
Anti-Inflammatory Effects of Berchemia floribunda in LPS-Stimulated RAW264.7 Cells through Regulation of NF-κB and MAPKs Signaling Pathway


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Preparation of the Extracts of Leaves, Branches and Fruit from BF
2.3. Cell Culture and Treatment
2.4. Measurement of Cell Viability
2.5. NO Inhibition Activity
2.6. Isolation of Nucleus Fraction
2.7. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
2.8. Sodium Dodecyl Sulphate–Polyacrylamide Gel Electrophoresis (SDS-PAGE) and Western Blot
2.9. Statistical Analysis
3. Results
3.1. Effect of BF-L on NO Production and iNOS, COX-2, IL-β, IL-6 and TNF-α Expression in LPS-Stimulated RAW264.7 Cells
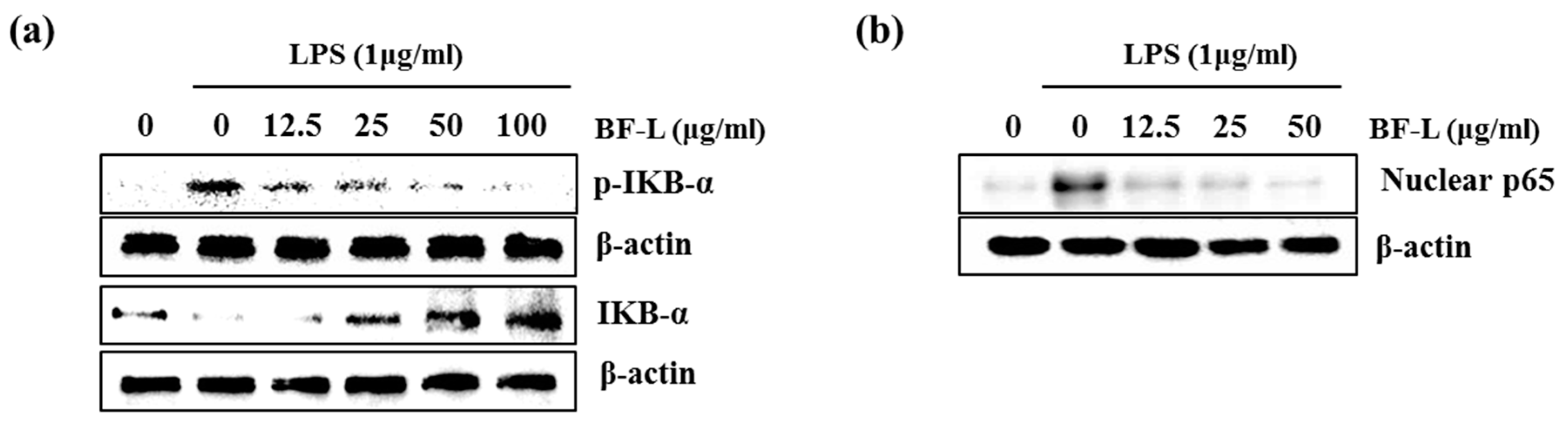
3.2. Effect of BF-L on NF-κB Signaling Activation in LPS-Stimulated RAW264.7 Cells
3.3. Effect of BF-L on MAPKs Signaling Activation in LPS-Stimulated RAW264.7 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, M.J.; Kim, K.B.W.R.; Jeong, D.H.; Ahn, D.H. Inhibitory effect of Sargassum fulvellum ethanolic extract on LPS induced inflammatory reaction in RAW 264.7 mouse macrophages. J. Appl. Biol. Chem. 2013, 56, 249–255. [Google Scholar] [CrossRef][Green Version]
- Lee, H.S.; Chon, S.H.; Kim, M.A.; Park, J.E.; Lim, Y.M.; Kim, E.J.; Son, E.K.; Kim, S.J.; So, J.H. Fermentation enhances the antioxidant and anti-inflammatory effects of Bat Faeces (Ye Ming Sha) via the ERK, p38 MAPK and NF-κB signaling pathways in RAW 264.7 cells. J. Appl. Biol. Chem. 2019, 62, 57–66. [Google Scholar] [CrossRef]
- Lim, Y.Y.; Kim, H.M.; Park, W.S.; Kim, J.H.; Shin, J.H.; Kim, M.N.; Kim, B.J. Anti-inflammatory and anti-pruritic effects of Portulaca oleracea L. extract using in vitro and in vivo inflammation model: LPS- treated RAW 264.7 cells, keratinocytes, NC/NGAmice and hairless SKH-1 mice. Korean J. Asthma Allergy Clin. Immunol. 2011, 31, 199–206. [Google Scholar]
- Lee, H.N.; Lim, D.Y.; Lim, S.Y.; Kim, J.D.; Yoon, J.H. Anti-inflammatory effect of ethanol extract from Eupatorium japonicum. Korean J. Food Sci. Technol. 2001, 43, 65–71. [Google Scholar] [CrossRef]
- Won, S.J.; Park, H.J.; Lee, K.T. Inhibition of LPS induced iNOS, COX-2 and cytokines expression by salidroside through the NF-κB inactivation in RAW 264.7 cells. Korean J. Pharm. 2008, 39, 110–117. [Google Scholar]
- Lee, S.H.; Jin, K.S.; Son, Y.R.; Kwon, H.J.; Kim, B.W. Anti-oxidative and anti-inflammatory activities of Desmodium heterocarpon extract in RAW 264.7 cells. J. Life Sci. 2018, 28, 216–222. [Google Scholar]
- Ahmed, H.; Khan, M.Z.I.; Waseem, D.; Nazli, A.; Baig, M.W. Phytochemical analysis and antioxidant potential of Ficus Benghalensis L. J. Bioresour. Manag. 2017, 4, 17–26. [Google Scholar]
- Mutuma, G.G.; Joseph, N.; King’ori, M.A.; Silas, K. Phytochemical and anti-inflammatory analysis of Prunus africana bark extract. Res. J. Pharm. 2020, 7, 31–38. [Google Scholar]
- Eo, H.J.; Kim, D.S.; Kang, Y.; Kim, K.; Park, Y.; Park, G.H. Antioxidant and immunoregulatory effects of Korean Rhamnaceae. J. Plant Biotechnol. 2020, 47, 254–259. [Google Scholar] [CrossRef]
- Song, J.H.; Lim, H.I.; Jang, K.H.; Han, J. Characteristics of leaf morphology and genetic variation of the rare woody plant, Berchemia racemosa var. magna. Korean J. Plant Res. 2013, 26, 613–618. [Google Scholar] [CrossRef]
- Song, J.H.; Lim, H.I.; Jang, K.H.; Hong, K.N.; Han, J.G. Genetic diversity and special genetic structure of Berchemia racemosa var. magna in Anmyeon island. Korean J. Hort. Sci. Technol. 2014, 32, 84–90. [Google Scholar]
- Kang, Y.; Eo, H.J.; Kim, D.S.; Park, Y.; Song, J.H.; Park, G.H. Berchemia floribunda-mediated proteasomal degradation of Cyclin D1 via GKS3β-dependent threonine-286 phosphorylation in human colorectal cancer cells. Korean J. Plant Res. 2020, 33, 271–278. [Google Scholar]
- Yan, H.J.; Li, Y.H.; Zhang, H.Y.; He, M.L. Pharmacognostical study on Berchemia floribunda. Zhong Yao Cai 2013, 36, 908–910. [Google Scholar] [PubMed]
- Vassallo, A.; Armentano, M.F.; Miglionico, R.; Caddeo, C.; Chirollo, C.; Gualtieri, M.J.; Ostuni, A.; Bisaccia, F.; Faraone, I.; Milella, L. Hura crepitans L. extract: Phytochemical characterization, antioxidant activity, and nanoformulation. Pharmaceutics 2020, 12, 553. [Google Scholar] [CrossRef]
- Sinisgalli, C.; Faraone, I.; Vassallo, A.; Caddeo, C.; Bisaccia, F.; Armentano, M.F.; Milella, L.; Ostuni, A. Phytochemical profile of Capsicum annuum L. cv Senise, incorporation into liposomes, and evaluation of cellular antioxidant activity. Antioxidants 2020, 9, 428. [Google Scholar] [CrossRef]
- Faraone, I.; Rai, D.K.; Chiummiento, L.; Fernandez, E.; Choudhary, A.; Prinzo, F.; Milella, L. Antioxidant activity and phytochemical characterization of Senecio clivicolus Wedd. Molecules 2018, 23, 2497. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, N.; Silva, E.M.E. Introduction to Phytochemicals: Secondary Metabolites from Plants with Active Principles for Pharmacological Importance. Phytochem. Source Antioxid. Role Dis. Prev. 2018, 25. [Google Scholar] [CrossRef]
- Wang, Y.F.; Cao, J.X.; Efferth, T.; Lai, G.F.; Luo, S.D. Cytotoxic and new tetralone derivatives from Berchemia floribunda (Wall.). Brongn. Chem. Biodivers. 2006, 3, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Jiang, J.S.; Feng, Z.M.; Zhang, P.C. Anthraquinone-benzisochromanquinone dimers from the roots of Berchemia floribunda. Chem. Pharm. Bull. 2008, 56, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar]
- Comalada, M.; Camuesco, D.; Sierra, S.; Ballester, I.; Xaus, J.; Gálvez1, J.; Zarzuelo, A. In vivo quercitrin anti-inflammatory effect involves release of quercetin, which inhibits inflammation through down-regulation of the NF-κB pathway. Eur. J. Immunol. 2005, 35, 584–592. [Google Scholar] [CrossRef]
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, antioxidant and anti-inflammatory activities of kaempferol and its corresponding glycosides and the enzymatic preparation of kaempferol. PLoS ONE 2018, 13, e0197563. [Google Scholar] [CrossRef] [PubMed]
- Durmaz, L. Antioxidant, antiepileptic, and anticholinergic properties of 4′,5,7-Trihydroxy-3,6-dimethoxyflavone as natural phenolic compound: A toxicology approach. Toxin Rev. 2019, 1–8. [Google Scholar] [CrossRef]
- Islam, M.N.; Ishita, I.J.; Jin, S.E.; Choi, R.J.; Lee, C.M.; Kim, Y.S.; Jung, H.A.; Choi, J.S. Anti-inflammatory activity of edible brown alga Saccharina japonica and its constituents pheophorbide a and pheophytin a in LPS-stimulated RAW 264.7 macrophage cells. Food Chem. Toxicol. 2013, 55, 541–548. [Google Scholar] [CrossRef]
- Jew, S.S.; Bae, O.N.; Chung, J.H. Anti-inflammatory effects of asiaticoside on inducible nitric oxide synthase and cyclooxygenase-2 in RAW 264.7 cell line. J. Toxicol. Public Health 2003, 19, 33–37. [Google Scholar]
- Lu, J.; Fang, K.; Wan, S.; Xiong, L.; Zhang, C.; Liu, Z.; Guan, X.; Zheng, R.; Wang, G.; Zheng, J.; et al. Anti-inflammatory effect of columbianetin on lipopolysaccharide-stimulated human peripheral blood mononuclear cells. Mediat. Inflamm. 2018, 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.T.; Jeong, Y.R.; Ha, M.H.; Kim, S.H.; Byun, M.W.; Jo, S.K. Induction of nitric oxide and TNF-α by herbal plant extracts in mouse macrophages. J. Korean Soc. Food Sci. Nutr. 2000, 29, 342–348. [Google Scholar]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, C.; Jia, X.; Wang, K.; Tu, Y.; Wang, R.; Liu, K.; Lu, T.; He, C. In vitro and in vivo anti-inflammatory effects of polyphyllin VII through downregulating MAPK and NF-B pathways. Molecules 2019, 24, 875. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal. Transduct. Target. Ther. 2017, 2, e17023. [Google Scholar] [CrossRef]
- Kim, H.N.; Park, G.H.; Park, S.B.; Kim, J.D.; Eo, H.J.; Son, H.J.; Song, J.H.; Jeong, J.B. Sageretia thea inhibits inflammation through suppression of NF-κB and MAPK and activation of Nrf2/HO-1 signaling pathways in RAW264.7 cells. Am. J. Chin. Med. 2019, 47, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Wang, Y.; Dong, L.; Li, M.; Cai, W. Anti-inflammatory effect of resveratrol through the suppression of NF-κB and JAK/STAT signaling pathways. Acta Biochim. Biophys. Sin. 2015, 47, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K. Role of ERK/MAPK signalling pathway in anti-inflammatory effects of Ecklonia cava in activated human mast cell line-1 cells. Asian Pac. J. Trop. Med. 2014, 7, 703–708. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eo, H.J.; Jang, J.H.; Park, G.H. Anti-Inflammatory Effects of Berchemia floribunda in LPS-Stimulated RAW264.7 Cells through Regulation of NF-κB and MAPKs Signaling Pathway. Plants 2021, 10, 586. https://doi.org/10.3390/plants10030586
Eo HJ, Jang JH, Park GH. Anti-Inflammatory Effects of Berchemia floribunda in LPS-Stimulated RAW264.7 Cells through Regulation of NF-κB and MAPKs Signaling Pathway. Plants. 2021; 10(3):586. https://doi.org/10.3390/plants10030586
Chicago/Turabian StyleEo, Hyun Ji, Jun Hyuk Jang, and Gwang Hun Park. 2021. "Anti-Inflammatory Effects of Berchemia floribunda in LPS-Stimulated RAW264.7 Cells through Regulation of NF-κB and MAPKs Signaling Pathway" Plants 10, no. 3: 586. https://doi.org/10.3390/plants10030586
APA StyleEo, H. J., Jang, J. H., & Park, G. H. (2021). Anti-Inflammatory Effects of Berchemia floribunda in LPS-Stimulated RAW264.7 Cells through Regulation of NF-κB and MAPKs Signaling Pathway. Plants, 10(3), 586. https://doi.org/10.3390/plants10030586