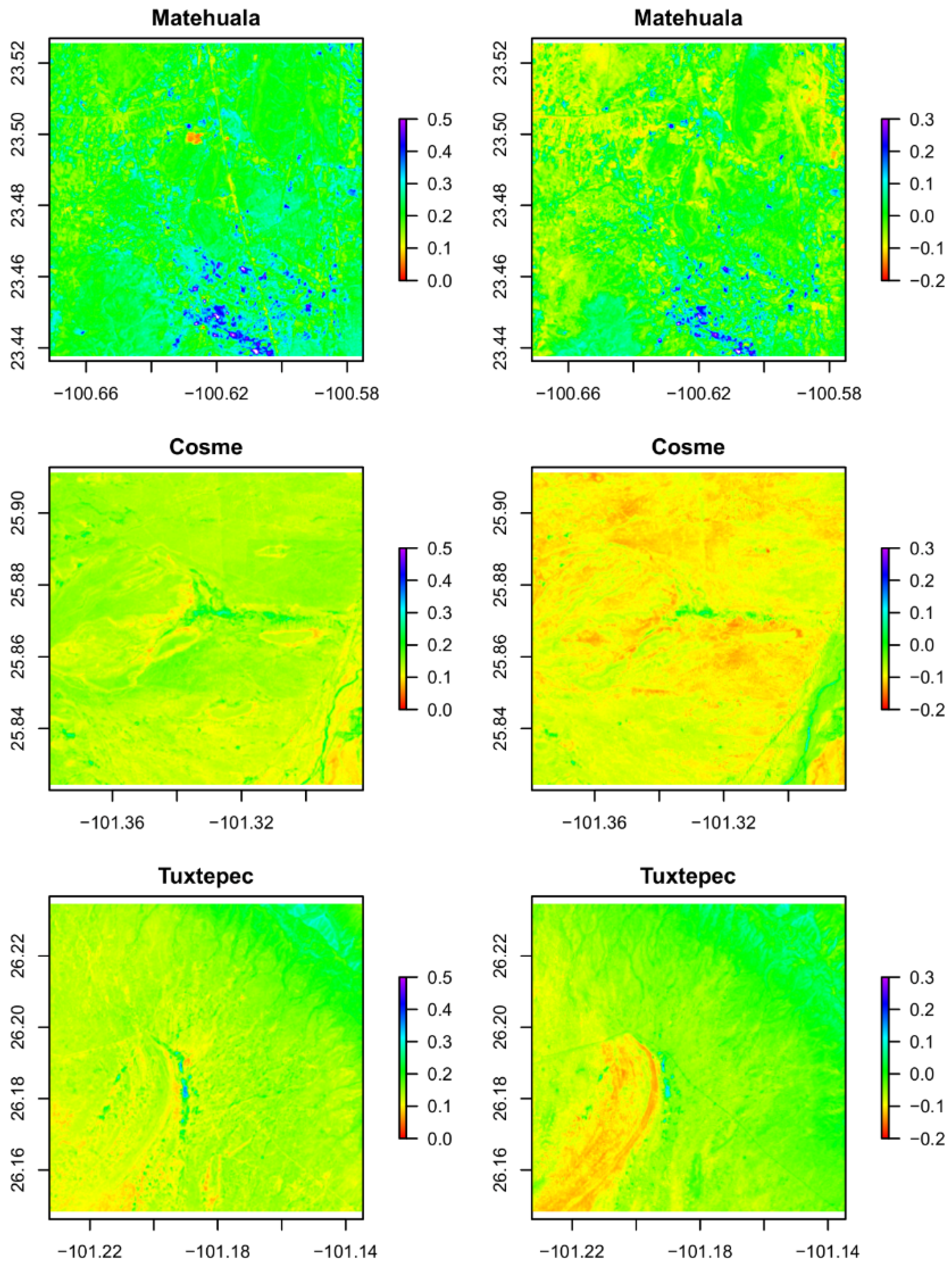
2.3. Environmental Characterization from Remote Sensing
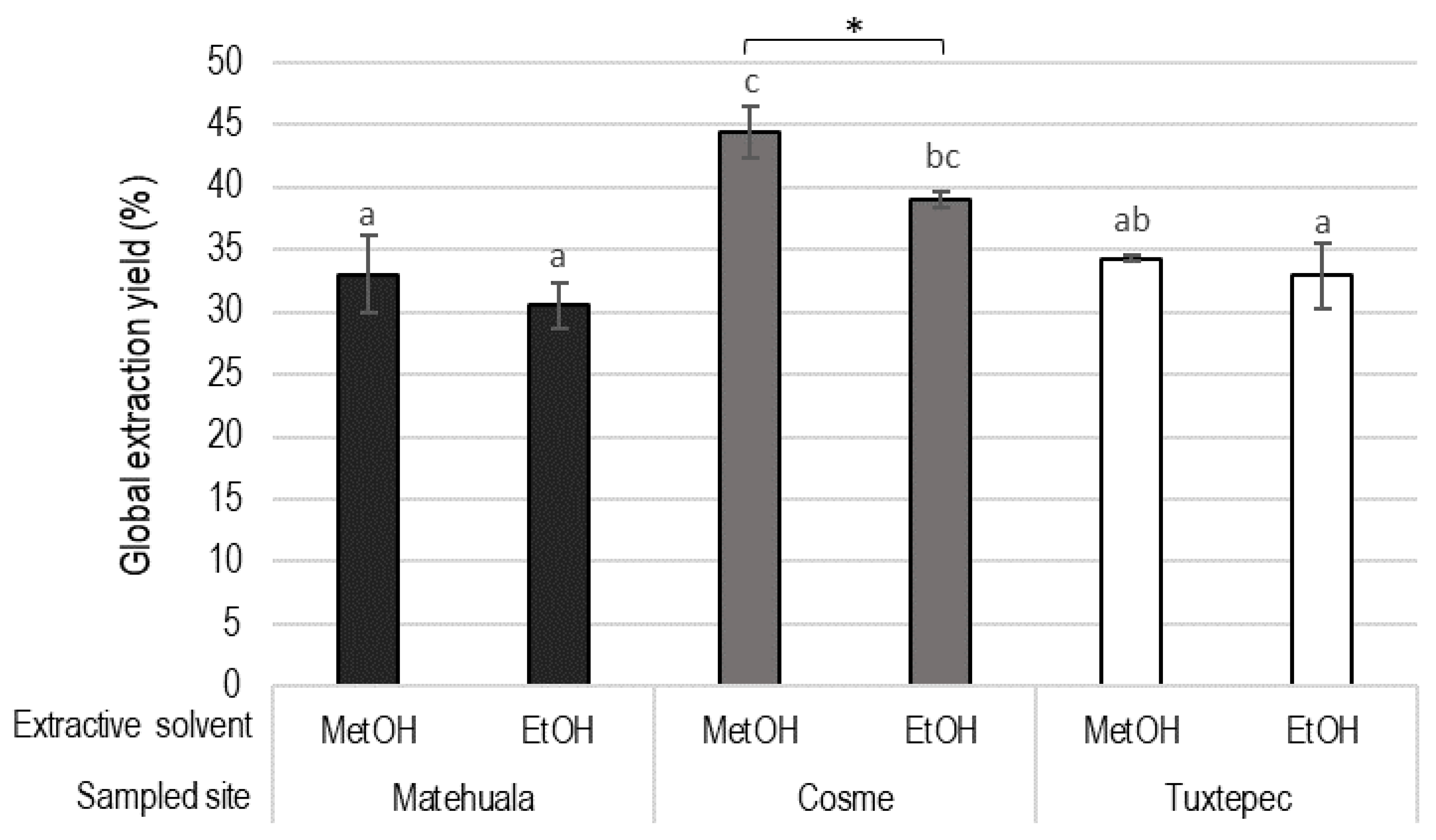
2.4. Phytochemical Extraction
2.5. Chemical Characterization
2.5.1. Stock Solutions of Extracted Phytochemicals
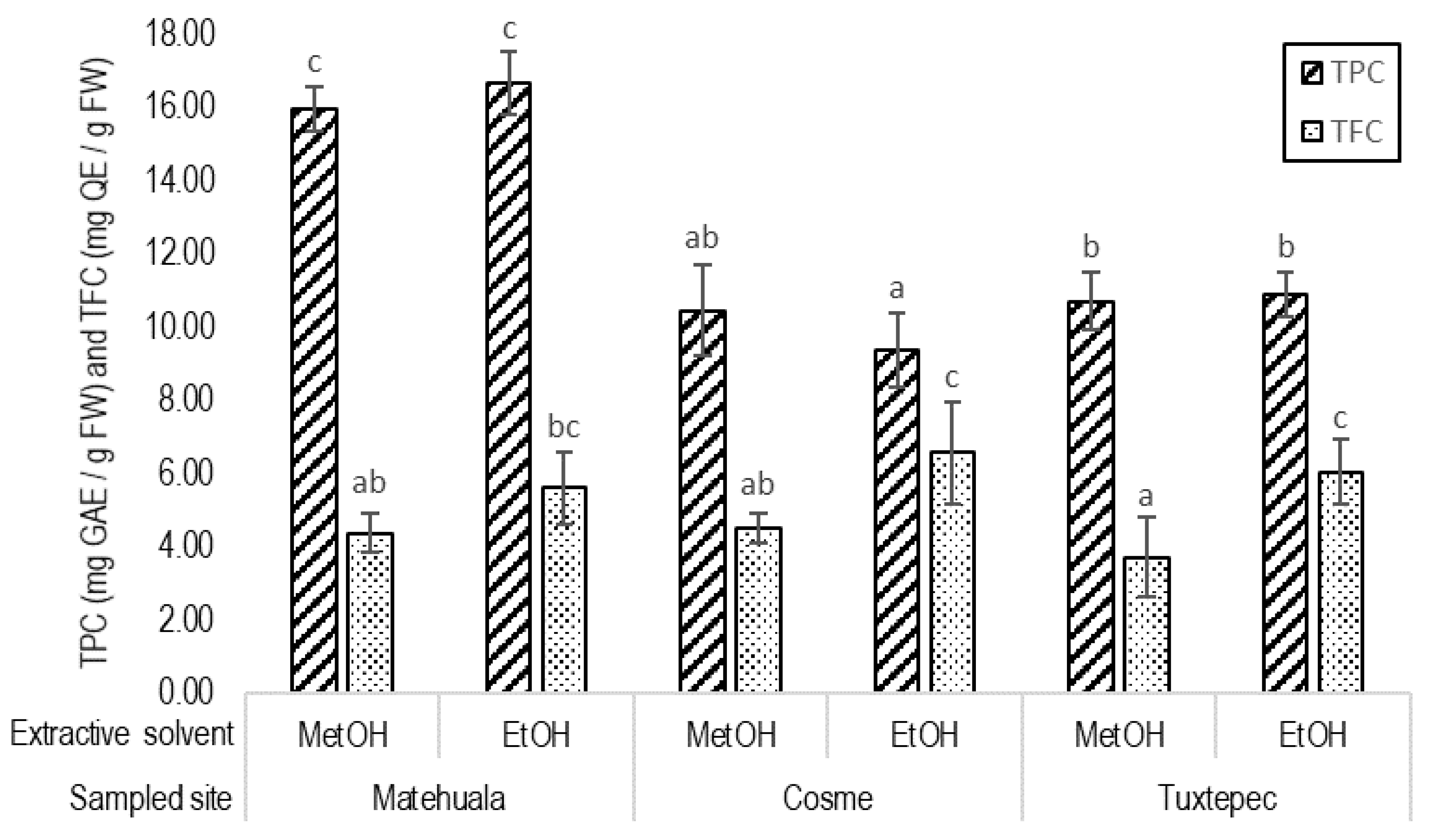
2.5.2. Total Polyphenolic Content (TPC)
2.5.3. Total Flavonoid Content (TFC)
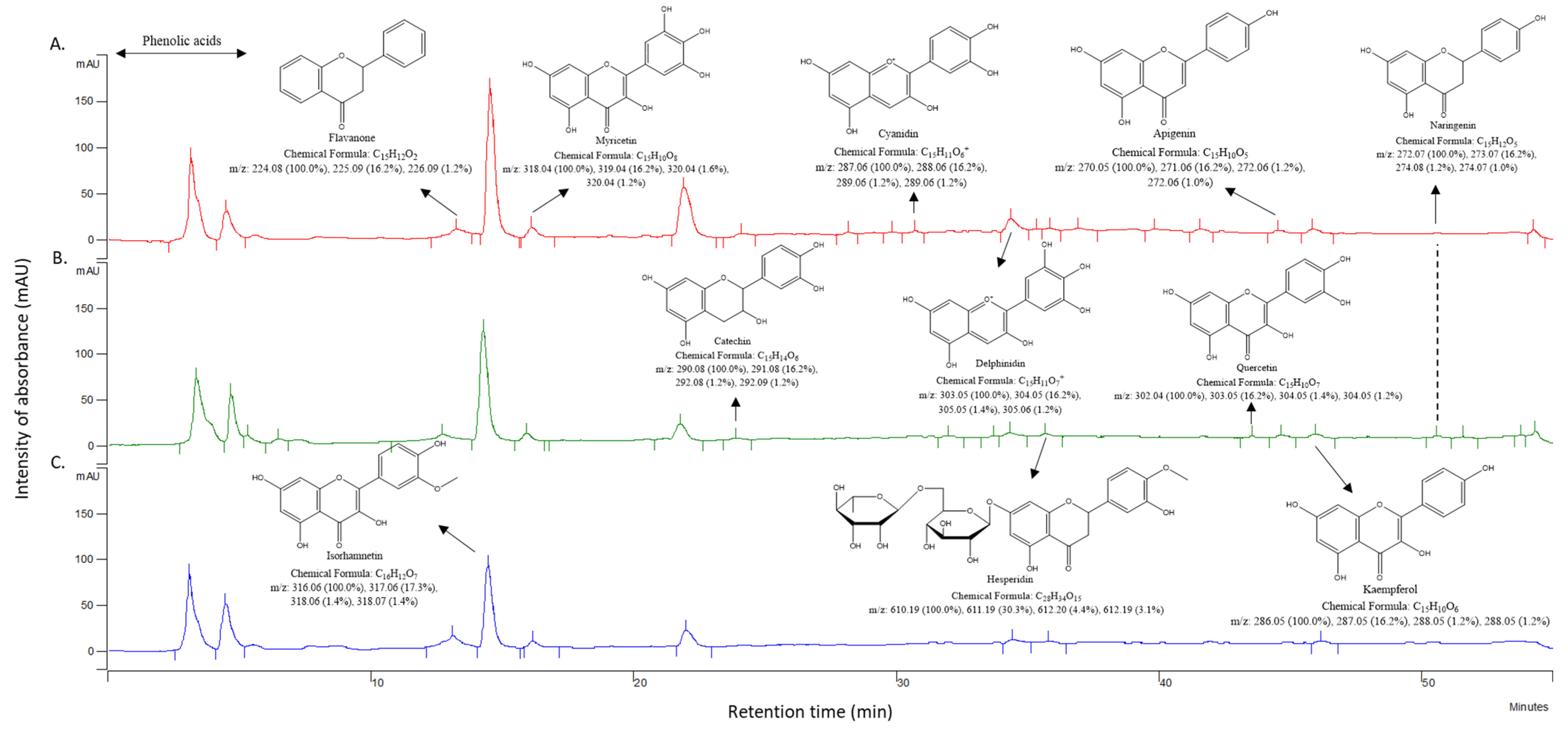
2.5.4. HPLC-UV-MS/MS
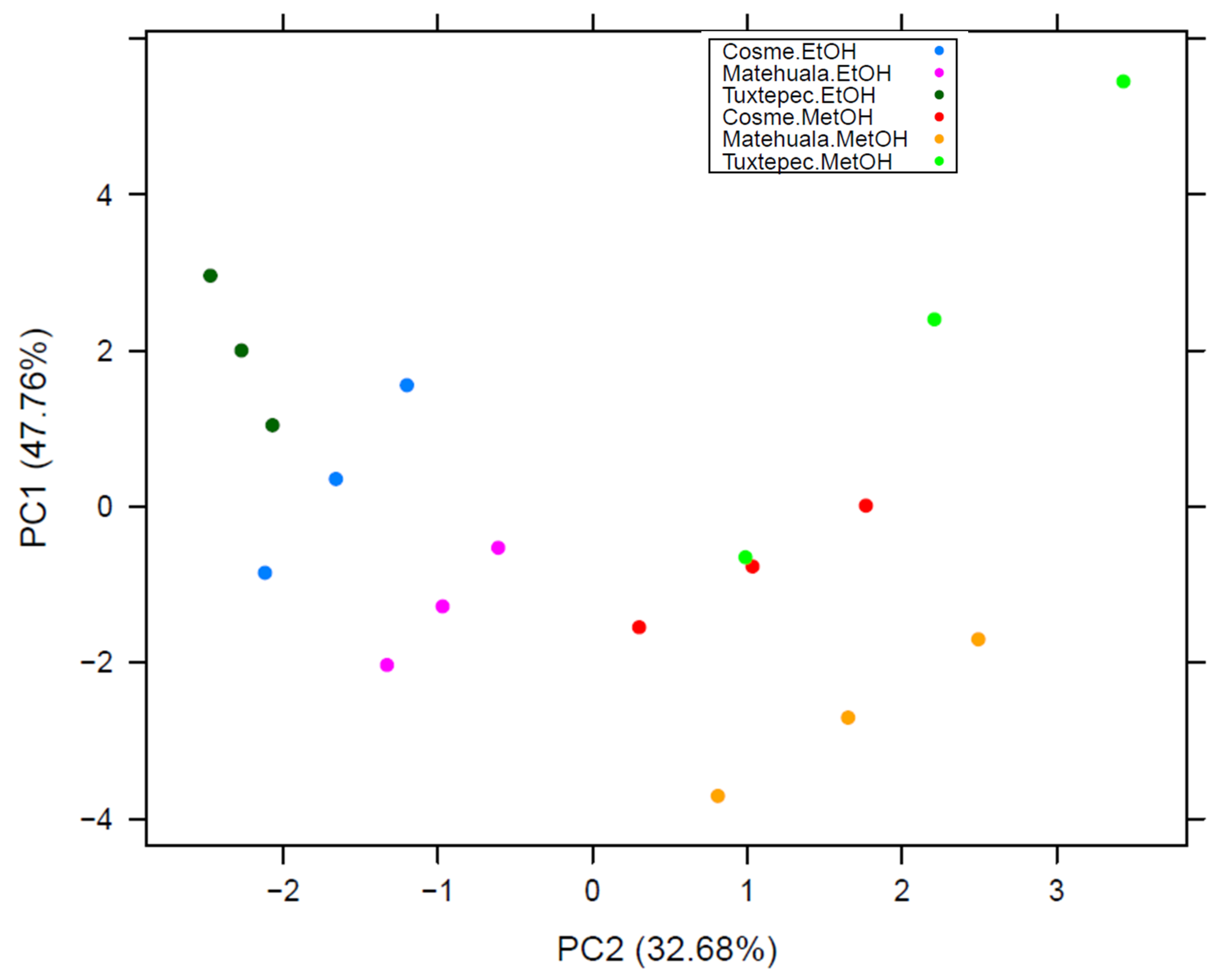
2.6. Statistical Analysis
3. Results
3.1. Geo-Climatic Conditions
3.2. Total Polyphenolic and Flavonoid Contents
3.3. Flavonoid Profiles
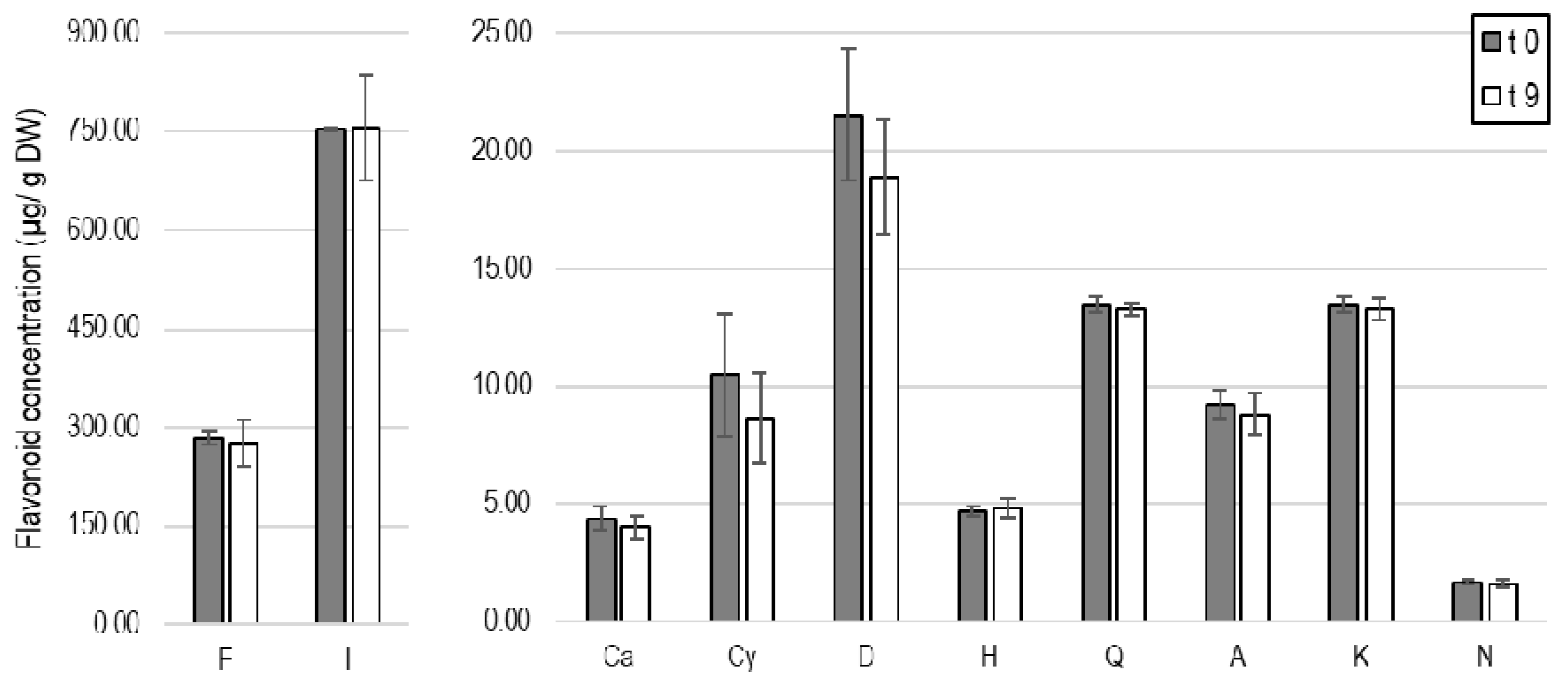
3.4. Conservation of Bioactive Flavonoids
4. Discussion
4.1. Geo-Climatic Variation
4.2. Agronomical and Biotechnological Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gentry, H.S. Agaves of Continental North America; University of Arizona Press: Tucson, AZ, USA, 1982; ISBN 978-0816523955. [Google Scholar]
- Díaz-Jiménez, L.; Carlos-Hernández, S.; Jasso de Rodríguez, D.; Rodríguez-García, R. Conceptualization of a biorefinery for guishe revalorization. Ind. Crops. Prod. 2019, 138, 111441. [Google Scholar] [CrossRef]
- Burciaga, M.O.U.M.; Quiroz, M.D.C.; Arreola, I.O.M.; Saltillo, I.D.C.E. Caracterización y Selección de Sitios Para Plantaciones de Lechuguilla (Agave Lechuguilla Torr.) en el Estado de Coahuila. Available online: http://www.inifapcirne.gob.mx/Biblioteca/Publicaciones/886.pdf (accessed on 22 February 2021).
- Pando-Moreno, M.; Pulido, R.; Castillo, D.; Jurado, E.; Jiménez, J. Estimating fiber for lechuguilla (Agave lecheguilla Torr., Agavaceae), a traditional non-timber forest product in Mexico. For. Ecol. Manag. 2008, 255, 3686–3690. [Google Scholar] [CrossRef]
- Castillo-Quiroz, D.; Sáenz-Reyes, J.; Narcia-Velasco, M.; Vásquez-Ramos, J. Physical and mechanical properties of Agave lechuguilla Torr. Fiber under plantation of five provenances. Rev. Mex. Cienc. For. 2013, 4, 78–91. [Google Scholar] [CrossRef][Green Version]
- Consejo Nacional de Población (CONAPO). Índice de Marginación Por Localidad. 2010. Available online: http://www.conapo.gob.mx/es/CONAPO/indice_de_marginacion_por_localidad_2010 (accessed on 22 February 2021).
- Consejo Nacional de Evaluación de la Política de Desarrollo Social (CONEVAL). Medición de la Pobreza. 2015. Available online: https://www.coneval.org.mx/Medicion/Paginas/POBREZA-URBANA-EN-MEXICO-2015.aspx (accessed on 22 February 2021).
- Reyes-Agüero, J.A.; Aguirre-Rivera, J.R.; Peña-Valdivia, C.B. Biología y Aprovechamiento de Agave lechuguilla Torrey. Bol. Soc. Bot. Mex. 2000, 67, 75–88. [Google Scholar] [CrossRef][Green Version]
- Castillo-Quiroz, D.; Cano-Pineda, A.; Berlanga-Reyes, C.A. Establecimiento y Aprovechamiento de Lechuguilla (Agave lechuguilla Torr.). Comisión Nacional Forestal-Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. 2012. Available online: http://www.conafor.gob.mx/biblioteca/Establecimiento_y_aprovechamiento_de_la_lechuguilla_Agave%20_lechuguilla_Torr.pdf (accessed on 22 February 2021).
- Secretaria de Medio Ambiente y Recursos Naturales (SEMARNAT). Norma Oficial Mexicana NOM-008-SEMARNAT-1996. Procedimientos, Criterios y Especificaciones Para Realizar Aprovechamiento, Transporte y Almacenamiento de Cogollos. Diario Oficial de la Federacion. 2003. Available online: http://www.profepa.gob.mx/innovaportal/file/3310/1/nom-008-semarnat-1996.pdf (accessed on 22 February 2021).
- Pando-Moreno, M.; Eufracio, O.; Jurado, E.; Estrada, E. Post-harvest growth of Lechuguilla (Agave lecheguilla Torr., Agavaceae) in Northeastern Mexico. Econ. Bot. 2004, 58, 78–82. [Google Scholar] [CrossRef]
- Taylor, N.T.; Davis, K.M.; Abad, H.; McClung, M.R.; Moran, M.D. Ecosystem services of the big bend region of the chihuahuan desert. Ecosyst. Serv. 2017, 27, 48–57. [Google Scholar] [CrossRef]
- Juárez, D.S.; Flores, L.M.L.; Sánchez-Robles, J.H.; De la Cerda, S.P.C.; Rodríguez, C.A.V.; Jiménez, D.L. Variability of saponins concentration in guishe collected in different geographical areas and weather conditions. J. Chem. 2014, 2, 105–115. [Google Scholar] [CrossRef][Green Version]
- Ortiz-Méndez, O.; Morales-Martínez, T.; Rios-González, L.; Rodríguez-de la Garza, J.; Quintero, J.; Aroca, G. Bioethanol production from Agave lechuguilla biomass pretreated by autohydrolysis. Rev. Mex. Ing. Quim. 2017, 16, 467–476. [Google Scholar]
- Díaz-Blanco, D.I.; de La Cruz, J.R.; López-Linares, J.C.; Morales-Martínez, T.K.; Ruiz, E.; Ríos-González, L.J.; Romero, I.; Castro, E. Optimization of dilute acid pretreatment of Agave lechuguilla and ethanol production by co-fermentation with Escherichia coli MM160. Ind. Crops. Prod. 2018, 114, 154–163. [Google Scholar] [CrossRef]
- Oliva-Rodríguez, A.G.; Quintero, J.; Medina-Morales, M.A.; Morales-Martínez, T.K.; Rodríguez-De la Garza, J.A.; Moreno-Dávila, M.; Aroca, G.; Ríos González, L.J. Clostridium strain selection for co-culture with Bacillus subtilis for butanol production from agave hydrolysates. Bioresour. Technol. 2019, 275, 410–415. [Google Scholar] [CrossRef]
- Ríos-González, L.J.; Medina-Morales, M.A.; Rodríguez-De la Garza, J.A.; Romero-Galarza, A.; Medina, D.D.; Morales-Martínez, T.K. Comparison of dilute acid pretreatment of agave assisted by microwave versus ultrasound to enhance enzymatic hydrolysis. Bioresour. Technol. 2021, 319, 124099. [Google Scholar] [CrossRef] [PubMed]
- Juárez, C.; Durán, A.; Valdez, P.; Fajardo, G. Performance of “Agave lecheguilla” natural fiber in portland cement composites exposed to severe environment conditions. Build. Environ. 2007, 42, 1151–1157. [Google Scholar] [CrossRef]
- Dávila-Pompermayer, R.; López-Yepez, L.G.; Valdez-Tamez, P.; Juárez, C.A.; Durán-Herrera, A. Lechuguilla natural fiber as internal curing agent in self compacting concrete (SCC): Mechanical properties, shrinkage and durability. Cem. Concr. Compos. 2020, 112, 103686. [Google Scholar] [CrossRef]
- Romero-González, J.; Peralta-Videa, J.R.; Rodríguez, E.; Delgado, M.; Gardea-Torresdey, J.L. Potential of Agave lechuguilla biomass for Cr(III) removal from aqueous solutions: Thermodynamic studies. Bioresour. Technol. 2006, 97, 178–182. [Google Scholar] [CrossRef] [PubMed]
- Alcázar-Medina, F.; Núñez-Núñez, C.; Rodríguez-Rosales, M.; Valle-Cervantes, S.; Alarcón-Herrera, M.; Proal-Nájera, J. Lead removal from aqueous solution by spherical agglomeration using an extract of Agave lechuguilla Torr. as biosurfactant. Rev. Mex. Ing. Quim. 2019, 19, 71–84. [Google Scholar] [CrossRef]
- Velásquez-Martínez, A.; Díaz-Díaz, A.; Hernández-Escobar, C.A.; Zaragoza-Contreras, E.A. Agave lechuguilla Torrey fiber as reinforcement of polyester resin. Polym. Compos. 2011, 32, 1601–1606. [Google Scholar] [CrossRef]
- Castillo, F.; Hernández, D.; Gallegos, G.; Méndez, M.; Rodríguez, R.; Reyes, A.; Aguilar, C.N. In vitro antifungal activity of plant extracts obtained with alternative organic solvents against Rhizoctonia Solani Kühn. Ind. Crops. Prod. 2010, 32, 324–328. [Google Scholar] [CrossRef]
- Jasso de Rodríguez, D.; Rodríguez García, R.; Hernández Castillo, F.; Aguilar González, C.N.; Sáenz Galindo, A.; Villarreal Quintanilla, J.A.; Moreno Zuccolotto, L.E. In vitro antifungal activity of extracts of Mexican Chihuahuan Desert plants against postharvest fruit fungi. Ind. Crops. Prod. 2011, 34, 960–966. [Google Scholar] [CrossRef]
- Castillo-Reyes, F.; Hernández-Castillo, F.; Clemente-Constantino, J.; Gallegos-Morales, G.; Rodríguez-Herrera, R.; Aguilar, C. In vitro antifungal activity of polyphenols-rich plant extracts against Phytophthora cinnamomi Rands. Afr. J. Agric. Res. 2015, 10, 4552–4560. [Google Scholar] [CrossRef]
- Ontiveros, G.J.G.; Cerna, C.E.; María, Y.; Fuentes, Y.M.O.; Landeros, F.J.; Flores, J.L.; Aguirre, U.L.A.; Hernández, J.A.H. Insecticidal activity of plant extracts against whitefly nymphs Bemisia tabaci (Hemiptera: Aleyrodidae) in laboratory. J. Entomol. Zool. 2020, 8, 595–599. [Google Scholar]
- Peña-Rodríguez, A.; Pelletier-Morreeuw, Z.; García-Luján, J.; Rodríguez-Jaramillo, M.D.C.; Guzmán-Villanueva, L.; Escobedo-Fregoso, C.; Tovar-Ramírez, D.; Reyes, A.G. Evaluation of Agave lechuguilla by-product crude extract as a feed additive for juvenile shrimp Litopenaeus vannamei. Aquac. Res. 2020, 51, 1336–1345. [Google Scholar] [CrossRef]
- Méndez, M.; Rodríguez, R.; Ruiz, J.; Morales-Adame, D.; Castillo, F.; Hernández-Castillo, F.D.; Aguilar, C.N. Antibacterial activity of plant extracts obtained with alternative organics solvents against food-borne pathogen bacteria. Ind. Crops. Prod. 2012, 37, 445–450. [Google Scholar] [CrossRef]
- López-Romero, J.C.; Ayala-Zavala, J.F.; González-Aguilar, G.A.; Peña-Ramos, E.A.; González-Ríos, H. Biological activities of Agave by-products and their possible applications in food and pharmaceuticals. J. Sci. Food Agric. 2017, 98, 2461–2474. [Google Scholar] [CrossRef] [PubMed]
- Padmavathi, M. Chronic disease managment with nutraceuticals. Int. J. Pharm. Sci. Invent. 2013, 2, 1–11. [Google Scholar]
- Casillas, F.R.; Cardenas, A.O.; Morales, C.; Verde Star, M.J.; Cruz-Vega, D.E. Cytotoxic activity of Agave lechuguilla Torr. Afr. J. Biotechnol. 2012, 11, 12229–12231. [Google Scholar] [CrossRef]
- Santos-Zea, L.; Leal-Diaz, A.; Cortes-Ceballos, E.; Gutierrez-Uribes, J. Agave (Agave spp.) and its traditional products as a source of bioactive compounds. Curr. Bioact. Compd. 2012, 8, 1–14. [Google Scholar] [CrossRef]
- Alonso-Castro, A.J.; Domínguez, F.; Maldonado-Miranda, J.J.; Castillo-Pérez, L.J.; Carranza-Álvarez, C.; Solano, E.; Isordia-Espinoza, M.A.; Juárez-Álvarez, M.d.C.; Zapata-Morales, J.R.; Argueta-Fuertes, M.A.; et al. Use of medicinal plants by health professionals in Mexico. J. Ethnopharmacol. 2017, 198, 81–86. [Google Scholar] [CrossRef]
- Morales-Luckie, R.A.; Lopezfuentes-Ruiz, A.A.; Olea-Mejía, O.F.; Argurta-Figueroa, L.; Sanchez-Mendieta, V.; Brostow, W.; Hinestroza, J.P. Synthesis of silver nanoparticles using aqueous extracts of Heterotheca inuloides as reducing agent and natural fibers as templates: Agave lechuguilla and silk. Mater. Sci. Eng. C 2016, 69, 429–436. [Google Scholar] [CrossRef]
- Anguiano-Sevilla, L.A.; Lugo-Cervantes, E.; Ordaz-Pichardo, C.; Rosas-Trigueros, J.L.; Jaramillo-Flores, M.E. Apoptosis induction of Agave lechuguilla Torrey extract on human lung adenocarcinoma cells (SK-LU-1). Int. J. Mol. Sci. 2018, 19, 3795. [Google Scholar] [CrossRef]
- Almaraz-Abarca, N.; González-Elizondo, M.D.S.; Campos, M.D.G.; Ávila-Sevilla, Z.E.; Delgado-Alvarado, E.A.; Ávila-Reyes, J.A. Variability of the foliar phenol profiles of the Agave victoriae-reginae complex (Agavaceae). Bot. Sci. 2013, 91, 265–306. [Google Scholar] [CrossRef]
- Hammuel, C.; Yebpella, G.G.; Shallangwa, G.A.; Magomya, A.M.; Agbaji, A.S. Phytochemical and antimicrobial screening of methanol and aqueous extract of Agave sisalana. Acta Pol. Pharm. 2011, 68, 535–539. [Google Scholar] [PubMed]
- Escamilla-Treviño, L.L. Potential of Plants from the Genus Agave as Bioenergy Crops. Bioenerg. Res. 2011, 5, 1–9. [Google Scholar] [CrossRef]
- Morreeuw, Z.P.; Escobedo-Fregoso, C.; Ríos-González, L.J.; Castillo-Quiroz, D.; Reyes, A.G. Transcriptome-based metabolic profiling of flavonoids in Agave lechuguilla waste biomass. Plant Sci. 2021, 305, 110748. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.M.; Karimi, M.; Venditti, A. Plants adapted to arid areas: Specialized metabolites. Nat. Prod. Res. 2019, 1–18. [Google Scholar] [CrossRef]
- Szabó, S.; Gácsi, Z.; Balázs, B. Specific features of NDVI, NDWI and MNDWI as reflected in land cover categories. Landsc. Environ. 2016, 10, 194–202. [Google Scholar] [CrossRef]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package Version 3.3-13. Available online: https://CRAN.R-project.org/package=raster (accessed on 22 February 2021).
- Singleton, V.; Rossi, J. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Lauranson-Broyer, J.; Lebreton, P. Flavonoids and morphological traits of needles, as markers of natural hybridization between Pinus uncinata Ram and Pinus sylvestris L. Biochem. Syst. Ecol. 1993, 21, 241–247. [Google Scholar] [CrossRef]
- Méndez-Flores, A.; Hernández-Almanza, A.; Saenz-Galindo, A.; Morlett-Chavez, J.; Aguilar, C.N.; Ascacio-Valdés, J. Ultrasound-assisted extraction of antioxidant polyphenolic compounds from Nephelium lappaceum L. (Mexican variety) husk. Asian Pac. J. Trop. Med. 2018, 11, 676–681. [Google Scholar] [CrossRef]
- Rasouli, H.; Farzaei, M.H.; Mansouri, K.; Mohammadzadeh, S.; Khodarahmi, R. Plant cell cancer: May natural phenolic compounds prevent onset and development of plant cell malignancy? A literature review. Molecules 2016, 21, 1104. [Google Scholar] [CrossRef]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Pollet, B.; Legrand, M. Flavonoid accumulation in Arabidopsis repressed in lignin synthesisafects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef]
- Mithöfer, A.; Maffei, M.E. General mechanisms of plant defense and plant toxins. In Plant Toxins; Gopalakrishnakone, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–22. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The origin and evolution of plant flavonoid metabolism. Front. Plant Sci. 2019, 10, 943. [Google Scholar] [CrossRef] [PubMed]
- Mandujano, M.C.; Pisanty, I.; Eguiarte, L.E. (Eds.) Plant Diversity and Ecology in the Chihuahuan Desert; Cuatro Ciénegas Basin: An Endangered Hyperdiverse Oasis; Springer International Publishing: Berlin/Heidelberg, Germany, 2020. [Google Scholar] [CrossRef]
- Nobel, P.S. Achievable productivities of certain CAM plants: Basis for high values compared with C3 and C4 plants. New Phytol. 1991, 119, 183–205. [Google Scholar] [CrossRef]
- Houri, A.; Machaka-Houri, N. Agave lechuguilla as a potential biomass source in arid areas. J. Sustain. Dev. Energy Water Environ. 2016, 4, 89–93. [Google Scholar] [CrossRef]
- Rahmani, H.; Benali, F.T.; Koudach, F.; Dif, M.; Bouazza, S. Phenolic quantification and Agave americana leaves de geoclimatic area. Adv. Environ. Biol. 2016, 10, 194–200. [Google Scholar]
- Nobel, P.S. Environmental Biology of Agaves and Cacti; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]
- Puente-Garza, C.A.; Meza-Miranda, C.; Ochoa-Martínez, D.; García-Lara, S. Effect of in vitro drought stress on phenolic acids, flavonols, saponins, and antioxidant activity in Agave salmiana. Plant Physiol. Biochem. 2017, 115, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef]
- Silva-Montellano, A.; Eguiarte, L.E. Geographic patterns in the reproductive ecology of Agave lechuguilla (Agavaceae) in the Chihuahuan desert. I. Floral characteristics, visitors, and fecundity. Am. J. Bot. 2003, 90, 377–387. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [PubMed]
- López-Lozano, N.E.; Echeverría Molinar, A.; Ortiz Durán, E.A.; Hernández Rosales, M.; Souza, V. Bacterial diversity and interaction networks of Agave lechuguilla rhizosphere differ significantly from bulk soil in the oligotrophic basin of Cuatro Cienegas. Front. Plant Sci. 2020, 11, 1028. [Google Scholar] [CrossRef]
- Sosa, V.; Vásquez-Cruz, M.; Villarreal-Quintanilla, J.A. Influence of climate stability on endemism of the vascular plants of the Chihuahuan Desert. J. Arid. Environ. 2020, 177, 104139. [Google Scholar] [CrossRef]
- Morales-Martínez, T.K.; Medina-Morales, M.A.; Ortiz-Cruz, A.L.; Rodríguez-De la Garza, J.A.; Moreno-Dávila, M.; López-Badillo, C.M.; Ríos-González, L. Consolidated bioprocessing of hydrogen production from agave biomass by Clostridium acetobutylicum and bovine ruminal fluid. Int. J. Hydrogen Energy 2020, 45, 13707–13716. [Google Scholar] [CrossRef]
- Kong, C.H.; Zhao, H.; Xu, X.H.; Wang, P.; Gu, Y. Activity and allelopathy of soil of flavone O-glycosides from rice. J. Agric. Food Chem. 2007, 55, 6007–6012. [Google Scholar] [CrossRef] [PubMed]
- Iser, M.; Martinez, Y.; Ni, H.; Jiang, H.; Valdivié Navarro, M.; Wu, X.; Al-Dhabi, N.A.; Rosales, M.; Duraipandiyan, V.; Fang, J. The Effects of Agave fourcroydes powder as a dietary supplement on growth performance, gut morphology, concentration of IgG and hematology parameters in Broiler Rabbits. BioMed Res. Int. 2016, 2016–3414319. [Google Scholar] [CrossRef]
- Rizwan, K.; Zubair, M.; Rasool, N.; Riaz, M. Phytochemical and Biological Studies of Agave attenuata. Int. J. Mol. Sci. 2012, 13, 6440–6451. [Google Scholar] [CrossRef]
- Ahumada-Santos, Y.; Montes-Avila, J.; Uribe-Beltrán, M.J.; Diaz-Camacho, S.; López-Angulo, G.; Vega-Aviña, R.; López-Valenzuela, J.A.; Heredia, J.B.; Delgado-Vargas, F. Chemical characterization, antioxidant and antibacterial activities of six Agave species from Sinaloa, Mexico. Ind. Crops. Prod. 2013, 49, 143–149. [Google Scholar] [CrossRef]
- Carmona, J.E.; Morales-Martínez, T.K.; Mussatto, S.I.; Castillo-Quiroz, D.; Ríos-González, L.J. Propriedades quimicas, estructurales y funcionales de la lechuguilla (Agave lechuguilla Torr.). Rev. Mex. Cienc. For. 2017, 8, 100–122. [Google Scholar]
- Torres-García, I.; Rendón-Sandoval, F.J.; Blancas, J.; Casas, A.; Moreno-Calles, A.I. The genus Agave in agroforestry systems of Mexico. Bot. Sci. 2019, 97, 263–290. [Google Scholar] [CrossRef]
- Campos, H.; Trejo, C.; Peña-Valdivia, C.B.; García-Nava, R.; Conde-Martínez, F.V.; Cruz-Ortega, R. Water availability effects on germination, membrane stability and initial root growth of Agave lechuguilla and A. salmiana. Flora 2020, 268, 151606. [Google Scholar] [CrossRef]
- Food and Drug Administration (FDA). 2020. Available online: https://www.fda.gov/ (accessed on 22 February 2021).
- Mammen, D.; Daniel, M. A critical evaluation on the reliability of two aluminum chloride chelation methods for quantification of flavonoids. Food Chem. 2012, 135, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Kejík, Z.; Kaplánek, R.; Masařík, M.; Babula, P.; Matkowski, A.; Filipenský, P.; Veselá, K.; Gburek, J.; Sýkora, D.; Martásek, P.; et al. Iron complexes of flavonoids-antioxidant capacity and beyond. Int. J. Mol. Sci. 2021, 22, 646. [Google Scholar] [CrossRef]
- Iser, M.; Valdivié, M.; Figueredo, L.; Núñez, E.; Mas, D.; Martínez, Y. Secondary metabolites, quality indicators and organoleptic characteristics of stems meal from Agave fourcroydes (Henequen). Cuba. J. Agric. Sci. 2020, 54. Available online: http://www.cjascience.com/index.php/CJAS/article/view/937 (accessed on 22 February 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak N° | Compounds | Concentration (µg/g DW) | |||||
|---|---|---|---|---|---|---|---|
| Site | |||||||
| Matehuala | Cosme | Tuxtepec | |||||
| Extractive Solvent | |||||||
| MetOH | EtOH | MetOH | EtOH | MetOH | EtOH | ||
| 1 | Flavanone ** | 520.13 ± 39.98 c | 276.19 ± 16.35 ab | 359.86 ± 55.98 b | 224.78 ± 19.59 a | 214.10 ± 24.10 a | 284.01 ± 10.61 ab |
| 2 | Isorhamnetin ** | 1416.70 ± 150.32 c | 753.97 ± 48.25 a | 981.55 ± 89.85 b | 614.32 ± 24.94 a | 1251.96 ± 58.24 c | 752.80 ± 2.91 a |
| 4 | Catechin * | 2.79 ± 0.26 a | 1.80 ± 0.07 a | 3.98 ± 1.30 ab | 4.97 ± 2.26 ab | 7.91 ± 3.62 b | 4.38 ± 0.50 ab |
| 5 | Cyanidin ** | 0.00 a | 0.00 a | 0.00 a | 3.53 ± 0.22 b | 0.00 a | 10.48 ± 2.61 c |
| 6 | Delphinidin ** | 0.00 a | 0.00 a | 0.00 a | 11.42 ± 0.32 b | 0.00 a | 21.55 ± 3.79 c |
| 7 | Hesperidin ** | 32.90 ± 4.54 b | 4.00 ± 0.13 a | 32.96 ± 3.73 b | 4.05 ± 0.29 a | 34.23 ± 0.95 b | 4.69 ± 0.19 a |
| 8 | Quercetin * | 12.08 ± 0.69 a | 11.65 ± 0.66 a | 13.24 ± 0.80 ab | 13.73 ± 0.25 ab | 15.57 ± 2.47 b | 13.48 ± 0.32 ab |
| 9 | Apigenin * | 6.93 ± 0.29 a | 7.88 ± 0.33 ab | 7.95 ± 0.54 ab | 7.85 ± 0.84 ab | 9.70 ± 1.96 b | 9.25 ± 0.61 ab |
| 10 | Kaempferol | 12.29 ± 1.37 a | 12.78 ± 0.75 a | 12.82 ± 0.27 a | 12.77 ± 0.43 a | 13.71 ± 1.01 a | 13.48 ± 0.31 a |
| 11 | Naringenin * | 1.26 ± 0.08 a | 1.43 ± 0.07 ab | 1.45 ± 0.10 ab | 1.43 ± 0.15 ab | 1.76 ± 0.36 b | 1.68 ± 0.11 ab |
| Total | 2005.08 | 1069.70 | 1413.56 | 898.85 | 1548.94 | 1115.80 | |
| Compounds 1 | m/z ([M − H]− Fragments) | Matehuala | Cosme | Tuxtepec |
|---|---|---|---|---|
| Apigenin | 268.9 (203.9) | + | + | + |
| Apigenin 7-O-glycoside | 431.1 (268.9; 203.9) | + | + | + |
| Apigenin 7-O-rutinoside | 578.5 (268.9) | + | + | + |
| Catechin | 288.9 | + | + | + |
| Cyanidin | 287.1 | − | + | + |
| Cyanidin 3-O-glycoside | 478.4 (287.1) | − | + | + |
| Cyanidin O-diglycoside | 609.2 (449.3; 287.1) | + | + | + |
| Delphinidin | 319.1 | − | + | + |
| Delphinidin 3-O-glycoside | 465.4 (319.0) | − | + | + |
| Flavanone | 223.1 | + | + | + |
| Hesperidin | 610.2 | + | + | + |
| Isorhamnetin | 316.1 | + | + | + |
| Isorhamnetin-glycoside | 478.4 (272.1) | + | + | + |
| Isorhamnetin 3-O-rutinoside | 622.3 (146.1) | + | + | + |
| Isorhamnetin diglycoside 1 | 579.4 (316.1; 272.1) | − | + | + |
| Isorhamnetin diglycoside 2 | 609.2 (449.1; 272.1; 146.1) | + | + | + |
| Isorhamnetin triglycoside 1 | 756.5 (162.8; 146.1) | − | + | + |
| Isorhamnetin triglycoside 2 | 801.6 (316.1) | + | − | + |
| Kaempferol | 285.1 (153.0; 107.0) | + | + | + |
| Kaempoferol 3-O-glycoside | 431.1 | + | + | + |
| Kaempferol 3-O-rutinoside | 593.4 (285.1; 431.1) | + | + | + |
| Kaempoferol diglycoside | 595.1 (285.1) | + | + | + |
| Kaempoferol triglycoside | 772.5 (285.0) | + | + | + |
| Myricetin | 317.0 | + | + | + |
| Myricetin 3-O-glycoside | 463.1 (317.0; 179.1) | + | - | + |
| Myricetin diglycoside | 625.2 (317.0) | + | + | + |
| Naringenin | 270.9 | + | + | + |
| Naringenin O-rutinoside | 579.5 (270.9) | + | + | + |
| Quercetin | + | + | + | |
| Quercetin-3-O-xyloside | 433.2 (300.9; 179.1) | + | + | + |
| Quercetin 3-O-glycoside 1 | 447.1 (301.0; 146.1) | + | + | + |
| Quercetin 3-O-glycoside 2 | 463.1 (301.0; 177.0) | − | + | + |
| Quercetin 3-O-rutinoside | 609.2 (300.9; 463.1; 146.1) | + | + | + |
| Quercetin 3-O-diglycoside 1 | 595.1 (300.9; 447.1) | + | + | + |
| Quercetin 3-O-diglycoside 2 | 595.1 (300.9; 463.1) | − | + | + |
| Matehuala (San Luis Potosi) | Cosme-Tuxtepec (Coahuila) | ||
|---|---|---|---|
| Scientific Name | Family | Scientific Name | Family |
| Hechtia texensis S. Watson | Bromeliaceae | Larrea tridentata (DC.) Colville | Zygophyllaceae |
| Karwinskia humboldtiana (Schult.) Zucc. | Rhamnaceae | Fouquieria splendens Engelm. | Fouquieriaceae |
| Yucca carnerosana (Trel.) McKelvey | Asparagaceae | Euphorbia antisyphilitica Zucc. | Euphorbiaceae |
| Echinocactus platyacantus Link and Otto | Cactaceae | Hechtia texensis S. Watson | Bromeliaceae |
| Jatropha dioica Sessé ex Cerv. | Euphorbiaceae | Opuntia microdasys (Lehm.) Pfeiff. | Cactaceae |
| Leucophyllum laevigatum Standl. | Scrophulariaceae | Flourensia cernua DC. | Asteraceae |
| Viguieria stenoloba S.F. Blake | Asteraceae | Echinocereus stramineus (Engelm.) Rümpler | Cactaceae |
| Prosopis glandulosa (DC.) F.M. Knuth | Fabaceae | Cylindropuntia leptocaulis (DC.) F.M. Knuth | Cactaceae |
| Notholaena sinuata (Lag. ex Sw.) Kaulf. | Pteridaceae | Cylindropuntia kleiniae (DC.) F.M. Knuth | Cactaceae |
| Cylindropuntia kleiniae (DC.) F.M. Knuth | Cactaceae | Acacia berlandieri Benth. | Fabaceae |
| Larrea tridentata (DC.) Coville | Zygophyllaceae | Vachellia constricta (Benth.) Seigler and Ebinger | Fabaceae |
| Cylindropuntia imbricata (Haw.) F.M. Knuth | Cactaceae | Epithelantha micromeris (Engelm.) F.A.C. Weber ex Britton & Rose | Cactaceae |
| Agave striata subsp. falcata (Engelm.) Gentry | Asparagaceae | ||
| Echinocereus conglomeratus C.F. Först. | Cactaceae | ||
| Jatropha dioica Sessé ex Cerv | Euphorbiaceae | ||
| Environmental Parameter | Matehuala | Cosme | Tuxtepec |
|---|---|---|---|
| Location | 23°28′54.3″ N; 100°37′22.1″ W | 25°52′03.6″ N; 101°19′51.1″ W | 26°11′29.8″ N; 101°11′0.96″ W |
| Temperature | 16.0–29.0 °C; Max 35 °C | 18.8–30.6 °C; Max 40 °C | 19.7–32.9 °C; Max, 35.9 °C |
| Photoperiod | 13 h 12 min–12 h 36 min | 13 h 18 min–12 h 39 min | 13 h 18 min–12 h 39 min |
| Cloudy | 36–42% | 47–49% cloudy | 47–49% |
| Precipitation | 55.6 mm | 44.8 mm | 29.0 mm |
| Rainy days | 4.3 days | 2.8 days | 3.4 days |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morreeuw, Z.P.; Castillo-Quiroz, D.; Ríos-González, L.J.; Martínez-Rincón, R.; Estrada, N.; Melchor-Martínez, E.M.; Iqbal, H.M.N.; Parra-Saldívar, R.; Reyes, A.G. High Throughput Profiling of Flavonoid Abundance in Agave lechuguilla Residue-Valorizing under Explored Mexican Plant. Plants 2021, 10, 695. https://doi.org/10.3390/plants10040695
Morreeuw ZP, Castillo-Quiroz D, Ríos-González LJ, Martínez-Rincón R, Estrada N, Melchor-Martínez EM, Iqbal HMN, Parra-Saldívar R, Reyes AG. High Throughput Profiling of Flavonoid Abundance in Agave lechuguilla Residue-Valorizing under Explored Mexican Plant. Plants. 2021; 10(4):695. https://doi.org/10.3390/plants10040695
Chicago/Turabian StyleMorreeuw, Zoé P., David Castillo-Quiroz, Leopoldo J. Ríos-González, Raúl Martínez-Rincón, Norma Estrada, Elda M. Melchor-Martínez, Hafiz M. N. Iqbal, Roberto Parra-Saldívar, and Ana G. Reyes. 2021. "High Throughput Profiling of Flavonoid Abundance in Agave lechuguilla Residue-Valorizing under Explored Mexican Plant" Plants 10, no. 4: 695. https://doi.org/10.3390/plants10040695
APA StyleMorreeuw, Z. P., Castillo-Quiroz, D., Ríos-González, L. J., Martínez-Rincón, R., Estrada, N., Melchor-Martínez, E. M., Iqbal, H. M. N., Parra-Saldívar, R., & Reyes, A. G. (2021). High Throughput Profiling of Flavonoid Abundance in Agave lechuguilla Residue-Valorizing under Explored Mexican Plant. Plants, 10(4), 695. https://doi.org/10.3390/plants10040695