
Seed Dormancy and Preharvest Sprouting in Quinoa (Chenopodium quinoa Willd)



Abstract
1. Introduction to Quinoa, Cultivars, Breeding Issues, and Preharvest Sprouting
2. Model Systems: A Theoretical Framework for Quinoa Seed Dormancy, Hormone Signaling, and PHS
3. Orthodox and Unorthodox Seed Types
4. Physical Dormancy in Quinoa
5. Environmental Regulation of Quinoa Seed Dormancy
6. Breeding Strategies to Mitigate PHS in Quinoa
7. Other Tools for Mitigating PHS in Quinoa
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morales, A.; Zurita-Silva, A.; Maldonado, J.; Silva, H. Transcriptional responses of chilean quinoa (Chenopodium quinoa Willd.) under water deficit conditions uncovers ABA-independent expression patterns. Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Albani, D.; Hammond-Kosack, M.C.U.; Smith, C.; Conlan, S.; Colot, V.; Holdsworth, M.; Bevan, M.W. The wheat transcriptional activator SPA: A seed-specific bZIP protein that recognizes the GCN4-like motif in the bifactorial endosperm box of prolamin genes. Plant Cell 1997, 9, 171–184. [Google Scholar]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa abiotic stress responses: A review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [PubMed]
- Tapia, M. The long journey of quinoa: Who wrote its history. In State of the Art Report on Quinoa around the World; Bazile, D., Bertero, H.D., Nieto, C., Eds.; FAO: Santiago, Chile; CIRAD: Montpellier, France, 2013; Volume 1, pp. 1–7. ISBN 978-92-5-108558-5. [Google Scholar]
- Risi, J.; Galwey, N.W. The Chenopodium grains of the Andes: Inca crops for modern agriculture. Adv. Appl. Biol. 1984, 10, 145–216. [Google Scholar]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef]
- Sanchez, H.B.; Lemeur, R.; Damme, P.V.; Jacobsen, S.-E. Ecophysiological analysis of drought and salinity stress of quinoa (Chenopodium Quinoa willd.). Food Rev. Int. 2003, 19, 111–119. [Google Scholar] [CrossRef]
- Jacobsen, S.-E.; Mujica, A.; Jensen, C.R. The resistance of quinoa (Chenopodium quinoa Willd.) to adverse abiotic factors. Food Rev. Int. 2003, 19, 99–109. [Google Scholar] [CrossRef]
- Adolf, V.I.; Jacobsen, S.-E.; Shabala, S. Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Ruiz-Carrasco, K.; Antognoni, F.; Coulibaly, A.K.; Lizardi, S.; Covarrubias, A.; Martínez, E.A.; Zurita-Silva, A. Variation in salinity tolerance of four lowland genotypes of quinoa (Chenopodium quinoa Willd.) as assessed by growth, physiological traits, and sodium transporter gene expression. Plant Physiol. Biochem. 2011, 49, 1333–1341. [Google Scholar] [CrossRef]
- Walck, J.L.; Baskin, J.M.; Baskin, C.C.; Hidayati, S.N. Defining transient and persistent seed banks in species with pronounced seasonal dormancy and germination patterns. Seed Sci. Res. 2005, 15, 189–196. [Google Scholar] [CrossRef]
- Arc, E.; Sechet, J.; Corbineau, F.; Rajjou, L.; Marion-Poll, A. ABA crosstalk with ethylene and nitric oxide in seed dormancy and germination. Front. Plant Sci. 2013, 4, 1–19. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef]
- Willis, C.G.; Baskin, C.C.; Baskin, J.M.; Auld, J.R.; Venable, D.L.; Cavender-Bares, J.; Donohue Kathleen de Casas, R.R.; The NESCent Germination Working Group. The evolution of seed dormancy: Environmental cues, evolutionary hubs, and diversification of the seed plants. New Phytolog. 2014, 203, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Nikolaeva, M.G. Physiology of Deep Dormancy in Seeds; Leningrad, Russia: Izdateljstvo ‘Nauka’ (in Russian). [Translated from Russian by Z. Shapiro (1969). National Science Foundation: Washington, DC, USA, 1967; p. 219. [Google Scholar]
- Nikolaeva, M.G. On criteria to use in studies of seed evolution. Seed Sci. Res. 2004, 14, 315–320. [Google Scholar] [CrossRef]
- Karssen, C.M. Uptake and effect of abscisic acid during induction and progress of radicle growth in seeds of Chenopodium album. Physiolog. Plant. 1976, 36, 259–263. [Google Scholar] [CrossRef]
- Hauvermale, A.L.; Ariizumi, T.; Steber, C.M. Gibberellin signaling: A theme and variations on DELLA repression. Plant Physiolog. 2012, 160, 83–92. [Google Scholar] [CrossRef]
- Tuttle, K.M.; Martinez, S.A.; Schramm, E.C.; Takebayashi, Y.; Seo, M.; Steber, C.M. Grain dormancy loss is associated with changes in ABA and GA sensitivity and hormone accumulation in bread wheat, Triticum aestivum L. Seed Sci. Res. 2015, 25, 179–193. [Google Scholar] [CrossRef]
- Koornneef, M.; Jorna, M.L.; der Brinkhorst-van Swan, D.L.C.; Karssen, C.M. The isolation of abscisic acid (ABA) deficient mutants by selection of induced revertants in non-germinating gibberellin sensitive lines of Arabidopsis thaliana L. heynh. Theor. Appl. Gen. 1982, 61, 385–393. [Google Scholar] [CrossRef]
- Finkelstein, R.R.; Reeves, W.; Ariizumi, T.; Steber, C.M. Molecular aspects of seed dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef]
- Karssen, C.M.; Lacka, E. A revision of the hormone balance theory of seed dormancy: Studies on gibberellin and/or abscisic acid-deficient mutants of Arabidopsis thaliana. Plant Growth Sub. 1985, 315–323. [Google Scholar]
- Field, R.J.; Jackson, D.I. A hormone balance theory of apical dominance. In Bulletin of the Royal Society of New Zealand; Springer: Berlin, Germany, 1974; pp. 655–657. [Google Scholar]
- Luckwill, L. Growth-inhibiting and growth-promoting substances in relation to the dormancy and after-ripening of apple seeds. J. Hort. Sci. 1952, 27, 53–67. [Google Scholar] [CrossRef]
- Walker-Simmons, M. ABA levels and sensitivity in developing wheat embryos of sprouting resistant and susceptible cultivars. Plant Physiol. 1987, 84, 61–66. [Google Scholar] [CrossRef]
- Morris, C.F.; Moffatt, J.M.; Sears, R.G.; Paulsen, G.M. Seed dormancy and responses of caryopses, embryos, and calli to abscisic acid in wheat. Plant Physiol. 1989, 90, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Taylor, I.B.; Burbidge, A.; Thompson, A.J. Control of abscisic acid synthesis. J. Exper. Bot. 2000, 51, 1563–1574. [Google Scholar] [CrossRef]
- Kushiro, T.; Okamoto, M.; Nakabayashi, K.; Yamagishi, K.; Kitamura, S.; Asami, T.; Hirai, N.; Koshiba, T.; Kamiya, Y.; Nambara, E. The Arabidopsis cytochrome P450 CYP707A encodes ABA 8’-hydroylases: Key enzymes in ABA catabolism. Emb. J. 2004, 23, 1647–1656. [Google Scholar] [CrossRef]
- Chono, M.; Honda, I.; Shinoda, S.; Kushiro, T.; Kamiya, Y.; Nambara, E.; Kawakami, N.; Kaneko, S.; Watanabe, Y. Field studies on the regulation of abscisic acid content and germinability during grain development of barley: Molecular and chemical analysis of pre-harvest sprouting. J. Exper. Bot. 2006, 57, 2421–2434. [Google Scholar] [CrossRef] [PubMed]
- Finch-Savage, W.E.; Leubner-Metzger, G. Seed dormancy and the control of germination. New Phytolog. 2006, 171, 501–523. [Google Scholar] [CrossRef]
- Millar, A.A.; Jacobsen, J.V.; Ross, J.J.; Helliwell, C.A.; Poole, A.T.; Scofield, G.; Reid, J.B.; Gubler, F. Seed dormancy and ABA metabolism in arabidopsis and barley: The role of ABA 8’-hydroxylase. Plant J. 2006, 45, 942–954. [Google Scholar] [CrossRef]
- Okamoto, M.; Kuwahara, A.; Seo, M.; Kushiro, T.; Asami, T.; Hirai, N.; Kamiya, Y.; Koshiba, T.; Nambara, E. CYP707A1 and CYP707A2, which encode abscisic acid 8’-hydroxylases, are indispensable for proper control of seed dormancy and germination in Arabidopsis. Plant Physiol. 2006, 141, 97–107. [Google Scholar] [CrossRef]
- Barrero, J.M.; Talbot, M.J.; White, R.G.; Jacobsen, J.V.; Gubler, F. Anatomical and transcriptomic studies of the coleorhiza reveal the importance of this tissue in regulating dormancy in barley. Plant Physiol. 2009, 150, 1006–1021. [Google Scholar] [CrossRef]
- Schramm, E.C.; Abellera, J.C.; Strader, L.C.; Campbell, K.G.; Steber, C.M. Isolation of ABA-responsive mutants in allohexaploid bread wheat (Triticum aestivum L.): Drawing connections to grain dormancy, preharvest sprouting, and drought tolerance. Plant Sci. 2010, 179, 620–629. [Google Scholar] [CrossRef]
- Barrero, J.M.; Jacobsen, J.V.; Talbot, M.J.; White, R.G.; Swain, S.M.; Garvin, D.F.; Gubler, F. Grain dormancy and light quality effects on germination in the model grass Brachypodium distachyon. New Phytol. 2012, 193, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Ariizumi, T.; Hauvermale, A.L.; Nelson, S.K.; Hanada, A.; Yamaguchi, S.; Steber, C.M. Lifting DELLA repression of Arabidopsis seed germination by nonproteolytic gibberellin signaling. Plant Physiol. 2013, 162, 2125–2139. [Google Scholar] [CrossRef] [PubMed]
- Bewley, J.D.; Bradford, K.J.; Hilhorst, H.W.M.; Nonogaki, H. SEEDS: Physiology of Development, Germination and Dormancy; Springer: New York, NY, USA, 2013. [Google Scholar]
- Chono, M.; Matsunaka, H.; Seki, M.; Fujita, M.; Kiribuchi-Otobe, C.; Oda, S.; Kojima, H.; Kobayashi, D.; Kawakami, N. Isolation of a wheat (Triticum aestivum L.) mutant in ABA 8’-hydroxylase gene: Effect of reduced ABA catabolism on germination inhibition under field condition. Breed. Sci. 2013, 63, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Schramm, E.C.; Nelson, S.K.; Kidwell, K.K.; Steber, C.M. Increased ABA sensitivity results in higher seed dormancy in soft white spring wheat cultivar ‘Zak’. Theor. Appl. Gen. 2013, 126, 791–803. [Google Scholar] [CrossRef]
- Barrero, J.M.; Downie, A.B.; Xu, Q.; Gubler, F. A role for barley CRYPTOCHROME1 in light regulation of grain dormancy and germination. Plant Cell 2014, 26, 1094–1104. [Google Scholar] [CrossRef]
- Hauvermale, A.L.; Tuttle, K.M.; Takebayashi, Y.; Seo, M.; Steber, C.M. Loss of Arabidopsis thaliana seed dormancy is associated with increased accumulation of the GID1 GA hormone receptors. Plant Cell Physiol. 2015, 56, 1773–1785. [Google Scholar] [CrossRef]
- Koornneef, M.; van der Veen, J.H. Induction and analysis of gibberellin sensitive mutants in Arabidopsis thaliana L. heynh. Theor. Appl. Gen. 1980, 58, 257–263. [Google Scholar] [CrossRef]
- Jacobsen, J.V.; Pearce, D.W.; Poole, A.T.; Pharis, R.P.; Mander, L.N. Abscisic acid, phaseic acid and gibberellin contents associated with dormancy and germination in barley. Physiol. Plant. 2002, 115, 429–441. [Google Scholar] [CrossRef]
- Gualano, N.; Carrari, F.; Rodriguez, M.V.; Perez-Flores, L.; Sanchez, R.; Iusem NBenech-Arnold, R.L. Reduced embryo sensitivity to abscisic acid in a sprouting-susceptible sorghum (Sorghum bicolor) variety is associated with altered ABA signaling. Seed Sci. Res. 2007, 17, 81–90. [Google Scholar] [CrossRef]
- Gubler, F.; Hughes, T.; Waterhouse, P.; Jacobsen, J. Regulation of dormancy in barley by blue light and after-ripening: Effects on abscisic acid and gibberellin metabolism. Plant Physiol. 2008, 147, 886–896. [Google Scholar] [CrossRef]
- Li, W.; Liu, X.; Khan, M.A.; Yamaguchi, S. The effect of plant growth regulators, nitric oxide, nitrate, nitrite and light on the germination of dimorphic seeds of Suaeda salsa under saline conditions. J. Plant Res. 2005, 118, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Halwas, S.J. Domesticating Chenopodium: Applying Genetic Techniques and Archaeological Data to Understanding Pre-Contact Plant Use. Ph.D. Thesis, Departments of Biological Sciences and Anthropology, University of Manitoba, MB, Canada, 2017. [Google Scholar]
- Yao, S.; Lan, H.; Zhang, F. Variation of seed heteromorphism in Chenopodium album and the effect of salinity stress on the descendants. Annu. Bot. 2010, 105, 1015–1025. [Google Scholar] [CrossRef]
- Vetch, J.; Stougaard, R.N.; Martin, J.M.; Giroux, M.J. Review: Revealing the genetic mechanisms of pre-harvest sprouting in hexaploidy wheat (Triticum aestivum L.). Plant Sci. 2019, 281, 180–185. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L. Bases of preharvest sprouting resistance in barley: Physiology, molecular biology and environmental control of dormancy in the barley grain. In Barley Science. Recent Advances from Molecular Biology to Agronomy of Yield and Quality; Slafer, G.A., Molina-Cano, J.L., Savin, R., Araus, J.L., Romagosa, I., Eds.; Food Product Press: New York, NY, USA, 2001; pp. 481–502. [Google Scholar]
- Farnsworth, E. The ecology and physiology of viviparous and recalcitrant seeds. Annu. Rev. Ecol. Syst. 2000, 31, 107–138. [Google Scholar] [CrossRef]
- Baker, H.G. Characteristics and Modes of Origin of Weeds. In The Genetics of Colonizing Species; Academic Press: Cambridge, MA, USA, 1965; pp. 147–168. [Google Scholar]
- Murphy, K.M.; Matanguihan, J.B.; Fuentes, F.F.; Gómez-Pando, L.R.; Jellen, E.N.; Maughan, P.J.; Jarvis, D.E. Quinoa breeding and genomics. Plant Breed. Rev. 2018, 42, 257–320. [Google Scholar]
- Peroni, P.A. Field and laboratory investigations of seed dormancy in red maple (Acer rubrum L.) from the North Carolina piedmont. For. Sci. 1995, 41, 378–386. [Google Scholar]
- Ellis, R.H.; Hong, T.D.; Roberts, E.H. A low-moisture-content to logarithmic relations between seed moisture content and longevity. Annu. Bot. 1988, 61, 405–408. [Google Scholar] [CrossRef]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–10761. [Google Scholar] [CrossRef]
- Kermode, A.R. Role of abscisic acid in seed dormancy. J. Plant Growth Regul. 2005, 24, 319–344. [Google Scholar] [CrossRef]
- Berjak, P. Unifying perspectives of some mechanisms basic to desiccation tolerance across life forms. Seed Sci. Res. 2006, 16, 1–15. [Google Scholar] [CrossRef]
- Nonogaki, M.; Nonogaki, H. Prevention of preharvest sprouting through hormone engineering and germination recovery by chemical biology. Front. Plant Sci. 2017, 8, 90. [Google Scholar] [CrossRef]
- Dorne, A.J. Variation in seed germination inhibition of Chenopodium bonus-henricus in relation to altitude of plant growth. Can. J. Bot. 1981, 59, 1893–1901. [Google Scholar] [CrossRef]
- Crocker, W.; Barton, L.V. Physiology of Seeds; Chronica Bot. Co.: Waltham, MA, USA, 1957; p. 267. [Google Scholar]
- Heydecker, W. Glossary of terms. In Seed Ecology; Heydecker, W., Ed.; Butterworths: Oxford, UK; Pennsylvania State University Press: University Park, PA, USA, 1973; pp. 553–557. [Google Scholar]
- Ceccato, D.; Bertero, D.; Batlla, D.; Galati, B. Structural aspects of dormancy in quinoa (Chenopodium quinoa): Importance and possible action mechanisms of the seed coat. Seed Sci. Res. 2015, 25, 267–275. [Google Scholar] [CrossRef]
- Metzger, R.J.; Silbaugh, B.A. Location of genes for seed coat color in hexaploid wheat Triticum aestivum L. Crop Sci. 1970, 10, 495–496. [Google Scholar] [CrossRef]
- Himi, E.; Yamashita, Y.; Haruyama, N.; Yanagisawa, T.; Maekawa, M.; Taketa, S. Ant28 gene for proanthocyanidin synthesis encoding the R2R3 MYB domain protein (Hvmyb10) highly affects grain dormancy in barley. Euphytica 2012, 188, 141–151. [Google Scholar] [CrossRef]
- Himi, E.; Mares, D.J.; Yanagiswa, A.; Noda, K. Effect of grain colour gene (R) on grain dormancy and sensitivity of the embryo to abscisic acid (ABA) in wheat. J. Exp. Bot. 2002, 53, 1569–1574. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Cai, S.; Wang, S.; Liu, S.; Zhang, G.; Bai, G. Genotyping-by-sequencing (GBS) identified SNP tightly linked to QTL for preharvest sprouting resistance. Theor. Appl. Gen. 2015, 128, 1385–1395. [Google Scholar] [CrossRef] [PubMed]
- Jacques, R. Action de la lumiere par l’intermediaire du phytochrome sur la germination, la croissance et le developpement de Chenopodium polyspermum L. Physiol. Veg. 1968, 6, 137–164. [Google Scholar]
- Karssen, C.M. The light promoted germination of the seeds of Chenopodium album L. III. Effect of the photoperiod during growth and development of the plants on the dormancy of the produced seeds. Acta Bot. Neerl. 1970, 19, 81–94. [Google Scholar] [CrossRef]
- Pourrat, Y.; Jacques, R. The influence of photoperiodic conditions received by the mother plant on morphological and physiological characteristics of Chenopodium polyspermum L. seeds. Plant Sci. Lett. 1975, 4, 273–279. [Google Scholar] [CrossRef]
- Jacobsen, S.E.; Bach, A.P. The influence of temperature on seed germination rate in quinoa (Chenopodium quinoa Willd). Seed Sci. Tech. 1998, 26, 515–523. [Google Scholar]
- Bois, J.F.; Winkel, T.; Lhomme, J.P.; Raffaillac, J.P.; Rocheteau, A. Response of some Andean cultivars of quinoa (Chenopodium quinoa Willd.) to temperature: Effects on germination, phenology, growth and freezing. Eur. J. Agron. 2006, 25, 299–308. [Google Scholar] [CrossRef]
- Präger, A.; Munz, S.; Nkebiwe, P.M.; Mast, B.; Graeff-Hönninger, S. Yield and quality characteristics of different quinoa (Chenopodium quinoa willd.) cultivars grown under field conditions in southwestern Germany. Agronomy 2018, 8, 197. [Google Scholar] [CrossRef]
- Williams, J.T.; Harper, J.L. Seed polymorphism and germination: I. The influence of nitrates and low temperatures on the germination of Chenopodium album. Weed Res. 1965, 5, 141–150. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L.; Fenner, M.; Edwards, P.J. Changes in germinability, ABA content and ABA embryonic sensitivity in developing seeds of Sorghum bicolor L. Moench. induced by water stress during grain filling. New Phytol. 1991, 118, 339–347. [Google Scholar] [CrossRef]
- Marques, A.; Nijveen, H.; Somi, C.; Ligterink, W.; Hilhorst, H. Induction of desiccation tolerance in desiccation sensitive Citrus limon seeds. J. Integr. Plant Biol. 2019, 61, 624–638. [Google Scholar] [CrossRef]
- Bendevis, M.A.; Sun, Y.; Shabala, S.; Rosenqvist, E.; Liu, F.; Jacobsen, S.E. Differentiation of photoperiod-induced ABA and soluble sugar responses of two quinoa (Chenopodium quinoa Willd.) cultivars. J. Plant Growth Red. 2014, 33, 562–570. [Google Scholar] [CrossRef]
- Ceccato, D.V.; Bertero, H.D.; Batlla, D. Environmental control of dormancy in quinoa (Chenopodium quinoa) seeds: Two potential genetic resources for preharvest sprouting tolerance. Seed Sci. Res. 2011, 21, 133–141. [Google Scholar] [CrossRef]
- Gualano NABenech-Arnold, R.L. The effect of water and nitrogen availability during grain filling on the timing of dormancy release in malting barley crops. Euphytica 2009, 168, 291–301. [Google Scholar] [CrossRef]
- Benech-Arnold, R.L. Inception, maintenance and termination of dormancy in grain crops: Physiology, genetics and environmental control. In Handbook of Seed Physiology: Applications to Agriculture; Benech-Arnold, R.L., Sanchez, R.A., Eds.; The Haworth Press: New York, NY, 2004; pp. 169–198. [Google Scholar]
- Gubler, F.; Millar, A.A.; Jacobsen, J.V. Dormancy release, ABA and preharvest sprouting. Plant Biol. 2005, 8, 183–187. [Google Scholar]
- Gómez, M.B.; Castro, P.A.; Mignone, C.; Bertero, H.D. Can yield potential be increased by manipulation of reproductive partitioning in quinoa (Chenopodium quinoa)? Evidence from gibberellic acid synthesis inhibition using Paclobutrazol. Funct. Plant Biol. 2011, 38, 420–430. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds—Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Martinez, S.A.; Godoy, J.; Huang, M.; Zhang, Z.; Carter, A.H.; Garland Campbell, K.A.; Steber, C.M. Genome-wide association mapping for tolerance to preharvest sprouting and low falling numbers in wheat. Front. Plant Sci. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Fofana, B.; Humphreys, D.G.; Rasul, G.; Cloutier, S.; Brûlé-Babel, A.; Woods, S.; Somers, D.J. Mapping quantitative trait loci controlling preharvest sprouting resistance in a red× white seeded spring wheat cross. Euphytica 2009, 165, 509–521. [Google Scholar] [CrossRef]
- Hauvermale, A.L.; Sanad, M.N.M.E. Phenological plasticity of wild and cultivated plants. In Tech Open. Plant Phenology; IntechOpen Limited: London, UK, 2019; ISBN 978-953-51-5871-4. [Google Scholar]
- Jellen, E.N.; Jarvis, D.E.; Hunt, S.P.; Mangelsen, H.H.; Maughan, P.J. New seed collections of North American pitseed goosefoot (Chenopdium berlandieri) and efforts to identify its diploid ancestors through whole-genome sequencing. Int. J. Agric. Nat. Res. 2019, 46, 187–196. [Google Scholar]
- Souza, F.F.D.J.; Devilla, I.A.; de Souza, R.T.; Teixeria, I.R.; Spehar, C.R. Physiological quality of quinoa seeds submitted to different storage conditions. Afr. J. Agric. Res. 2016, 11, 1299–1308. [Google Scholar]
- Beadmoore, T.; Whittle, C.A. Induction of tolerance to desiccation and cryopreservation in silver maple (Acer saccharinum) embryonic axes. Tree Physiol. 2005, 25, 965–972. [Google Scholar] [CrossRef]
- Pilati, S.; Bagagli, G.; Sonego, P.; Moretto, M.; Brazzale, D.; Castorina, G.; Simoni, L.; Tonelli, C.; Guella, G.; Engelen, K.; et al. Abscisic acid is a major regulator of grape berry ripening onset: New insights into ABA signaling. Front. Plant Sci. 2017, 21, 1–16. [Google Scholar] [CrossRef]



{kind=link}
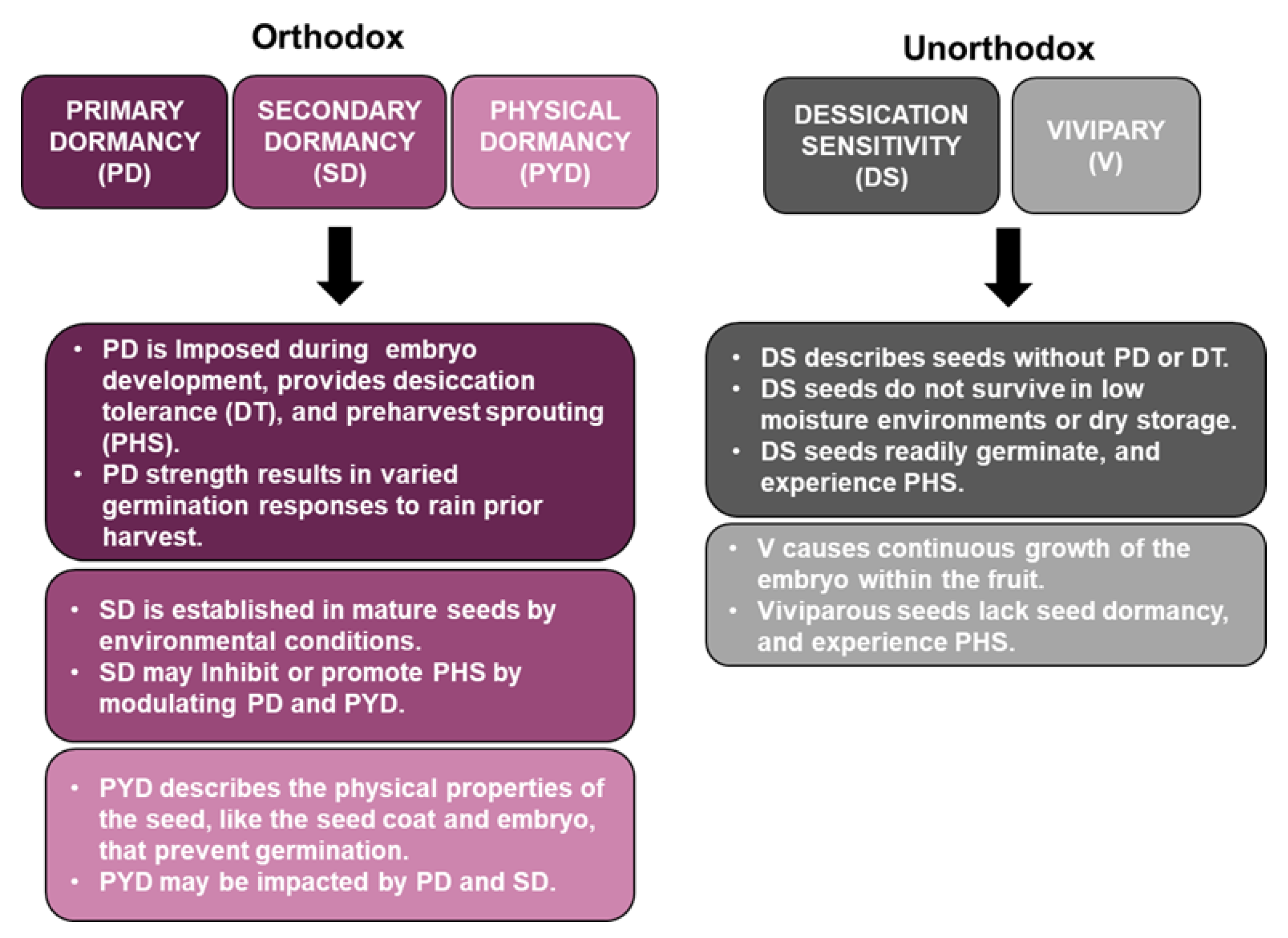{kind=link}
| Dormancy Category | Description |
|---|---|
| Primary | Established during embryo maturation by the plant hormone abscisic acid (ABA) and prevent germination. |
| Secondary | Established in mature seeds by environment stimuli preventing germination. |
| Physiological | Physiological responses to environmental or hormonal stimuli that prevent germination. |
| Morphological | Fully differentiated embryos remain physically too small to carry out radicle/cotyledon emergence. |
| Morphophysiological | Physiological responses and physical limitations that prevent germination. |
| Physical | Specialized physical features of the seed that prevent germination. |
| Combinational | Specialized physical features and physiological responses that prevent germination. |
| Embryo | External or internal physical or biochemical signals that prevent embryo growth and germination. |
| Seed Coat-imposed | Imposed by a hard, impermeable seed coat requiring physical damage to induce germination. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McGinty, E.M.; Murphy, K.M.; Hauvermale, A.L. Seed Dormancy and Preharvest Sprouting in Quinoa (Chenopodium quinoa Willd). Plants 2021, 10, 458. https://doi.org/10.3390/plants10030458
McGinty EM, Murphy KM, Hauvermale AL. Seed Dormancy and Preharvest Sprouting in Quinoa (Chenopodium quinoa Willd). Plants. 2021; 10(3):458. https://doi.org/10.3390/plants10030458
Chicago/Turabian StyleMcGinty, Emma M., Kevin M. Murphy, and Amber L. Hauvermale. 2021. "Seed Dormancy and Preharvest Sprouting in Quinoa (Chenopodium quinoa Willd)" Plants 10, no. 3: 458. https://doi.org/10.3390/plants10030458
APA StyleMcGinty, E. M., Murphy, K. M., & Hauvermale, A. L. (2021). Seed Dormancy and Preharvest Sprouting in Quinoa (Chenopodium quinoa Willd). Plants, 10(3), 458. https://doi.org/10.3390/plants10030458