Potassium Control of Plant Functions: Ecological and Agricultural Implications



{kind=link}
{kind=link}
Abstract
1. Introduction
2. Stable Cation in Solution Necessary for Plant Functional Homeostasis and Production Control
2.1. The Biogeochemical Properties of K+
2.2. The Multiple Functions of K+ in Plants
3. Plant K Uptake Mechanisms
3.1. The Role of Transporters and Channels
3.2. Plant Morphology and Ecological Aspects in Plant K+ Uptake
4. Internal Transport of K+, Water, Nutrients, and Biomolecules
4.1. The Control of Internal Water Transport
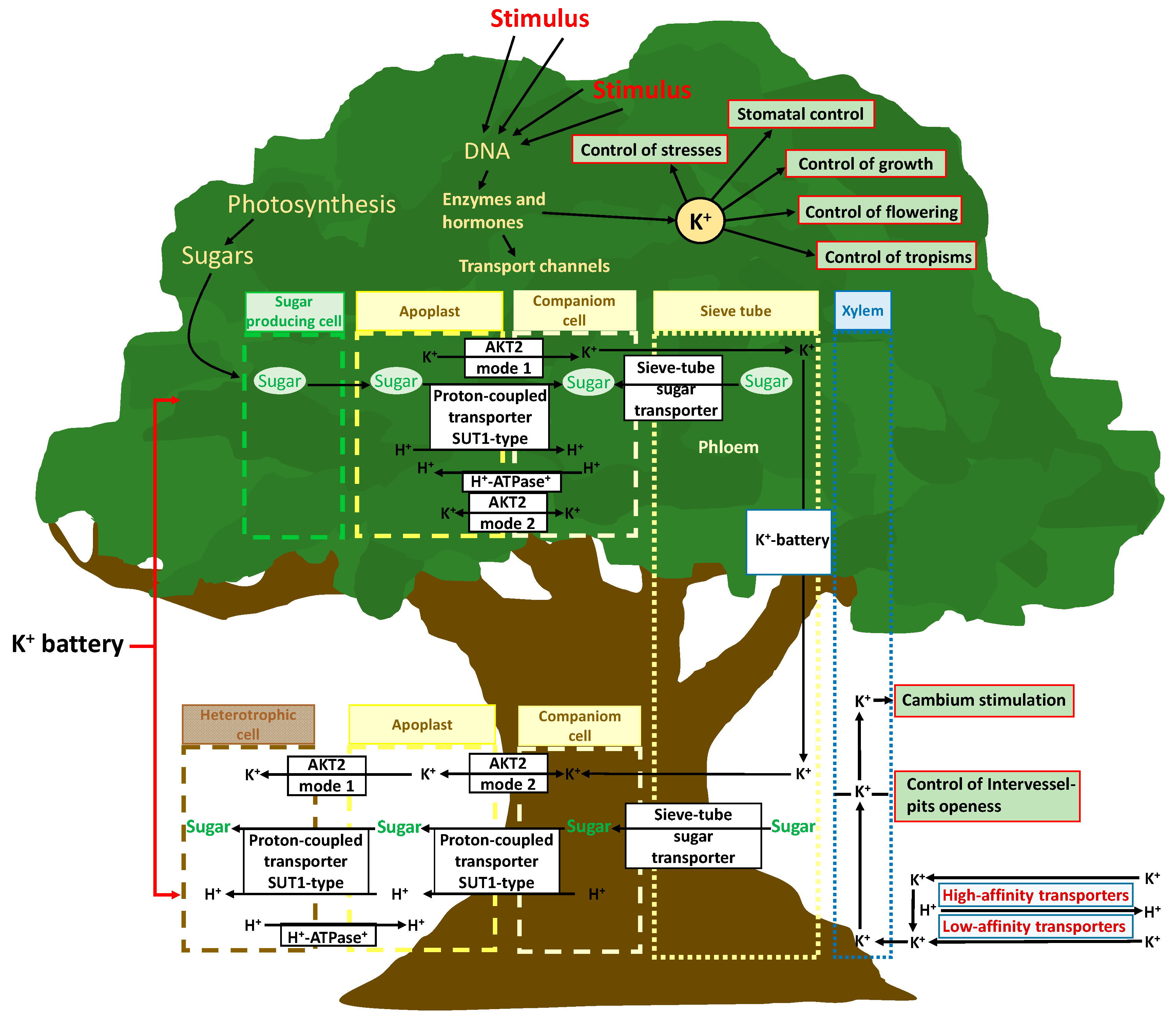
4.2. The K+ Role in the Complex Mechanisms of Internal Biomolecules, Nutrients, and Energy in Plants: “The Potassium Battery”
5. Role of K in Plant Responses to External Environmental Conditions
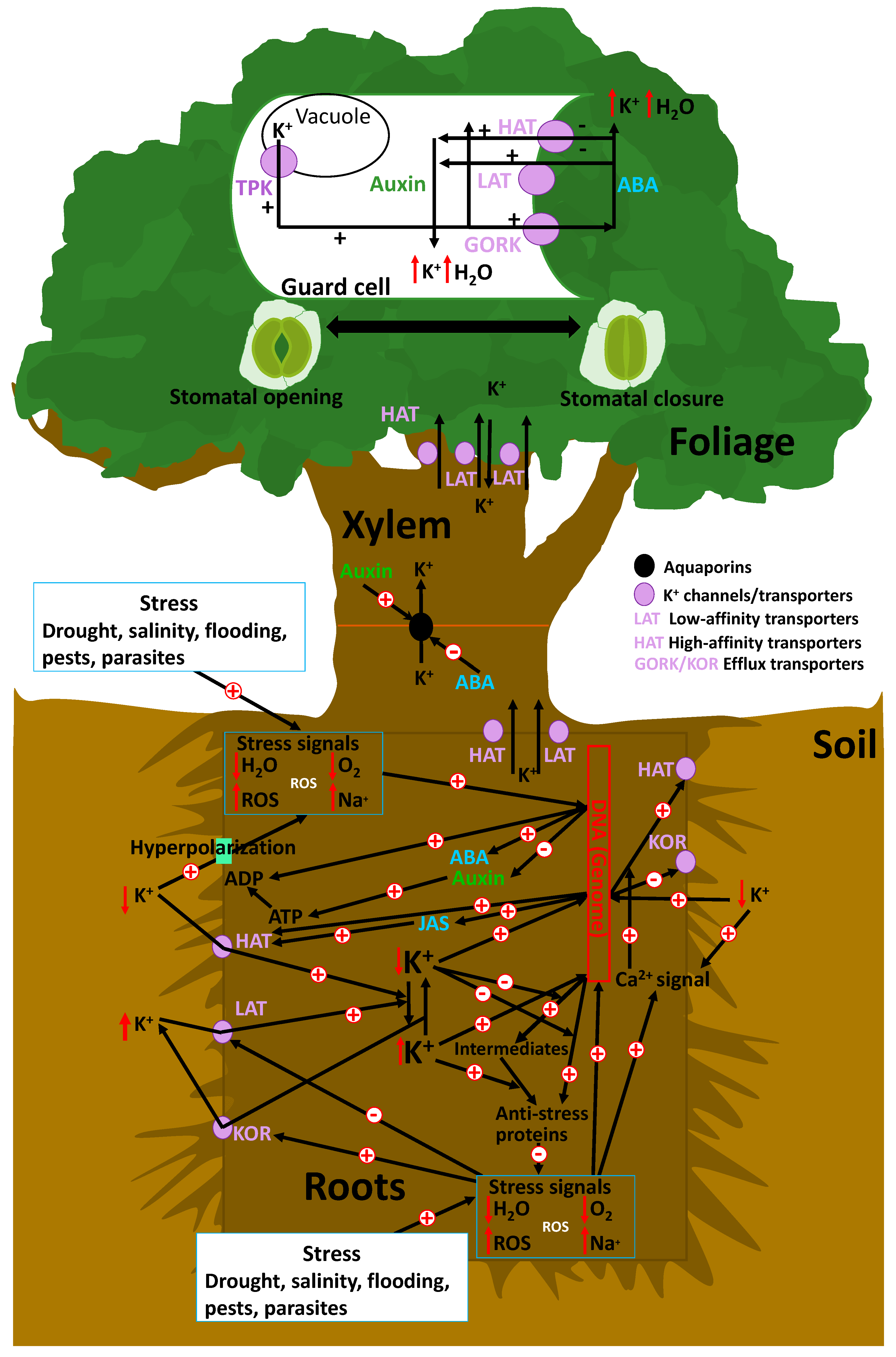
5.1. Abiotic Stress
5.2. Biotic Stress
6. Role of K in Terrestrial Ecosystems
6.1. Ecological Aspects of the K+ Cycle in the Plant–Soil System
6.2. K+’s Role in Plant–Soil System Shifts under Distinct Abiotic and Biotic Environmental Circumstances
6.3. Human Impacts on the K+ Potassium Cycle
7. K and Food Security and Human Health
7.1. K Fertilizers and Food Security
7.2. K Fertilization, Food Potassium Content, and Human Health
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gallardo, J.F.; Martín, A.; Moreno, G. Nutrient efficiency and resorption in Quercus pyrenaica oak coppices under different rainfall regimes of the Sierra de Gata mountains (central western Spain). Ann. For. Sci. 1999, 56, 321–331. [Google Scholar] [CrossRef]
- Trémolières, M.; Schnitzler, A.; Sánchez-Pérez, J.M.; Schmitt, D. Changes in foliar nutrient content and resorption in Fraxinus excelsior L., Ulmus minor Mill. and Clematis vitalba L. after prevention of floods. Ann. For. Sci. 1999, 56, 641–650. [Google Scholar] [CrossRef]
- Vergutz, L.; Manzoni, S.; Porporato, A.; Novais, R.F.; Jackson, R.B. Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol. Monogr. 2012, 82, 205–220. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium: A neglected nutrient in global change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef]
- Tripler, C.E.; Kaushal, S.S.; Linkens, G.E.; Walter, M.T. Patterns in potassium dynamics in forest ecosystems. Ecol. Let. 2006, 9, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Tränker, M.; Tavakol, A.; Jákli, B. Functioning of potassium and magnesium in photosynthesis, photosynthate translocation and photoprotection. Physiol. Plant. 2018, 163, 414–431. [Google Scholar]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in root growth and development. Plants 2019, 8, 435. [Google Scholar] [CrossRef]
- Dreyer, J.; Gomez-Porras, J.L.; Riedelsberger, J. The potassium battery: A mobile energy source for transport processes in plant vascular tissues. New Phytol. 2017, 216, 1049–1053. [Google Scholar] [CrossRef]
- Jia, Q.; Zheng, C.; Sun, S.; Amjad, H.; Liang, K.; Lin, W. The role of plant cation/proton antiporter gene family in salt tolerance. Biol. Plant 2018, 62, 617–629. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Shankar, A.; CHandran, A.K.N.; Sharma, M.; Jung, K.H.; Suprasanna, P.; Pandey, G.K. Emerging concepts of potassium homeostasis in plants. J. Exp. Bot. 2019, 71, 608–619. [Google Scholar] [CrossRef]
- Cuin, T.A.; Dreyer, I.; Michard, E. The role of potassium channels in Arabidopsis thaliana long distance electrical signaling: AKT2 modulates tissue excitability while GORK shapes action potentials. Int. J. Mol. Sci. 2018, 19, 926. [Google Scholar] [CrossRef]
- Locascio, A.; Marques, M.C.; Garcia-Martinez, G.; Corratge-Faillie, C.; Andres-Colas, N.; Rubio, L.; Fernandez, J.A.; Very, A.A.; Mulet, J.M.; Yenush, L. BCL2-associated athanogene 4 regulates the KAT1 potassium cannel and controls stomatal movement. Plant Physiol. 2019, 181, 1277–1294. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, P.; Abdel Latef, A.A.; Abd-Allah, E.F.; Hashem, A.; Sarwat, M.; Anjum, N.A.; Gucel, S. Calcium and potassium supplementation enhanced growth, osmolity secondary metabolite production, and enzymatic antioxidant machinery in Cadmium-exposed chickpea (Cicer arietinum L). Front. Plant Sci. 2016, 7, 513. [Google Scholar] [CrossRef]
- Abass, M.; Tomar, N.S.; Tttal, M.; Argal, S.; Agarwal, R.M. Plant growth under water/salt stress: ROS production; antioxidants and significant of added potassium under such conditions. Physiol. Mol. Biol. Plants 2017, 23, 731–744. [Google Scholar]
- Ahanger, M.A.; Agarwal, R.M.; Tomar, N.S.; Shrivastava, M. Potassium induces positive changes in nitrogen metabolism and antioxidant system of oat (Avena sativa L. cultivar Kent). J. Plant Int. 2015, 10, 211–223. [Google Scholar]
- Chen, G.; Li, C.L.; Gao, Z.Y.; Zhang, Y.; Zhu, L.; Hu, J.; Ren, D.Y.; Xu, G.H.; Qian, Q. Driving the expression of RAA1 with a drought-responsive promoter enhances root growth in rice, its accumulation of potassium and its tolerance to moisture stress. Environ. Exp. Bot. 2018, 147, 147–156. [Google Scholar] [CrossRef]
- Leigh, R.R.; Wyn Jones, R.G. A hypothesis relating critical potassium concentrations for growth to the distribution and functions of this ion in plant cell. New Phytol. 1984, 97, 1–13. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Potassium and potassium-permeable channels in plant salt tolerance. In Ion Channels and Plant Stress Responses; Demidchik, V., Maathuis, F., Eds.; Springer: Basel, Switzerland, 2010; pp. 87–110. [Google Scholar]
- White, P.; Karley, A. Potassium. In Cell Biology of Metals and Nutrients; Hell, R., Mendel, R.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 199–224. [Google Scholar]
- Dreyer, I.; Uozumi, N. Potassium channels in plant cells. FEBS J. 2011, 278, 4293–4303. [Google Scholar] [CrossRef]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Martínez, V.; Benito, B.; Rubio, F. Comparison between Arabidopsis and rice for main pathways of K+ and Na+ uptake by roots. Front. Plant Sci. 2016, 7, 992. [Google Scholar] [CrossRef]
- Hawkesford, M.; Horst, W.; Kichey, T.; Lambers, H.; Schjoerring, J.; Skrumsager Moller, I.; White, P. Function of macronutrients. In Marschner’s Mineral Nutrition of Higher Plants; Marschner, P., Ed.; Academic Press: London, UK, 2012; pp. 135–189. [Google Scholar]
- Ward, J.M.; Schroeder, J.I. Calcium-activated K+ channels and calcium-induced calcium release by slow vacuolar ion channels in guard cell vacuoles implicated in the control of stomatal closure. Plant Cell 1994, 6, 669–683. [Google Scholar] [CrossRef]
- Ng, C.K.Y.; Mcainsh, M.R.; Gray, J.E.; Hunt, L.; Leckie, C.P.; Mills, L.; Hetherington, A.M. Calcium-based signaling systems in guard cells. New Phytol. 2001, 151, 109–120. [Google Scholar] [CrossRef]
- Wang, Y.M.; Wu, W.H. Potassium transport and signaling in higher plants. Ann. Rev. Plant Biol. 2013, 64, 451–476. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef]
- Arteca, R.N. Introduction to Horticultural Science, 2nd ed.; Cengage Learning: Stamford, CT, USA, 2016; p. 611. [Google Scholar]
- Akenga, P.; Salim, A.; Onditi, A.; Yusuf, A.; Waudo, W. Determination of selected micro and macronutrients in sugarcane growing soils at Kakamega north district, Kenya. IOSR J. Appl. Chem. (IOSR-JAC) 2014, 7, 34–41. [Google Scholar] [CrossRef]
- Marcus, Y. Ionic Radii in Aqueous Solutions. Chem. Rev. 1988, 88, 1475–1498. [Google Scholar] [CrossRef]
- Russo, N.; Toscano, M.; Grand, A. Bond energies and attachments sites of sodium and potassium cations to DNA and RNA nucleic acid bases in gas phase. J. Am. Chem. Soc. 2001, 123, 10272–10279. [Google Scholar] [CrossRef] [PubMed]
- Persson, I. Hydrated model ions in aqueous solution: How regular are their structures? Pure Appl. Chem. 2010, 82, 1901–1917. [Google Scholar] [CrossRef]
- Gajdanowicz, P.; Michard, E.; Sandmann, M.; Rocha, M.; Guedes Corrêa, L.G.; Ramírez-Aguilar, S.J.; Gómez-Porras, J.L.; González, W.; Thibaud, J.B.; van Dongen, J.T. Potassium (K+) gradients serve as a mobile energy source in plant vascular tissues. Proc. Natl. Acad. Sci. USA 2011, 108, 864–869. [Google Scholar] [CrossRef]
- Langer, K.; Ache, P.; Geiger, D.; Stinzing, A.; Arend, M.; Wind, C.; Regan, S.; Fromm, J.; Hedrich, R. Poplar potassium transporters capable of controlling K+ homeostasis and K+-dependent xylogenesis. Plant J. 2002, 32, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Amtmann, A.; Troufflard, S.; Armengaud, P. The effect of potassium nutrition on pest and disease resistance in plants. Physiol. Plant 2008, 133, 682–691. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Sardans, J.; Pérez-Trujillo, M.V.; Estiarte, M.; Peñuelas, P. Drought enhances folivory by shifting foliar metabolomes in Quercus ilex trees. New Phytol. 2014, 202, 874–885. [Google Scholar] [CrossRef]
- Nissen, P. The structural basis of ribosome activity in peptide bond synthesis. Science 2000, 289, 920–930. [Google Scholar] [CrossRef]
- Andrés, Z.; Pérex-Hormaeche, J.P.; Leidi, E.O.; Schlücking, K.; Steinhorst, L.; McLachlan, D.H.; Schumacher, K.; Hetherington, A.M.; Kudla, J.V.; Cubero, B.; et al. Control of vacuolar dynamics and regulation of stomatal aperture to tonoplast potassium uptake. Proc. Natl. Acad. Sci. USA 2014, 111, 1806–1814. [Google Scholar] [CrossRef]
- Carroll, M.J.; Slaughter, L.H.; Krouse, J.M. Turgor potential and osmotic constituents of Kentucky Bluegrass leaves supplied with four levels of potassium. Agron. J. 1994, 86, 1079. [Google Scholar] [CrossRef]
- Jia, Y.B.; Yang, X.E.; Feng, Y.; Jilani, G. Differential response of root morphology to potassium deficient stress among rice genotypes varying in potassium efficiency. J. Zhejiang Uni. Sci. B 2008, 9, 427–434. [Google Scholar] [CrossRef]
- Jákli, B.; Tavakol, E.; Tränkner, M.; Senbayram, M.; Dittert, K. Quantitative limitations to photosynthesis in K deficient sunflower and their implications on water-use efficiency. J. Plant Physiol. 2017, 209, 20–30. [Google Scholar] [CrossRef]
- Zahoor, R.M.; Zhao, W.Q.; Dong, H.R.; Snider, J.L.; Abid, M.; Iqbal, B.; Zhou, Z.G. Potassium improves photosynthetic tolerance to and recovery from episodic drought stress in functional leaves of cotton (Gossypium hirsutum L.). Plant Physiol. Biochem. 2017, 119, 21–32. [Google Scholar] [CrossRef]
- Austin, J.; First, E.A. Potassium functionally replaces the second lysine of the KMSKS signature sequence in human tyrosyl-tRNA synthetase. J. Biol. Chem. 2002, 277, 20243–20248. [Google Scholar] [CrossRef]
- Leigh, R.A. Potassium homeostasis and membrane transport. J. Plant Nutr. Soil Sci. 2001, 164, 193–198. [Google Scholar] [CrossRef]
- Fricke, W.; Leigh, R.A.; Tomos, A.D. Concentrations of inorganic and organic solutes in extracts from individual epidermal, mesophyll and bundle sheath cells of barley leaves. Planta 1994, 192, 310–316. [Google Scholar] [CrossRef]
- Gonzales, E.M.; Arrese-Igor, C.; Aparicio-Tejo, P.M.; Royuela, M.; Koyro, H.W. Solute heterogeneity and osmotic adjustment in different leaf structures of semi-leafless pea (Pisum sativum L.) subjected to water stress. Plant Biol. 2002, 4, 558–566. [Google Scholar] [CrossRef]
- Shabala, S.; Babourina, O.; Newman, I. Ion-specific mechanisms of osmoregulation in bean mesophyll cells. J. Exp. Bot. 2000, 51, 1243–1253. [Google Scholar] [CrossRef]
- Cuin, T.A.; Miller, A.J.; Laurie, S.A.; Leigh, R.A. Potassium activities in cell compartments of salt-grown barley leaves. J. Exp. Bot. 2003, 54, 657–661. [Google Scholar] [CrossRef]
- Nio, S.A.; Cawthray, G.R.; Wade, L.J.; Colmer, T.D. Pattern of solutes accumulated during leaf osmotic adjustment as related to duration of water deficit for wheat at the reproductive stage. Plant Physiol. Biochem. 2011, 49, 1126–1137. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, S.; Uozumi, N. Organelle-localized potassium transport systems in plants. J. Plant Physiol. 2014, 171, 743–747. [Google Scholar] [CrossRef]
- Lu, Z.F.; Lu, J.W.; Pan, Y.H.; Li, X.K.; Cong, R.H.; Ren, T. Genotypic variation in photosynthetic limitation responses to K deficiency of Brassica napus is associated with potassium utilization efficiency. Funct. Plant Biol. 2016, 42, 880–891. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant. 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Rodríguez-Navarro, A.; Rubio, F. High-affinity potassium and sodium transport systems in plants. J. Exp. Bot. 2006, 57, 1149–1160. [Google Scholar] [CrossRef]
- Szczerba, M.W.; Britto, D.T.; Kronzucker, H.J. K+ transport in plants: Physiology and molecular biology. J. Plant Physiol. 2009, 166, 447–466. [Google Scholar] [CrossRef]
- Rubio, F.; Alemán, F.; Nieves-Cordones, M.; Vicente, M. Studies on Arabidopsisathak5, atakt1 double mutants disclose the range of concentrations at which AtHAK5, AtAKT1 and unknown systems mediate K+ uptake. Physiol. Plant. 2010, 139, 220–228. [Google Scholar] [CrossRef]
- Pottosin, I.; Dobrovinskaya, O. Non-selective cation channels in plasma and vacuolar membranes and their contribution to K+ transport. J. Plant Physiol. 2014, 171, 732–742. [Google Scholar] [CrossRef]
- Martínez-Cordero, M.A.; Martínez, V.; Rubio, F. Cloning and functional characterization of the high-affinity K+ transporter HAK1 of pepper. Plant Mol. Biol. 2004, 56, 413–421. [Google Scholar] [CrossRef]
- Cai, K.F.; Gao, H.Z.; Wu, X.J.; Zhang, S.; Han, Z.G.; Chen, X.H.; Zhang, G.P.; Zeng, F.R. The ability to regulate transmembrane potassium transport in root is critical for drought tolerance in barley. Int. J. Mol. Sci. 2019, 20, 4111. [Google Scholar] [CrossRef] [PubMed]
- Rubio, F.; Santa-Maria, G.E.; Rodríguez-Navarro, A.A. Cloning of Arabidopsis and barley cDNAs encoding HAK potassium transporters in root and shoot cells. Physiol. Plant. 2000, 109, 34–43. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Martinez-Cordero, M.A.; Martinez, V.; Rubio, F. An NH4+-sensitive component dominates high-affinity K+ uptake in tomato plants. Plant Sci. 2007, 172, 273–280. [Google Scholar] [CrossRef]
- Kavka, M.; Polle, A. Phosphate uptake kinetics and tissue-specific transporter expression profiles in poplar (Populus x canescens) at different phosphorus availabilities. BMC Plant Biol. 2016, 16, 206. [Google Scholar] [CrossRef]
- Kim, E.J.; Kwak, J.M.; Uozumi, N.; Schroeder, J.I. AtKUP1: An Arabidopsis gene encoding high-affinity potassium transport activity. Plant Cell 1998, 10, 51–62. [Google Scholar] [CrossRef]
- Hamamoto, S.; Horie, T.; Hauser, F.; Deinlein, U.; Schoroeder, J.I.; Uozumi, N. HKT transportes mediate salt stress resistance in plants: From structure and function to the field. Cur. Opin. Biotech. 2015, 32, 113–120. [Google Scholar] [CrossRef]
- Britto, D.T.; Kronzucker, H.J. Nitrogen acquisition, PEP carboxylase, and cellular pH homeostasis: New views on old paradigms. Plant Cell Environ. 2005, 28, 183–188. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Dabravolski, S.A.; Pan, T.M.; Shabala, S. Phylogenetic diversity and physiological roles of plant monovalent cation/H+ antiporters. Front. Plant Sci. 2020, 11, 573564. [Google Scholar] [CrossRef] [PubMed]
- Guerth, M.; Mäser, P. Potassium transporters in plants-involvement in K+ acquisition, redistribution and homeostasis. FEBS Lett. 2007, 581, 5355–5360. [Google Scholar] [CrossRef] [PubMed]
- Maathius, F.J.M.; Filatov, V.; Herzyk, P.; Krijger, G.C.; Axelsen, K.B.; Chen, S.; Green, B.J.; Li, Y.; Madagan, K.L.; Sánchez-Fernández, R.; et al. Transcriptomic analysis of root transporters reveals participation of multiple gene families in the response to cation stress. Plant J. 2003, 35, 675–692. [Google Scholar] [CrossRef] [PubMed]
- Padmanaban, S.; Czerny, D.D.; Levin, K.A.; Leydon, A.R.; Su, R.T.; Maugel, T.K.; Zou, Y.; Chanroj, S.; Cheung, A.Y.; Johnson, M.A.; et al. Transporters involved in pH and K+ homeostasis affect pollen wall formation, male fertility, and embrio development. J. Exp. Bot. 2017, 68, 3165–3178. [Google Scholar] [CrossRef]
- Qu, Y.; Guan, R.M.; Bose, J.; Henderson, S.W.; Wege, S.; Qiu, L.J.; Gilliham, M. GmSALT3 confers shoot Na+ and Cl- exclusion in soybean via two distinct processes. bioRxiv 2020, 2020, 896456. [Google Scholar]
- Rubio, F.; Nieves-Cordones, M.; Horie, T.; Shabala, S. Doing business as usual comes with a cost: Evaluating energy cost of maintaining plant intracellular K+ homeostasis under saline conditions. New Phytol. 2020, 225, 1097–1104. [Google Scholar] [CrossRef]
- Aranda-Sicilia, M.N.; Cagnac, O.; CHanroj, S.; Sze, H.; Rodríguez-Rosales, M.P.; Venema, K. Arabidopsis KEA2, a homolog of bacterial KefC, encodes a K+/H+ antiporter with a chloroplast transit peptide. Biochem. Biophys. Acta 2012, 1818, 2362–2371. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Pan, T.; Zhang, X.; Fan, L.; Quintero, F.J.; Zhao, H.; Su, X.; Li, X.; Vilalta, I.; Mendoza, I.; et al. K+ efflux antiporters 4, 5, and 6 mediate pH and K+ homeostasis in endomembrane compartments. Plant Physiol. 2018, 178, 1657–1678. [Google Scholar] [CrossRef]
- Wang, T.; Tang, R.J.; Yang, X.; Zheng, X.; Shao, Q.; Tang, Q.L.; Fu, A.; Luan, S. Golgi-localized cation/proton exchangers regulate ionic homeostasis and skotomorphogenesis in Arabidopsis. Plant Cell Environ. 2019, 42, 673–687. [Google Scholar] [CrossRef] [PubMed]
- Amtmann, A.; Blatt, M.R. Regulation of macronutrient transport. New Phytol. 2008, 181, 35–52. [Google Scholar] [CrossRef]
- Sharma, T.; Dreyer, I.; Riedelsberger, J. The role of K+ channels in uptake and redistribution of potassium in the model plant Arabidopsis thaliana. Front. Plant Sci. 2003, 4, 224. [Google Scholar] [CrossRef]
- Ragel, P.; Raddatz, N.R.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ nutrition in plants. Front. Plant Sci. 2019, 10, 281. [Google Scholar]
- Shrivastava, I.; Guy, H. Inward Rectifier K+ channels. In Bacterial Ion Channels and Their Eukaryotic Homologs; Kubalski, A., Martinac, B., Eds.; ASM Press: Washington, DC, USA, 2005; pp. 123–132. [Google Scholar]
- Zhao, X.-h.; Qiu, H.; Wen, J.; Wang, X.; Du, Q.; Wang, J.; Wang, Q. Response of root morphology, physiology and endogenous hormones in maize (Zea mays L.) to potassium deficiency. J. Integr. Agric. 2016, 15, 785–794. [Google Scholar] [CrossRef]
- Garcia, K.; Ané, J.M. Polymorphic responses of Medicago truncatula accessions to potassium deprivation. Plant Signal. Behav. 2017, 12, e1307494. [Google Scholar] [CrossRef]
- Feng, H.; Tang, Q.; Cai, J.; Xu, B.; Xu, G.; Yu, L. Rice OsHAK16 functions in potassium uptake and translocation in shoot, maintaining potassium homeostasis and salt tolerance. Planta 2019, 250, 549–561. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Li, B.; Xiong, C.; Eneji, A.E.; Zhang, M.; Li, F.; Tian, X.; Li, Z. The cotton high-affinity K+ transporter, GhHAK5a, is essential for shoot regulation of K+ uptake in root under potassium deficiency. Plant Cell Physiol. 2019, 60, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Kellermeier, F.; Armengaud, P.; Seditas, T.J.; Danku, J.; Salt, D.E.; Amtmann, A. Analysis of the root system architecture of Arabidopsis provides a quantitative readout of crosstalk between nutritional signals. Plant Cell 2014, 26, 1480–1496. [Google Scholar] [CrossRef]
- Li, J.; Wu, W.H.; Wang, Y. Potassium channel AKT1 is involved in the auxin-mediated root growth inhibition in Arabidopsis response to low K+ stress. J. Int. Plant Biol. 2017, 59, 895–909. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Miller, A.J.; Aleman, F.; Martinez, V.; Rubio, F. A putative role for the plasma membrane potential in the control of the expression of the gene encoding the tomato high-affinity potassium transporter HAK5. Plant Mol. Biol. 2008, 68, 521–532. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, W.H. Plant sensing and signaling in response to K+-deficiency. Mol. Plant 2010, 3, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Shin, R.; Berg, R.H.; Schachtman, D.P. Reactive oxygenspecies and root hairs in Arabidopsis root response to nitrogen, phosphorus and potassium deficiency. Plant Cell Physiol. 2005, 46, 1350–1357. [Google Scholar] [CrossRef] [PubMed]
- Pitzschke, A.; Forzani, C.; Hirt, H. Reactive oxygen speciessignaling in plants. Antioxid. Redox Signal. 2006, 8, 1757–1764. [Google Scholar] [CrossRef]
- Demidchik, K.; Shabala, S.; Isayenkov, S.; Cuin, T.A.; Pottosin, I. Calcium transport across plant membranes: Mechanisms and function. New Phytol 2018, in press. [Google Scholar] [CrossRef]
- Pei, Z.M.; Murata, Y.; Benning, G.; Thomine, S.; Klusener, B.; Allen, G.J.; Grill, E.; Schroeder, J.I. Calcium channels activated by hydrogen peroxide mediate abscisic acid signaling in guard cells. Nature 2000, 406, 731–734. [Google Scholar] [CrossRef]
- Yang, T.; Poovaiah, B.W. Hydrogen peroxide homeostasis:activation of plant catalase by calcium/calmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef]
- Shin, R.; Schachtman, D.P. Hydrogen peroxide mediates plant root cell response to nutrient deprivation. Proc. Natl. Acad. Sci. USA 2004, 101, 8827–8832. [Google Scholar] [CrossRef]
- Zhao, J.; Cheng, N.H.; Motes, C.M.; Blancaflor, E.B.; Moore, M.; Gonzales, N.; Padmanaban, S.; Sze, H.; Ward, J.M.; Hirschi, K.D. AtCHX13 is a plasma membrane K+ transporter. Plant Physiol. 2008, 148, 796–807. [Google Scholar] [CrossRef]
- Li, Y.; Peng, L.R.; Xie, C.Y.; Shi, X.Q.; Dong, C.X.; Shen, Q.R.; Xu, Y.C. Genome-wide identification, characterization, and expression analyses of the HAK/KUP/KT potassium transporter gene family reveals their involvement in K+ deficient and abiotic stress responses in pear rootstock seedlings. Plant Growth Regul. 2018, 85, 187–198. [Google Scholar] [CrossRef]
- Lee, S.C.; Lan, W.Z.; Kim, B.G.; Li, L.; Cheong, Y.H.; Pandey, G.K.; Lu, G.; Buchanan, B.B.; Lua, S. A protein phosphorylation/dephospholiration network regulates a plant potassium channel. Proc. Natl. Acad. Sci. USA 2007, 104, 15959–15964. [Google Scholar] [CrossRef] [PubMed]
- Stephan, A.; Schroeder, J.I. Plant salt stress status is transmitted systemically via propagating calcium waves. Proc. Natl. Acad. Sci. USA 2014, 111, 6126–6127. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, D.; Basset, M.; Lepetit, M.; Conejero, G.; Gaymard, H.; Astruc, S.; Grignon, C. Tissue-specific expression of Arabidopsis AKT1 gene is consistent with a role in K+ nutrition. Plant J. 1996, 9, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, I.; Stolzle, S.; Ivashuikina, N.; Hedrich, R. Ricr K+ uptake channel OsAKT1 is sensitive to salt stress. Planta 2005, 221, 212–221. [Google Scholar] [CrossRef]
- Philippar, K.; Fuchs, I.; Luthen, H.; Hoth, S.; Bauer, C.S.; Haga, K.; Thiel, G.; Ljug, K.; Sandberg, G.; Böttger, M.; et al. Auxin-induced K+ channel expression represents an essential step in coleoptile growth and gravitropism. Proc. Natl. Acad. Sci. USA 1999, 96, 12186–12191. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, S.; Talke, I.; Ehrhardt, T.; Nast, G.; Müller-Röber, B. Characterization of SKT1, an inwardly rectifying potassium channel from potato, by heterologous expression in insect cells. Plant Physiol. 1998, 116, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Buschmann, P.H.; Vaidyanathan, R.M.; Gassmann, W.; Schroeder, J.I. enhancement of Na+ uptake currents, time-dependent inward-rectifying K+ channels currents, and K+ channel transcripts by K+ starvation in wheat root cells. Plant Physiol. 2000, 122, 1387–1397. [Google Scholar] [CrossRef]
- Lebaudy, A.; Véry, A.A.; Sentenac, H. K+ channel activity in plants: Genes, regulation and functions. FEBS Lett. 2007, 25, 2357–2366. [Google Scholar] [CrossRef]
- Hachez, C.; Veljanovski, V.; Reinhardt, H.; Guillaumot, D.; Vanhee, C.; Chaumont, F.; Batoko, H. The Arabidopsis stress-induced TSPO-related protein reduces cell-surface expression of the aquaporin PIP2;7 through protein-protein interactions and autophagic degradation. Plant Cell 2014, 26, 4974–4990. [Google Scholar] [CrossRef]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef] [PubMed]
- Maurel, C.; Verdoucq, L.; Luu, D.T.; Santoni, V. Plant aquaporins: Membrane channels with multiple integrated functions. Annu. Rev. Plant Biol. 2008, 59, 595–624. [Google Scholar] [CrossRef]
- Yue, X.; Zhao, X.Y.; Fei, Y.; Zhang, X. Correlation of aquaporins and transmembrane solute transporters revealed by genome-wide analysis developing maize leaf. Comp. Funct. Genom. 2012, 2012, 546930. [Google Scholar]
- Liu, H.Y.; Sun, W.N.; Su, W.A.; Tang, Z.C. Co-regulation of water channels and potassium channels in rice. Physiol. Plant. 2006, 128, 58–69. [Google Scholar] [CrossRef]
- Armengaud, P.; Breitling, R.; Amtmann, A. The potassium-dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling. Plant Physiol. 2004, 136, 2556–2576. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ding, L.; Gao, L.; Li, Y.; Shen, Q.; Guo, S. The interactions of aquaporins and mineral nutrients in higher plants. Int. J. Mol. Sci. 2016, 17, 1229. [Google Scholar] [CrossRef]
- Cuéllar, T.; Pascaud, F.; Verdeil, J.L.; Torregrosa, L.; Adam-Blondon, A.F.; Thibaud, J.B.; Sentenac, H.; Gaillard, I.A. Grapevine Shaker inward K+ channel activated by the calcineurin B-like calcium sensor 1-protein kinase CIPK23 network is expressed in grape berries under drought stress conditions. Plant J. 2010, 61, 58–69. [Google Scholar] [CrossRef]
- Favreau, B.; Denis, M.; Ployet, R.; Mounet, F.; Peireira de Silva, H.; Franceschini, L.; Laclau, J.P.; Labate, C.; Carrer, H. Distinct leaf transcriptomic response of water deficient Eucalyptus grandis submitted to potassium and sodium fertilization. PLoS ONE 2019, 14, e0218528. [Google Scholar] [CrossRef] [PubMed]
- Oddo, E.; Inzerillo, S.; La Bella, F.; Grisafi, F.; Salleo, S.; Nardini, A.; Goldstein, G. Short-term effectsof potassium fertilization on the hydraulic conductance of Laurus nobilis L. Tree Physiol. 2011, 31, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Liesche, J.; Windt, C.; Bohr, T.; Schulz, A.; Jensen, K.H. Slower phloem transport in gymnosperm trees can be attributed to higher sieve element resistance. Tree Physiol. 2015, 35, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Deeken, R.; Geiger, D.; Fromm, J.; Karoleva, O.; Ache, P.; Langenfeld-Heyser, R.; Sauer, N.; May, S.T.; Hedrich, R. Loss of the AKT2/3 potassium channel affects sugar loading into the phloem of Arabidopsis. Planta 2002, 216, 334–344. [Google Scholar] [CrossRef]
- Dreyer, I.; Blatt, M.R. What makes a gate? The ins and outs of Kv-live K+ channels in plants. Trends Plant Sci. 2009, 14, 383–390. [Google Scholar] [CrossRef]
- Jeschke, D.W.; Atkins, C.A.; Pate, J.S. Ion circulation via phloem and xylem between root and shoot of nodulated white lupin. J. Plant Physiol. 1985, 117, 319–330. [Google Scholar] [CrossRef]
- Sklodowski, K.; Riedelsberger, J.; Raddatz, N.; Riadi, G.; Caballero, J.; Chérel, I.; Schulze, W.; Graf, A.; Dreyer, I. The receptor-like pseudokinase MRH1 interacts with the voltage-gated potassium channel AKT2. Sci. Rep. 2017, 7, 44611. [Google Scholar] [CrossRef]
- Hedrich, R.; Salvador-Recatalà, V.; Dreyer, I. Electrical wiring and long-distance plant communication. Trends Plant Sci. 2016, 21, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chao, M.; Wang, S.; Bu, J.; Tang, J.; Li, F.; Wang, Q.; Zhang, B. Proteome quantification of cotton xylem sap suggests the mechanisms of potassium-defficiency-induced changes in plant resistance to environmental stresses. Sci. Rep. 2016, 6, 21060. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Anschütz, U.; Becker, D.; Shabala, S. Going beyond nutrition: Regulation of potassium homeostasis as a common denominator of plant adaptive response to environment. J. Plant Physiol. 2014, 171, 670–687. [Google Scholar] [CrossRef]
- Shahzad, Z.; Canut, M.; Tournaire-Roux, C.; Martinière, A.; Boursiac, Y.; Loudet, O.; Maurel, C. A potassium-dependent oxygen sensing pathway regulates plant root hydraulics. Cell 2016, 167, 87–98. [Google Scholar] [CrossRef]
- Demidchik, V.; Straltsova, V.; Medvedev, S.S.; Pozhvanov, G.A.; Sokolik, A.; Yurin, V. Stress-induced electrolyte leakage: The role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp. Bot. 2014, 65, 1259–1270. [Google Scholar] [CrossRef]
- Wang, Y.; Wu, W.H. Genetic approaches for improvement of the crop potassium acquisition and utilization efficiency. Cur. Opin. Plant Biol. 2015, 25, 46–52. [Google Scholar] [CrossRef]
- Ashley, M.K.; Grabov, M.G.A. Plant responses to potassium deficiencies: A role for potassium transport proteins. J. Exp. Bot. 2005, 57, 425–436. [Google Scholar] [CrossRef]
- Becker, D.; Hoth, S.; Ache, P.; Wenkel, S.; Roelfsema, M.R.; Meyerhoff, O.; Harting, W.; Hedrich, R. Regulation of the ABA-sensitive Arabidopsis potassium channel gene GORK in response to water stress. FEBS Lett. 2003, 554, 119–126. [Google Scholar] [CrossRef]
- Qi, J.G.; Sun, S.M.; Yang, L.; Li, M.J.; Ma, F.W.; Zou, Y.J. Potassium uptake and transport in apple roots under drought stress. Horticul. Plant J. 2019, 5, 10–16. [Google Scholar] [CrossRef]
- Bortner, C.D.; Hughes, F.M., Jr.; Cidlowski, J.A. A primary role for K+ and Na+ efflux in the activation of apoptosis. J. Biol. Chem. 1997, 272, 32436–32442. [Google Scholar] [CrossRef] [PubMed]
- Demidchik, V.; Cuin, T.A.; Svistunenko, D.; Smith, S.J.; Miller, A.J.; Shabala, S.; Slkolik, A.; Yurin, V. Root K+-efflux conductance activated by hydroxyl radicals: Single-channel properties, genetic basis and involvement in stress-induced cell death. J. Cell Sci. 2010, 1233, 1468–1479. [Google Scholar] [CrossRef] [PubMed]
- Pilot, G.; Gaymard, F.; Mouline, K.; Cherel, I.; Sentenac, H. Regulated expression of Arabidopsis shaker K+ channel genes involved in K+ uptake and distribution in the plant. Plant Mol. Biol. 2003, 51, 773–787. [Google Scholar] [CrossRef]
- Sharp, R.E.; Hsiao, T.C.; Silk, W.K. Growth of the maize primary root at low water potentials: II. Role of growth and deposition of hexose and potassium in osmotic adjustment. Plant Physiol. 1990, 93, 1337–1346. [Google Scholar] [CrossRef] [PubMed]
- Zain, N.A.M.; Ismail, M.R.; Puteh, A.; Mahmood, M.; Isla, M.R. Drought tolerance and ion accumulation of rice following application of additional potassium fertilizer. Commun. Soil Sci. Plant Anal. 2014, 45, 2502–2514. [Google Scholar] [CrossRef]
- Teixeira, J.; Pereira, S. High salinity and drought act on an organ-dependent manner on potato glutamine synthetase expression and accumulation. Environ. Exp. Bot. 2007, 60, 121–126. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Sreenivasulu, N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue? Plant Cell Environ. 2014, 37, 300. [Google Scholar] [CrossRef]
- Prasad, D.; Singh, R.; Singh, A. Management of sheath blight of rice with integrated nutrients. Indian Phytol. 2010, 63, 11–15. [Google Scholar]
- Zhao, D.; Oosterhuis, D.M.; Bednarz, C.W. Influence of potassium deficiency on photosynthesis, chlorophyll content, and chloroplast ultrastructure of cotton plants. Photosynthetica 2001, 39, 103–109. [Google Scholar] [CrossRef]
- Zhu, X.C.; Song, F.B.; Liu, S.Q.; Liu, F.L.; Li, X.N. Arbuscular mycorrhiza enhances nutrient accumulation in wheat exposure to elevated CO2 and soil salinity. J. Plant Nutr. Soil Sci. 2018, 181, 836–846. [Google Scholar] [CrossRef]
- Laus, M.N.; Soccio, M.; Trono, D.; Liberatore, M.T.; Pastore, D. Activation of the plant mitochondrial potassium channel by free fatty acids and acyl-CoA esters: A possible defence mechanism in the response to hyperosmotic stress. J. Exp. Bot. 2011, 62, 141–154. [Google Scholar] [CrossRef]
- Trono, D.; Laus, M.N.; Soccio, M.; Alfaro, M.; Pastore, D. Modulation of Potassium cannel activity in the balance of ROS and ATP production by durum wheat mitochondria-an amazing defense tool agaisnt hyperosmotic stress. Front. Plant Sci. 2015, 6, 1072. [Google Scholar] [CrossRef]
- Amjad, M.; Akhtar, J.; Anwar-ul-Haq, M.; Riaz, M.A.; Saqib, Z.A.; Murtaza, B.; Naeem, M.A. Effectiveness of potassium in mitigating the salt-induced oxidative stress in contrasting tomato genotypes. J. Plant Nutr. 2016, 39, 1926–1935. [Google Scholar] [CrossRef]
- Liang, T.B.; Wang, Z.L.; Wang, R.J.; Liu, L.L.; Shi, C.Y. Effects of potassium humate on ginger root growth and its active oxygen metabolism. Ying Yong Sheng Tai Xue Bao 2007, 18, 813–817. [Google Scholar]
- García-Martí, M.; Piñero, M.C.; García-Sanchez, F.; Mestre, T.C.; López-Delacalle, M.; Martínez, V.; Rivero, R.M. Amelioration of the oxidatice stress generated by simple or combined abiotic stress through the K+ and Ca2+ supplementation in tomato plants. Antioxidants 2019, 8, 81. [Google Scholar] [CrossRef] [PubMed]
- Shirazi, M.U.; Asif, S.M.; Khanzada, M.; Khan, M.A.; Ali, M.; Mumtaz, S.; Yousufzai, M.N.; Saif, M.S. Growth and ion accumulation in some wheat genotypes under NaCl stress. Pak. J. Biol. Sci. 2001, 4, 388–391. [Google Scholar]
- Zheng, Y.; Aijun, J.; Tangyuan, N.; Jialin, X.; Zengjia, L.; Gaoming, J. Potassium nitrate application alleviates sodium chloride stress in winter wheat cultivars differing in salt tolerance. J. Plant Physiol. 2008, 165, 1455–1465. [Google Scholar] [CrossRef] [PubMed]
- Bar-Tal, A.S.; Sparks, D.L.F. Potassium-salinity interaction in irrigated corn. Irrig. Sci. 2004, 12, 27–35. [Google Scholar] [CrossRef]
- Cha-um, S.; Siringam, K.; Juntawong, N.; Kirdmanee, C. Water relations, pigment stabilization, photosynthetic abilities and growth improvement in salt stressed rice plants treated with exogenous potassium nitrate application. Int. J. Plant Prod. 2010, 4, 187–198. [Google Scholar]
- Pyo, Y.J.; Gierth, M.; Schroeder, J.I.; Cho, M.H. High-affinity K+ transport in Arabidopsis: AtHAK5 and AKT1 are vital for seedling establishment and post germination growth under low-potassium conditions. Plant Physiol. 2010, 153, 863–875. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Transcriptional regulatory networks in cellular responses and tolerance to dehydration and cold stresses. An. Rev. Plant Biol. 2006, 57, 781–803. [Google Scholar] [CrossRef] [PubMed]
- Osabake, Y.; Arinaga, N.; Umezawa, T.; Katsura, S.; Nagamachi, K.; Tanaka, H.; Ohiraki, H.; Yamada, K.; Seo, S.U.; Abo, M.; et al. Osmotic stress responses and plant growth controlled by potassium transporters in Arabidopsis. Plant Cell 2013, 25, 609–624. [Google Scholar]
- Kim, T.H.; Böhmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard cell signal transduction network: Advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu. Rev. Plant Biol. 2010, 61, 561–591. [Google Scholar] [CrossRef]
- Ueno, K.; Kinoshita, T.; Inoue, S.; Emi, T.; Shimazaki, K. Bio chemical characterization of plasma membrane HC-ATPase activation in guard cell protoplasts of Arabidopsis thaliana in response to blue light. Plant Cell Physiol. 2005, 46, 955–963. [Google Scholar] [CrossRef]
- Pilot, G.; Lacombe, B.; Gaymard, F.; Cherel, I.; Boucherez, J.; Thibaud, J.B.; Sentenac, H. Guard cell inward KC channel activity in Arabidopsis involves expression of the twin channel subunits KAT1 and KAT2. J. Biol. Chem. 2001, 276, 3215–3221. [Google Scholar] [CrossRef] [PubMed]
- Szyroki, A.; Ivashikina, N.; Dietrich, P.; Roelfsema, M.R.; Ache, P.; Reintanz, B.; Deeken, R.; Godde, M.; Felle, H.; Steinmeyer, R.; et al. KAT1 is not essential for stomatal opening. Proc. Natl. Acad. Sci. USA 2001, 98, 2917–2921. [Google Scholar] [CrossRef] [PubMed]
- Hedrich, R. Ion channels in plants. Physiol. Rev. 2012, 92, 1777–1811. [Google Scholar] [CrossRef] [PubMed]
- Talbott, L.D.; Zeiger, E. The role of sucrose in guard cell osmoregulation. J. Exp. Bot. 1998, 49, 329–337. [Google Scholar] [CrossRef]
- Geiger, D.; Scherzer, S.; Mumm, P.; Stange, A.; Marten, I.; Bauer, H.; Ache, P.; Matschi, S.; Liese, A.; Al-Rasheid, K.A.; et al. Activity of guard cell anion channel SLAC1 is controlled by drought-stress signaling kinase-phosphatase pair. Proc. Natl. Acad. Sci. USA 2011, 106, 21425–21430. [Google Scholar] [CrossRef]
- Geiger, D.; Scherzer, S.; Mumm, P.; Marten, I.A.; Ache, P.; Matschi, S.; Liese, A.; Wellmann, C.; Al-Rasheid, K.A.; Grill, E.; et al. Guard cell anion channel SLAC1 is regulated by CDPK protein kinases with distinct Ca2+ affinities. Proc. Natl. Acad. Sci. USA 2010, 107, 8023–8028. [Google Scholar] [CrossRef]
- Brandt, B.; Brodsky, D.E.; Xue, S.; Negi, J.; Iba, K.; Kangasiärvi, J.; Ghassemian, M.; Stephan, A.B.; Hu, H.; Schroeder, J.I. Reconstitution of abscisic acid activation of SLAC1 anion channel by CPK6 and OST1 kinases and branched ABI1 PP2C phosphatase action. Proc. Natl. Acad. Sci. USA 2012, 109, 10593–10598. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Muroyama, D.; Munemasa, S.; Nakamura, Y.; Mori, I.C.; Murata, Y. Calcium-dependent protein kinase CPK6 positively functions in induction by yeast elicitor of stomatal closure and inhibition by yeast elicitor of light induced opening in Arabidopsis. Plant Physiol. 2013, 163, 591–599. [Google Scholar] [CrossRef]
- Endo, A.; Sawada, Y.; Takahashi, H.; Okamoto, M.; Ikegami, K.; Koi-wai, H. Drought induction of Arabidopsis 9-cis-epoxycarotenoid dioxygenase occurs in vascular parenchyma cells. Plant Physiol. 2008, 147, 1984–1993. [Google Scholar] [CrossRef]
- Melhorn, V.; Matsumi, K.; Koiwai, H.; Ikegami, K.; Okamoto, M.; Nambara, E. Transient expres-s ion of AtNCED3 andAAO3 genes in the guard cells causes stomatal closur in Viciafaba. J. Plant Res. 2008, 121, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Scholz-Starke, J.; De Angelis, A.; Kovermann, P.; Gambale, B.B.F.; Martinoia, E. Malate transport by the vacuolar AtALMT6 channel in guard cells is subject to multiple regulation. Plant J. 2011, 67, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Jeanguenin, L.; Lebaudy, A.; Xicluna, J.; Alcon, C.; Hosy, E.; Duby, G. Heteromerization of Arabidopsis Kv channela-subunits. Plant Signal. Behav. 2008, 3, 622–625. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, J.I.; Keller, B.U. Two types of anion channels currents in guard cells with distinct voltage regulation. Proc. Natl. Acad. Sci. USA 1992, 89, 5025–5029. [Google Scholar] [CrossRef]
- Willmer, C.; Fricker, M. Stomata, 2nd ed.; Chap-Man &Hall: London, UK, 1996. [Google Scholar]
- Munemasa, S.; Oda, K.; Watanabe-Sugimoto, M.; Nakamura, Y.; Shi-moishi, Y.; Murata, Y. The coronatine-insensitive 1 mutation reveals the hormonal signaling interaction between abscisic acid and methyl jasmonate in Arabidopsis guard cells. Specific impairment of ion channel activation and second messenger production. Plant Physiol. 2007, 143, 1398–1407. [Google Scholar] [CrossRef]
- Lebaduy, A.; Vavasseur, A.; Hosy, E.; Dreye, I.; Leonhardt, N.; Thibaud, J.B.; Véry, A.A.; Simonneau, T.; Sentenac, H. Plant adaptation to fluctuating environment and biomass production are strongly dependent on guard cell potassium channels. Proc. Natl. Acad. Sci. USA 2008, 105, 5271–5276. [Google Scholar] [CrossRef]
- Blatt, M.R.; Thiel, G. KC channels of stomatal guard cells:bimodal control of the KC inward-rectifier evoked by auxin. Plant J. 1994, 5, 55–68. [Google Scholar] [CrossRef]
- Tanaka, Y.; Sano, T.; Tamaoki, M.; Nakajima, N.; Kondo, N.; Hasezawa, S. Cytokin and auxin inhibit abscisic acid-induced stomatal closure by enhancing ethylene production in Arabidopsis. J. Exp. Bot. 2006, 57, 2259–2266. [Google Scholar] [CrossRef]
- Dietrich, P.; Sanders, D.; Hedrich, R. The role of ion channels in light-dependent stomatal opening. J. Exp. Bot. 2001, 52, 1959–1967. [Google Scholar] [CrossRef]
- Liu, K.; Fu, H.; Bei, Q.; Luan, S. Inward potassium channel in guard cells as a target for polyamine regulation of stomatal movements. Plant Physiol. 2000, 124, 1315–1325. [Google Scholar] [CrossRef]
- Fazeli, F.; Ghorbanli, M.; Niknam, V. Effect of drought on biomass, protein content, lipid peroxidation and antioxidant enzymes in two sesame cultivars. Biol. Plant. 2007, 51, 98–103. [Google Scholar] [CrossRef]
- Degenkolbe, T.; Do, P.T.; Zuther, E.; Repsilber, D.; Walther, D.; Hincha, D.K.; Köhl, K.I. Expression profiling of rice cultivars differing in their tolerance to long-term drought stress. Plant Mol. Biol. 2009, 69, 133–153. [Google Scholar] [CrossRef]
- Soleimanzadeh, H.; Habibi, D.; Ardakani, M.R.; Paknejad, F.; Rejali, F. Effect of potassium levels on antioxidant enzymes and malondialdehyde content under drought stress in sunflower (Helianthus annuus L.). Am. J. Agric. Biol. Sci. 2010, 5, 56–61. [Google Scholar] [CrossRef]
- Cakmak, I. The role of potassium in alleviating detrimental effects of abiotic stresses in plants. J. Plant Nutr. Soil Sci. 2005, 168, 521–530. [Google Scholar] [CrossRef]
- Tomar, N.S.; Agarwal, R.M. Influence of treatment of Jatropha curcas L. leachates and potassium on growth and phytochemical constituents of wheat (Triticum aestivum L.). Am. J. Plant Sci. 2013, 4, 1134–1150. [Google Scholar] [CrossRef]
- Jatav, K.S.; Agarwal, R.M.; Tomar, N.S.; Tyagi, S.R. Nitrogen metabolism, growth and yield responses of wheat (Triticum aestivum L.) to restricted water supply and varying potassium treatments. J. Indian Bot. Soc. 2014, 93, 177–189. [Google Scholar]
- Hernandez, M.; Fernandez-Garcia, N.; Garcia-Garma, J.; Rubio-Asensio, J.S.; Rubio, F.; Olmos, E. Potassium starvation induces oxidative stress in Solanum lycopersicum L. roots. J. Plant Physiol. 2012, 169, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Agarwal, R.M. Potassium up-regulates antioxidant metabolism and alleviates growth inhibitor under water and osmotic stress in wheat (Triticum aesticum L). Protoplasma 2017, 254, 1471–1486. [Google Scholar] [CrossRef]
- Waraich, E.A.; Ahmad, R.; Halim, A.; Aziz, T. Alleviation of temperature stress by nutrient management in crop plants: A review. J. Soil Sci. Plant Nutr. 2012, 12, 221–244. [Google Scholar] [CrossRef]
- Egilla, J.N.; Davies, J.F.T.; Boutton, T.W. Drought stress influences leaf water content, photosynthesis, and water-use efficiency of Hibiscus rosa-sinensis at three potassium concentrations. Photosynthetica 2005, 43, 135–140. [Google Scholar] [CrossRef]
- Raza, M.A.S.; Saleem, M.F.; Shah, G.M.; Khan, I.H.; Raza, A. Exogenous application of glycinebetaine and potassium for improving water relations and grain yield of wheat under drought. J. Soil Sci. Plant Nutr. 2014, 14, 348–364. [Google Scholar] [CrossRef]
- Kanai, S.; Moghaieb, R.E.; El-Shemy, H.A.; Panigrahi, R.; Mohapatra, P.K.; Ito, J.; Nguyen, N.T.; Saneoka, H.; Fujita, K. Potassium deficiency affects water status and photosynthetic rate of the vegetative sink in green house tomato prior to its effects on source activity. Plant Sci. 2011, 180, 368–374. [Google Scholar] [CrossRef]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N.; Bonnemain, J.L.; Laloi, M.; Coutos-Thévenot, P.; Maurouss, L.; et al. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef]
- Maathuis, F.J.M.; Amtmann, A. K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, X.; Giraldo, J.P.; Shabala, S. It is nota ll about sodium: Revealing tissue specificity and signalling roles of potassium in plant responses to salt stress. Plant Soil 2018, 431, 1–17. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J.M. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2020, 10, 80. [Google Scholar] [CrossRef] [PubMed]
- Marten, I.; Hoth, S.; Deeken, R.; Ache, P.; Ketchum, K.A.; Hoshi, T.; Hedrich, R. AKT3, a phloem-localized K1 channel, is blocked by protons. Proc. Natl. Acad. Sci. USA 1999, 96, 7581–7586. [Google Scholar] [CrossRef]
- Maathuis, F.J.M. The role of monovalent cation transporters in plant responses to salinity. J. Exp. Bot. 2006, 57, 1137–1147. [Google Scholar] [CrossRef]
- Checchetto, V.; Teardo, E.; Carratto, L.; Leanza, L.; Szabo, I. Physiology of intracellular potassium channels: A unifying role as mediators of counterion fluxes? Biochim. Biophys. Acta Biol. 2016, 1857, 1258–1266. [Google Scholar] [CrossRef]
- Latorre, R.; Muñoz, F.; Gonzalez, C.; Cosmelli, D. Structure and function of potassium channels in plants: Some inferences about the molecular origin of inward rectification in KAT1 channels (Review). Mol. Membr. Biol. 2003, 20, 19–25. [Google Scholar] [CrossRef]
- Reiser, V.; Raitt, D.C.; Saito, H. Yeast osmosensor Sln1 and plant cytokinn receptor Cre1 respond to changes in turgor pressure. J. Cell Biol. 2003, 161, 1035–1040. [Google Scholar] [CrossRef]
- Wang, H.Y.; Zhou, J.M.; Du, C.W.; Chen, X.Q. Potassium fractions in soils as affected by monocalcium phosphate, ammonium sulfate and potassium chloride application. Pedosphere 2010, 20, 368–377. [Google Scholar] [CrossRef]
- Britzke, D.; da Silva, L.S.; Moterle, D.F.; dos Santos Rheinheimer, D.; Bortoluzzi, E.C. A study of potassium dynamics and mineralogy in soils from subtropical Brazilian lowlands. J. Soil Sed. 2012, 12, 185–197. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought changes phosphorus and potassium accumulation patterns in an evergreen Mediterranean forest. Funct. Ecol. 2007, 21, 191–201. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Prieto, P.; Estiarte, M. Drought and warming induced changes in P and K concentration and accumulation in plant biomass and soil in a Mediterranean shrubland. Plant Soil 2008, 306, 261–271. [Google Scholar] [CrossRef]
- Soil Survey Staff. Soil Taxonomy. A Basic System of Soil Classification for Making and Interpreting soil Surveys, 2nd ed.; US Department of Agriculture Handbook 436; Natural Resources Conservation Service, United States Department of Agriculture: Buckeye, AZ, USA, 1999; Volume 1, pp. 1–871.
- Jobbágy, E.G.; Jackson, R.B. The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 2001, 53, 51–77. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Hydraulic redistribution by plants and nutrient stoichiometry. Shifts under global change. Ecohydrology 2014, 7, 1–20. [Google Scholar] [CrossRef]
- Chen, G.; Liu, C.L.; Gao, Z.Y.; Zhang, Y.; Jiang, H.Z.; Zhu, L.; Ren, D.Y.; Yu, L.; Xu, G.H.; Qian, Q. OsHAK1, a high-affinity potassium transporter, positively regulates responses to drought stress rice. Front. Plant Sci. 2017, 8, 1885. [Google Scholar] [CrossRef]
- Han, M.; Wu, W.; Wu, W.H.; Wang, Y. Potassium transporter KUP7 is involved in K+ acquisition and translocation in Arabidopsis root under K+-limited conditions. Mol. Plant 2016, 9, 437–446. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability; Part B: Regional aspects. Contribution of working group II to the fifth assessment report of the intergovernmental panel on climate change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Mach, K.J., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014; p. 688. [Google Scholar]
- Benlloch-Gonzalez, M.; Sanchez-Lucas, R.; Benlloch, M. Effects of olive root warming on potassium transport and plant growth. J. Plant Physiol. 2017, 218, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Ventura, M.; Scandellari, F.; Bonora, E.; Tagliavini, M. Nutrient release during decomposition of leaf litter in a peach (Prunus persica L.) orchard. Nutrit. Cycl. Agroecosys. 2010, 87, 115–125. [Google Scholar] [CrossRef]
- Jonczak, J. Decomposition of beech leaves in 120-years old stand on the area of Middle Pomerania. Sylvan 2014, 158, 621–629. [Google Scholar]
- Vinh, T.V.; Allenbach, M.; Linh, K.T.V.; Marchand, C. Changes in leaf litter quality during its decomposition in a tropic planted mangrove forest (Can Gio, Vietnam). Front. Environ. Sci. 2020, 8, 10. [Google Scholar] [CrossRef]
- Ranibar, F.; Jalali, M. Calcium, magnesium, sodium, and potassium release during decomposition of some organic residues. Com. Soil Sci. Plant Anal. 2012, 43, 645–659. [Google Scholar] [CrossRef]
- Palviainen, M.; Finer, L.; Kurka, A.M.; Mannerkoski, H.; Piirainen, S.; Starr, M. Release of potassium, calcium, iron and aluminium from Norway spruce, scots pine and silver birch logging residues. Plant Soil 2004, 259, 123–136. [Google Scholar] [CrossRef]
- Moro, H.; Kunito, T.; Saito, T.; Yaguchi, N.; Sato, T. Soil microorganisms are less susceptible tan crop plants to potassium deficiency. Arch. Agon. Soil Sci. 2014, 60, 1807–1813. [Google Scholar] [CrossRef]
- Etesami, H.; Emami, S.; Alikhani, H.A. Potassium solubilizing bacteria (KSB): Mechanisms, promotion of plant growth, and future prospects–a review. J. Soil Sci. Plant Nutrit. 2017, 17, 897–911. [Google Scholar] [CrossRef]
- Khanghahi, M.Y.; Pirdashti, H.; Rahimian, H.; Nematzadeh, G.H.; Sapanlou, M.G.; Salvatori, E.; Crecchio, C. Leaf photosynthetic characteristics and photosystem II photochemistry of rice (Oryza sativa L.) under potassium-solubilizing bacteria inoculation. Photosynthetica 2019, 57, 500–511. [Google Scholar] [CrossRef]
- Burghelea, C.; Zaharescu, D.G.; Dontsova, K.; Maier, R.; Huxman, T.; Chorover, J. Mineral nutritient mobilization by plants from rock: Influence of rock type and arbuscular mycorrhiza. Biogeochemistry 2015, 124, 187–203. [Google Scholar] [CrossRef]
- Ajithkumar, I.P.; Panneerselvam, R. ROS scavenging system, osmotic maintenance pigment and growth status of Panicum sumatrense Roth. Under drought stress. Cell Biochem. Biophys. 2013, 68, 587–595. [Google Scholar] [CrossRef]
- Wang, F.Y.; Sun, Y.H.; Shi, Z.Y. Arbuscular mycorrhiza enhances biomass production and salt tolerance of sweet sorghum. Microorganisms 2019, 7, 289. [Google Scholar] [CrossRef]
- Wang, H.Y.; Shen, Q.H.; Zhou, J.M.; Wang, J.; Du, C.W.; Chen, X.Q. Plants use alternative strategies to utilize nonexchangeable potassium in minerals. Plant Soil 2011, 343, 209–220. [Google Scholar] [CrossRef]
- Yang, Z.X.; Yu, S.Z.; Lin, Y.C.; Zhang, W.J.; Wang, Y.; Wang, R.G.; Xu, S.X.; Yang, T.Z.; Xue, G. Activation of potassium released from soil by root-secreted organic acids in different varieties of tabacco (Nicotiniana tabacum). Funct. Plant Biol. 2020, 47, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.F.; Yang, Z.X.; Yu, S.Z.; Chen, H.L. Organic acids exuded from roots increase the available potassium content in the rhizosphere soil: A rizobag experiment in Nicotiana tabacum. Hortscience 2019, 54, 23–27. [Google Scholar] [CrossRef]
- Gahoonia, T.S.; Nielsen, N.E. Root activity and acquisition of soil phosphorus and potassium. Edited by Box, J.E. Root Demographics and Their Efficiencies in Sustainable Agriculture, Grassland and Forest Ecosystems. Develop. Plant Soil Sci. 1998, 82, 331–344. [Google Scholar]
- Yuan, J.; Huang, J.G.; Li, X.L.; Christie, P. Biological mobilization of potassium from clay minerals by ectomycorrhizal fungi and eucalypt seedling roots. Plant Soil 2004, 262, 351–361. [Google Scholar] [CrossRef]
- Yang, T.; Lu, X.; Wang, Y.; Xie, Y.; Ma, J.; Cheng, X.; Xia, E.; Wan, X.; Zhang, Z. HAK/KUP/KT family potassium transporter genes are involved in potassium deficiency and stress responses in tea plants (Camelia sinensis L.): Expression and functional analysis. BMC Gen. 2020, 21, 556. [Google Scholar] [CrossRef]
- Argiolas, A.; Puleo, G.L.; Sinibaldi, E.; Mazzolai, B. Osmolyte cooperation affects turgor dynamics in plants. Sci. Rep. 2016, 6, 30139. [Google Scholar] [CrossRef] [PubMed]
- Grefen, C.; Karnik, R.; Larson, E.; Lefoulon, C.; Wang, Y.Z.; Waghmare, S.; Zhang, B.; Hills, A.; Blatt, M.R. A vesicle-trafficking protein commanders kv channel sensors for voltage-dependent secretion. Nat. Plants 2015, 1, 15108. [Google Scholar] [CrossRef] [PubMed]
- Mengel, K. Principles of Plant Nutrition, 5th ed.; Kluwer Academic: Dordrecht, The Netherlands, 2001; pp. 481–509. [Google Scholar]
- Zhao, W.Q.; Dong, H.R.; Zahoor, R.W.; Zhou, Z.G.; Snider, J.L.; Chen, Y.L.; Siddique, K.H.M.; Wang, Y.H. Ameliorative effects of potassium on drought-induced decreases in fiber length of cotton (Gossypium hirsutum L.) are associated with osmolyte dynamics during fiber development. Crop J. 2019, 7, 619–634. [Google Scholar] [CrossRef]
- Berg, W.K.; Cunningham, S.M.; Brouder, S.M.; Joern, B.C.; Johnson, K.D.; Volence, J.J. Influence of phosphorus and potassium on alfalfa yield, taproot C and N pools, and transcript levels of key genes after defoliation. Crop Sci. 2009, 49, 974–982. [Google Scholar] [CrossRef]
- Wright, S.J.; Yavitt, J.B.; Wurzburger, N.; Turner, B.L.; Tanner, E.V.J.; Sayer, E.J.; Santiago, L.S.; Kasparl, M.; Hedin, L.O.; Harms, K.E.; et al. Potassium, phosphorus, or nitrogen limit root allocation tree growth, or litter production in a lowland tropical forest. Ecology 2011, 92, 1616–1625. [Google Scholar] [CrossRef]
- Bond, W.J. Do nutrient-poor soils inhibit development of forest? A nutrient stock analysis. Plant Soil 2010, 334, 47–60. [Google Scholar] [CrossRef]
- Jin, S.H.; Huang, J.Q.; Li, X.Q.; Zheng, B.S.; Wu, J.; Wang, Z.J.; Liu, G.H.; Chen, M. Effects of potassium supply on limitations of photosynthesis by mesophyll diffusion conductance in Carya cathayensis. Tree Physiol. 2011, 31, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.; Stewart, G.R. Effect of potassium levels on the stomatal behavior of the hemi-parasite Striga hermonthica. Plant Physiol. 1990, 94, 1472–1476. [Google Scholar] [CrossRef] [PubMed]
- Carlson, C.A.; Fox, T.R.; Allen, H.L.; Albaugh, T.J.; Rubilar, R.A.; Stape, J. Growth response of Loblolly Pine in the southeast United States to Midrotation applications of nitrogen, phosphorus, potassium, and micronutrients. For. Sci. 2014, 60, 157–169. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11106. [Google Scholar] [CrossRef] [PubMed]
- Kerkhoff, A.J.; Enquist, B.J.; Elser, J.J.; Fagan, W.F. Plant allometry, stoichiometry and the temperature-dependence of primary productivity. Glob. Ecol. Biogeog. 2005, 14, 585–598. [Google Scholar] [CrossRef]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. Factors affecting nutrient concentration and stoichiometry of forest trees in Catalonia (NE Spain). For. Ecol. Manag. 2011, 262, 2024–2034. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Coll, M.; Vayreda, J.; Rivas-Ubach, A. Stoichiometry of potassium is largely determined by water availability and growth in Catalonian forests. Funct. Ecol. 2012, 26, 1077–1089. [Google Scholar] [CrossRef]
- Tomlinson, K.W.; Pooter, L.; Sterck, F.J.; Borghetti, F.; Ward, D.; de Bie, S.; van Langevelde, F. Leaf adaptations of evergreen and deciduous trees of semi-arid and humid savannas on three continents. J. Ecol. 2013, 101, 430–440. [Google Scholar] [CrossRef]
- Milla, R.; Castro-Díaz, P.; Maestro-Martínez, M.; Montserrat-Martí, G. Relationships between phenology and the remobilization of nitrogen, phosphorus and potassium in branches of eight Mediterranean evergreens. New Phytol. 2005, 168, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Rivas-Ubach, A.; Sardans, J.; Pérez-Trujillo, M.; Estiarte, M.; Peñuelas, J. Strong relationships between elemental stoichiometry and metabolome in plants. Proc. Natl. Acad. Sci. USA 2012, 109, 4181–4186. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climaste Change: Impacts, Adaptation and Vulnerability. In Contribution of Working Group II to the Fourth Assessment Report of the IPCC; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Benlloch-Gonzalez, M.; Quintero, J.M.; Suárez, M.P.; Sánchez-Lucas, R.; Fernández-Escobar, R.; Benlloch, M. Effect of moderate high temperatures on the vegetative growth and potassium allocation in olive plants. J. Plant Physiol. 2016, 207, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Ge, T.D.; Sun, N.B.; Bai, L.P.; Tong, C.L.; Sui, F.G. Effects of drought stress on phosphorus and potassium uptake dynamics in summer maize (Zea mays) through the growth cycle. Acta Physiol. Plant. 2012, 34, 2179–2186. [Google Scholar] [CrossRef]
- Watson, R.; Pritchard, J.; Malone, M. Direct measurement of sodium and potassium in the transpirating stream of salt-excluding and non-excluding varieties of wheat. J. Exp. Bot. 2001, 52, 1873–1881. [Google Scholar] [CrossRef][Green Version]
- Oliveira, R.H.; Rosolem, C.A.; Trigueiro, R.M. Importance of mass flow and diffusion on the potassium supply to cotton plants as affected by soil water and potassium. Rev. Brasil. Ciencia Solo 2004, 28, 439–445. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Plant–soil interactions in Mediterranean forest and shrublands: Impacts of climatic change. Plant Soil 2013, 365, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Ryel, R.J. Hydrailic redistribution. Progr. Bot. 2004, 65, 413–435. [Google Scholar]
- Smith, D.M.; Jackson, N.A.; Roberts, J.M.; Ong, C.K. Reverse flow of sap in tree roots and downward siphoning of water by Grevillea robusta. Funct. Ecol. 1999, 13, 256–264. [Google Scholar] [CrossRef]
- Florez-Florez, C.P.; Leon-Pelaez, J.D.; Osorio-Vega, N.W.; Restrepo-Llano, M.F. Nutrient dynamics in forest plantations of Azadirachta indica (Meliaceae) established for restoration of degraded lands in Colombia. Rev. Biol. Trop. 2013, 61, 515–529. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schreeg, L.A.; Mack, M.C.; Turner, B.L. Nutrient specific patterns of leaf litter across 41 lowland tropical woody species. Ecology 2013, 94, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Watmough, S.A.; Aherne, J.; Alewell, C.; Arp, P.; Bailey, S.; Clair, T.; Dillon, P.; Duchesne, L.; Eimers, C.; Fernandez, I.; et al. Sulphate, nitrogen and base cation budgets at 21 forested catchments in Canada, the United States and Europe. Environ. Monit. Assess. 2005, 109, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Anguelov, G.; Anguelova, I.; Bailey, N.O. Land-use impact on soil solution constituents from an Ultisol of north Florida. Nutri. Cycl. Agroecosyst. 2011, 90, 171–187. [Google Scholar] [CrossRef]
- Dunlop, J.; Glass, A.D.M.; Tomkins, B.D. Regulation of K+ uptake by ryegrass and white clover roots in relation to their competition for potassium. New Phytol. 1979, 83, 365–370. [Google Scholar] [CrossRef]
- Tilman, E.A.; Tilman, D.; Crawley, M.J.; Johnston, A.E. Biological weed control via nutrient competition: Potassium limitation of dandelions. Ecol. Monogr. 1999, 9, 103–111. [Google Scholar] [CrossRef]
- Neva, G.A.; Newbery, D.M.; Chuyong, G.B. Limitation of seedling growth by potassium and magnesium supply for two ectomycorrhizal tree species of a central African rain forest and its implication for their recruitment. Ecol. Evol. 2016, 6, 125–142. [Google Scholar]
- Khan, S.A.; Mulbaney, R.L.; Ellsworth, T.R. The potassium paradox: Implications for soil fertility, crop production an human health. Renew. Agric. Food Syst. 2014, 29, 3–27. [Google Scholar] [CrossRef]
- Gélinas, Y.; Lucotte, M.; Schmit, J.P. History of the atmospheric deposition of major and trace elements in the industrialized St. Lawrence Valley, Quebec, Canada. Atmos. Environ. 2000, 34, 1797–1810. [Google Scholar] [CrossRef]
- Ruoho-Airola, T.; Salminen, K. Trends in the base cation deposition in Finland. Air Pollution XI. Adv. Air Pollut. Ser. 2003, 13, 273–282. [Google Scholar]
- Allen, A.G.; Cardoso, A.A.; Wiate, A.G.; Machado, C.M.D.; Paterlini, W.C.; Baker, J. Influence of intensive agriculture on dry deposition of aerosol nutrients. J. Braz. Chem. Soc. 2010, 21, 87–97. [Google Scholar] [CrossRef]
- Ferm, M.; Hultberg, H. Dry deposition and internal circulation of nitrogen, sulphur and base cations to a coniferous forest. Atmos. Dep. 1999, 33, 4421–4430. [Google Scholar] [CrossRef]
- Golobocanin, D.; Zujic, A.; Milenkovic, A.; Miljevic, N. Precipitation composition and wet deposition temporal pattern in Central Serbia for the period from 1998 to 2004. Environ. Monit. Assess. 2009, 142, 185–198. [Google Scholar] [CrossRef]
- Poor, N.; Pollman, C.; Tate, P.; Begum, M.; Evans, M.; Campbell, S. Nature and magnitude of atmospheric fluxes of total inorganic nitrogen and other inorganic species to the Tampa Bay watershed, FL, USA. Water Air Soil Pollut. 2006, 170, 267–283. [Google Scholar] [CrossRef]
- Prathibha, P.; Kothai, P.; Saradhi, I.V.; Pandit, G.G.; Puranik, V.D. Chemical characterization of precipitation at a coastal site in Trombay, Mumbai, India. Environ. Monit. Assess. 2010, 168, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Friedland, A.J.; Miller, E.K. Major-element cycling in a high-elevation Adirondack forest: Patterns and changes, 1986–1996. Ecol. Appl. 1999, 9, 958–967. [Google Scholar]
- Watmough, S.A.; Dillon, P.J. Major element fluxes from a coniferous catchment in central Ontario, 1983–1999. Biogeochemistry 2004, 67, 369–398. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Sanford, R.L. Nutrient cycling in moist tropical forest. An. Rev. Ecol. Syst. 1986, 17, 585–594. [Google Scholar] [CrossRef]
- Muoghalu, J.I.; Oakhumen, A. Nutrient content of incident rainfall, throughfall and stemflow in a Nigerian secondary lowland rainforest. Appl. Veg. Sci. 2000, 3, 181–188. [Google Scholar] [CrossRef]
- Ahmad, Z.; Anjum, S.; Waraichj, E.A.; Ayub, M.A.; Ahmad, T.; Tariq, R.M.S.; Ahmad, R.; Iqbal, M.A. Growth, physiology, and biochemical activities of plant responses with foliar potassium application under drought stress–a review. J. Plant Nutr. 2018, 41, 1734–1743. [Google Scholar] [CrossRef]
- Farahani, S.; Heravan, E.M.; Rad, A.H.S.; Noormohammadi, G. Effect of potassium sulfate on quantitative and qualitative characteristics of canola cultivars upon late-season drought stress conditions. J. Plant Nutr. 2019, 42, 1543–1555. [Google Scholar] [CrossRef]
- Hussain, S.; Tahir, M.; Tanveer, A.; Ahmad, R. Mitigating the effects of drought stress in wheat through potassium application. Int. J. Bot. St. 2019, 4, 86–91. [Google Scholar]
- Ma, Q.F.; Bell, R.; Biddulph, B. Potassium application alleviates grain sterility and increases yield of wheat (Triticum aesticum) in frost-prone Mediterranean-type climate. Plant Soil 2019, 434, 203–2016. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Nahar, K.; Hossain, M.S.; Al Mahmud, J.; Hossen, M.S.; Masud, A.A.C.; Moumita, F.M. Potassium: A vital regulator of plant responses and tolerance to abiotic stresses. Agron. Basel 2018, 8, 31. [Google Scholar] [CrossRef]
- Ahmad, W.; Ayyub, C.M.; Shehzad, M.A.; Ziaf, K.; Ijaz, M.; Sher, A.; Abbas, T.; Shafi, J. Supplemental potassium mediates antioxidant metabolism, physiological processes, and osmoregulation to confer salt stress tolerant in cabbage (Brasica oleracea L.). Horticul. Environ. Biotechnol. 2019. [Google Scholar] [CrossRef]
- Gul, M.; Wakeel, A.; Steffens, D.; Lindberg, S. Potassium-induced decrease in cytosolic Na+ alleviates deleterious effects of salt stress on wheat (Triticum aesticum L.). Plant Biol. 2019, 21, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Senbayran, M.; Peiter, E. Potassium in Agriculture–status and perspectives. J. Plant Physiol. 2014, 171, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; Wu, X.; Härdter, R. The interaction between soil water regime and potassium availability on the growth of tea. Commun. Soil Sci. Plant Anal. 1997, 28, 89–98. [Google Scholar] [CrossRef]
- Römheld, V.; Kirkby, E.S. Research on potassium in agriculture: Needs and prospects. Plant Soil 2010, 335, 155–180. [Google Scholar] [CrossRef]
- Ryan, J.; Sommer, R. Soil fertility and crop nutrition research at an international center in the Mediterranean region: Achievements and future perspective. Arch. Agric. Soil Sci. 2012, 58, S41–S54. [Google Scholar] [CrossRef]
- Ryan, J.; Sommer, R.; Ibrikci, H. Fertilizer best management practices: A perspective from the dryland west Asia– North Africa region. J. Agric. Crop Sci. 2012, 198, 57–67. [Google Scholar] [CrossRef]
- Singh, K.; Bansai, S.K. Potassium indexing of crops grown on eight benchmark soil series of India. Com. Soil Sci. Plant Anal. 2009, 40, 1369–1379. [Google Scholar] [CrossRef]
- Sharifi, M.; Cheema, M.; McVicar, K.; LeBlanc, L.; Fillmore, S. Evaluation of liming properties and potassium bioavailability of three Atlantic Canada wood ash sources. Can. J. Plant Sci. 2013, 93, 1209–1216. [Google Scholar] [CrossRef]
- Mitchell, A.D.; Smethurst, P.J. Base cation availability and leaching after nitrogen fertilization of a eucalypt plantation. Aust. J. Soil Res. 2008, 46, 445–454. [Google Scholar] [CrossRef]
- Yavitt, J.B.; Harms, K.E.; Garcia, M.N.; Mirabello, M.J.; Wright, S.J. Soil fertility and fine root dynamics in response to 4 years of nutrient (N, P, K) fertilization in a lowland tropical moist forest Panama. Austral Ecol. 2011, 36, 433–445. [Google Scholar] [CrossRef]
- Glab, T.; Gondek, K. The influence of soil compaction and fertilization on physico-chemical properties of mollic fluvisol under red clover/grass mixture. Geoderma 2014, 226, 204–212. [Google Scholar] [CrossRef]
- Roberts, T.L. Global Potassium Reserves and Potassium Fertilizer Use. Presentation to Global Nutrient Cycling Symposium; International Plant Nutrition Institute: Peachtree Corners, GA, USA, 2008; Available online: http://www.ipni.net/ipniweb/portal.nsf/0/9c5cff1af71db2ce852574e8004ecc00/$FILE/Roberts%20-%20ASA%20Nutrient%20Cycling%20Symposium%20Potash.pdf (accessed on 5 December 2014).
- Cakmak, I. Potassium for better crop production and quality. Plant Soil 2010, 335, 1–2. [Google Scholar] [CrossRef]
- Dhillon, J.S.; Eickhoff, E.M.; Mullen, R.W.; Raun, W.R. World potassium use efficiency in Cereal crops. Agron. J. 2019, 111, 889–896. [Google Scholar] [CrossRef]
- Mikhailova, E.A.; Post, G.; Cope, M.; Post, C.; Schlautman, M.A.; Zhang, L. Quantifying and mapping atmospheric potassium deposition for soil ecosystem services assessment in the United States. Front. Environ. Sci. 2019, 7, 74. [Google Scholar] [CrossRef]
- Grimshaw, H.J.; Dolske, D.A. Rainfall concentrations and wet atmospheric deposition of phosphorus and other constituents in Florida, U.S.A. Water Air Soil Pollut. 2002, 137, 117–140. [Google Scholar] [CrossRef]
- Klumpp, A.; Domingos, M.; Klumpp, G. Foliar nutrient contents in tree species of the Atlantic rainforest as influenced by air pollution from the industrial complex of Cubatao, SE-Brazil. Water Air Soil Pollut. 2002, 133, 315–333. [Google Scholar] [CrossRef]
- Gorostiza, S.; Saurí, D. Naturalizing pollution: A critical social science view on the link between potash mining and salinization in the Llobregat river basin, northeast Spain. Phill. Trans. R. Soc. B 2018, 374, 20180006. [Google Scholar] [CrossRef]
- Urban, R.C.; Lima-Souza, M.; Caetano-Silva, L.; Queiroz, M.E.C.; Nogueira, R.F.P.; Allen, A.G.; Cardoso, A.A.; Held, G.; Campos, M.L.A.M. Use of levoglucosan, potassium, and water-soluble organic carbon to characterize the origins of biomass-burning aerosols. Atm. Environ. 2012, 61, 562–569. [Google Scholar] [CrossRef]
- Tung, P.G.A.; Yussok, M.K.; Majid, N.M.; Joo, G.J.; Huang, G.H. Effect of N and K fertilizers on nutrient leaching and groundwater quality under mature oil palm in Sabah during the Monsoon period. Am. J. Appl. Sci. 2009, 6, 1788–1799. [Google Scholar] [CrossRef]
- Mendes, W.C.; Júnior, J.A.; da Cunha, P.C.R.; da Silva, A.R.; Evangelista, A.W.P.; Casaroli, D. Potassium leaching in different Soils as a function of irrigation depths. Rev. Brasil. Engenharia Agríla Amb. 2016, 20, 972–977. [Google Scholar] [CrossRef]
- Cowan, A.E.; Jun, S.; Tooze, J.A.; Eicher-Miller, H.A.; Dodd, K.W.; Gahche, J.J.; Guenther, P.M.; Dwyer, J.T.; Potischman, N.; Bhadra, A.; et al. Total Usual Micronutrient Intakes Compared to the Dietary Reference Intakes among U.S. Adults by Food Security Status. Nutrients 2020, 12, 38. [Google Scholar] [CrossRef]
- Pettigrew, W.T. Potassium influences on yield and quality production for maize, wheat, soybean and cotton. Physiol. Plant 2008, 133, 670–681. [Google Scholar] [CrossRef]
- Valentinuzzi, F.; Maver, M.; Fontanari, S.; Mott, D.; Savini, G.; Tiziani, R.; Pii, Y.; Mimmo, T.; Cesco, S. Foliar application of potassium-based fertilizer improves strawberry fruit quality. VIII International Symposium on Mineral Nutrition of Fruit Crops. Ed. by: Mimmo, T., Pii, Y., Scandellari, F. Book Series. Acta Hortic. 2018, 1217, 379–384. [Google Scholar] [CrossRef]
- Holland, J.E.; Hayes, R.C.; Refshauge, G.; Poile, G.J.; Newell, M.T.; Conyers, M.K. Biomass, feed quality, mineral concentration and grain yield responses to potassium fertilizer of dual-purpose crops. N. Z. J. Agric. Res. 2019, 62, 476–494. [Google Scholar] [CrossRef]
- Milford, G.F.J.; Armstrong, M.J.; Jarvis, P.J.; Houghton, B.J.; Bellett-Travers, D.M.; Jones, J.; Leigh, R.A. Effect of potassium fertilizer on the yield, quality and potassium offtake of sugar beet crops grown on soils of different potassium status. J. Agric. Sci. 2000, 135, 1–10. [Google Scholar] [CrossRef]
- Wu, L.Q.; Cui, Z.L.; Chen, X.P.; Zhao, R.F.; Si, D.X.; Sun, Y.X.; Yue, S.C. High-Yield maize production in relation to potassium uptake requirements in China. Agron. J. 2014, 106, 1153–1158. [Google Scholar] [CrossRef]
- Preciado-Rangel, P.; Salas-Pérez, L.; Gallegos-Robles, M.A.; Ruiz-Espinoza, F.H.; Ayala-Garay, A.V.; Fortis-Hernández, M.; Murillo-Amador, B. Increasing doses of potassium increases yield and Quality of muskmelon fruits under greenhouse. Hortic. Bras. 2018, 36, 184–188. [Google Scholar] [CrossRef]
- Umar, S.; Bansal, S.K.; Imas, P.; Magen, H. Effect of foliar fertilization of potassium on yield, quality, and nutrient uptake of groundnut. J. Plant Nutr. 2008, 22, 1785–1795. [Google Scholar] [CrossRef]
- Yawson, D.O.; Adu, M.O.; Ason, B.; Armah, F.A.; Boateng, E.; Quansah, R. Ghanaians might be a risk of excess dietary intake of potassium based on food supply data. J. Nutrit. Metab. 2018, 1, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lanham-New, S.; Lambert, H. Potassium. Adv. Nutr. 2012, 3, 820–821. [Google Scholar] [CrossRef]
- Dorup, I. Magnesium and potassium deficiency. Its diagnosis, occurrence and treatment in diuretic therapy and its consequences for growth, protein synthesis and growth factors. Acta Physiol. Scad. 1994, 618, 1–55. [Google Scholar]
- Sharkey, J.R.; Nalty, C.; Johnson, C.M.; Dean, W.R. Children’s very low food security is associated with increased dietary intakes and energy, fat, and added sugar among Mexican-origin children (6-11 y) in Texas border Colonias. BMC Pediatr. 2012, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Park, G.A.; Kim, S.H.; Kim, S.J.; Yang, Y.J. Health and nutritional status of Korean adults according to age and household food security: Using the data from 2010–2012 Korea National Health and Nutrition Examination survey. J. Nutr. Health 2017, 50, 603–614. [Google Scholar] [CrossRef]
- Clegg, D.J.; Headley, S.A.; Germain, M.J. Impact of dietary potassium restrictions in CKD on clinical outcomes benefits of a plnat-based diet. Kidney Med. 2020, 2, 476–487. [Google Scholar] [CrossRef]
- Sun, H.B.; Weaver, C.M. Rise in potassium deficiency in the US population linked to agriculture practices and dietary potassium deficits. J. Agric. Food Chem. 2020, 68, 11121–11127. [Google Scholar] [CrossRef]
- He, F.J.; MacGregor, G.A. Beneficial effects of potassium on human health. Physiol. Plant. 2008, 133, 725–735. [Google Scholar] [CrossRef]
- Whelton, P.K.; He, J. Health effects of sodium and potassium in humans. Curr. Opin. Lipidol. 2014, 25, 75–79. [Google Scholar] [CrossRef]
- Bekele, M. Effects of different levels of potassium fertilization on yield, quality and storage life of onion (Allium cepa L.) at Jimma, Southwestern Ethiopia. J. Food Sci. Nutr. 2018, 1, 32–39. [Google Scholar] [CrossRef]
- Liu, Y.X.; Ma, J.C.; Ding, W.C.; He, W.T.; Lei, Q.L.; Gao, Q.; He, P. Temporal and spatial variation of potassium balance in agricultural land at national and regional levels in China. PLoS ONE 2017, 12, e0184156. [Google Scholar] [CrossRef]
- Panda, B.B.; Sharma, S.; Mohapatra, P.K.; Das, A. Application of excess nitrogen, phosphorus, and potassium fertilizers leads to lowering of grain iron content in high-yielding tropical rice. Com. Soil Sci Plant Anal. 2011, 43, 2590–2602. [Google Scholar] [CrossRef]
- Xu, X.; Du, X.; Wang, F.; Sha, J.; Chen, Q.; Tian, G.; Zhu, Z.; Ge, S.; Jiang, Y. Growth, accumulation and distribution of carbon, and nitrate metabolism in apple dwarf rootstock seedlings. Front. Plant Sci. 2020, 11, 904. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gao, M.; Li, S.; Alva, A.K.; Ashraf, M. Potassium fertilization mitigates the adverse effects of drought on selected Zea mays cultivars. Turk. J. Bot. 2014, 38, 713–723. [Google Scholar]
- Zheng, J.B.; Quan, X.Y.; He, X.Y.; Cai, S.G.; Ye, Z.L.; Chen, G.; Zhang, G.P. Root and leaf metabolite profiles analysis reveals the adaptive strategies to low potassium stress in barley. BMC Plant Biol. 2018, 18, 187. [Google Scholar]
- Mao, X.H.; Zheng, Y.M.; Xiao, K.Z.; Wei, Y.D.; Zhu, Y.S.; Cai, Q.H.; Chen, L.P.; Xie, H.A.; Zhang, J.F. OsPRX2 contributes to stomatal closure and improves potassium deficiency tolerance in rice. Biochem. Biophy. Res. Commun. 2018, 495, 461–467. [Google Scholar] [CrossRef]
- Cao, X.; Hu, L.; Chen, X.; Zhang, R.; Cheng, D.; Li, H.; Xu, Z.; Li, L.; Zhou, Y.; Liu, A.; et al. Genome-wide analysis and identification of the low potassium stress responsive gene SiMYB3 in foxtail millet (Setaria italica L.). BMC Genom. 2019, 20, 136. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sardans, J.; Peñuelas, J. Potassium Control of Plant Functions: Ecological and Agricultural Implications. Plants 2021, 10, 419. https://doi.org/10.3390/plants10020419
Sardans J, Peñuelas J. Potassium Control of Plant Functions: Ecological and Agricultural Implications. Plants. 2021; 10(2):419. https://doi.org/10.3390/plants10020419
Chicago/Turabian StyleSardans, Jordi, and Josep Peñuelas. 2021. "Potassium Control of Plant Functions: Ecological and Agricultural Implications" Plants 10, no. 2: 419. https://doi.org/10.3390/plants10020419
APA StyleSardans, J., & Peñuelas, J. (2021). Potassium Control of Plant Functions: Ecological and Agricultural Implications. Plants, 10(2), 419. https://doi.org/10.3390/plants10020419