Summary of Natural Products Ameliorate Concanavalin A-Induced Liver Injury: Structures, Sources, Pharmacological Effects, and Mechanisms of Action


 ,
, 
Abstract
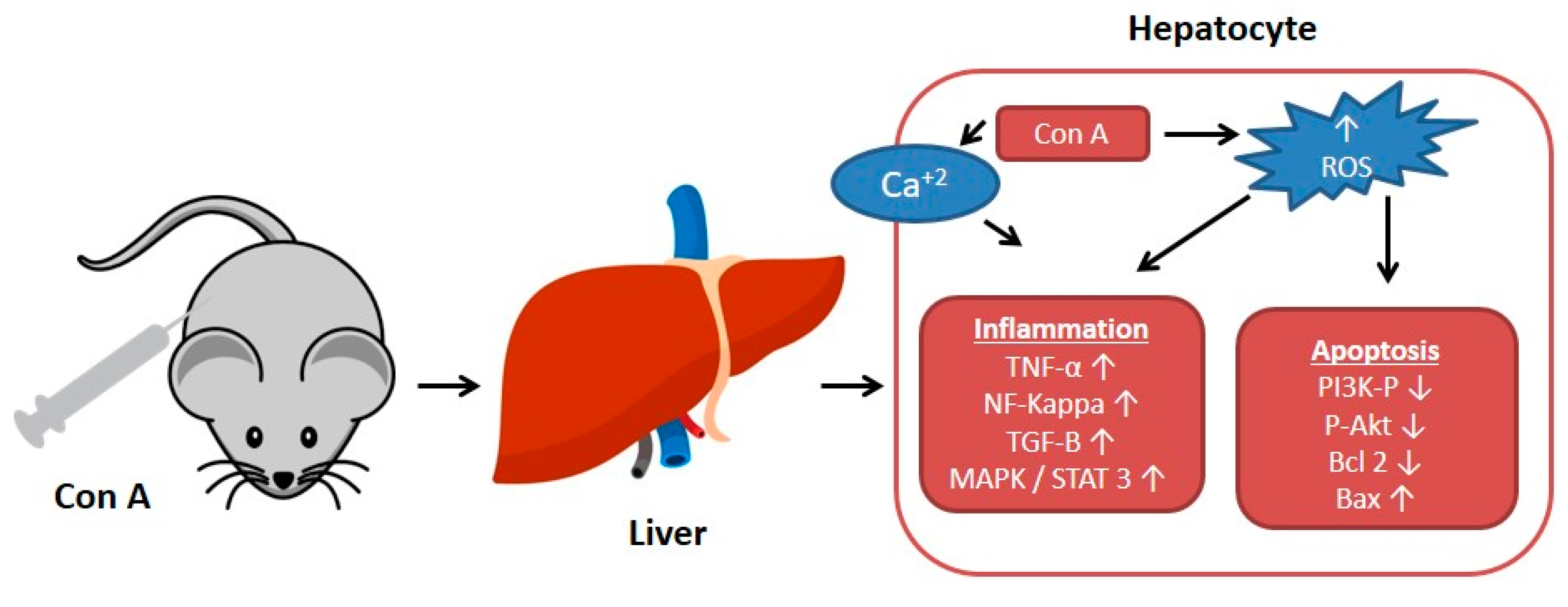
1. Introduction
2. Natural Compounds with Hepato-Protective Effect on Con A-Induced Injury
2.1. Alkaloids
2.2. Terpenes
2.3. Sterols
2.4. Phenolic Compounds
2.5. Coumarins and Coumarin Derivatives
2.6. Miscellaneous
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Gantner, F.; Leist, M.; Lohse, A.W.; Germann, P.G.; Tiegs, G. Concanavalin A-induced T-cell-mediated hepatic injury in mice: The role of tumor necrosis factor. Hepatology 1995, 21, 190–198. [Google Scholar] [PubMed]
- Miethke, T.; Wahl, C.; Heeg, K.; Echtenacher, B.; Krammer, P.H.; Wagner, H. T cell-mediated lethal shock triggered in mice by the superantigen Staphylococcal enterotoxin B: Critical role of tumor necrosis factor. J. Exp. Med. 1992, 175, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Tiegs, G.; Hentschel, J.; Wendel, A. A T cell-dependent experimental liver injury in mice inducible by concanavalin A. J. Clin. Investig. 1992, 90, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Bai, Y.; Gao, M.; Zhang, J.; Dong, G.; Yan, F.; Ma, Q.; Fu, X.; Zhang, Q.; Li, C.; et al. Hepatoprotective effect of capsaicin against concanavalin A-induced hepatic injury via inhibiting oxidative stress and inflammation. Am. J. Transl. Res. 2019, 11, 3029–3038. [Google Scholar]
- Zhang, Y.; Li, L.; Qi, C.; Hua, S.; Fei, X.; Gong, F.; Fang, M. Glycyrrhizin alleviates Con A-induced hepatitis by differentially regulating the production of IL-17 and IL-25. Biomed. Pharmacother. 2019, 110, 692–699. [Google Scholar] [CrossRef]
- Ballegeer, M.; Libert, C. Different cell types involved in mediating concanavalin A induced liver injury: A comprehensive overview. J. Gastroenterol. Hepatol. Res. 2016, 1. [Google Scholar] [CrossRef]
- Mikkelsen, R.B.; Schmidt-Ullrich, R. Concanavalin A induces the release of intracellular Ca2+ in intact rabbit thymocytes. J. Biol. Chem. 1980, 255, 5177–5183. [Google Scholar] [CrossRef]
- Liu, J.; Mao, Y. Eugenol attenuates concanavalin A-induced hepatitis through modulation of cytokine levels and inhibition of mitochondrial oxidative stress. Arch. Biol. Sci. 2019, 71, 339–346. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Q.; Zhou, C.; Zhao, Y.; Li, R.; Zhang, Y. Demethyleneberberine attenuates concanavalin A-induced autoimmune hepatitis in mice through inhibition of NF-κB and MAPK signaling. Int. Immunopharmacol. 2020, 80, 106137. [Google Scholar] [CrossRef]
- Sang, X.X.; Wang, R.L.; Zhang, C.E.; Liu, S.J.; Shen, H.H.; Guo, Y.M.; Zhang, Y.M.; Niu, M.; Wang, J.B.; Bai, Z.F.; et al. Sophocarpine protects mice from con A-induced hepatitis via inhibition of the IFN-gamma/STAT1 pathway. Front. Pharmacol. 2017, 8, 00140. [Google Scholar] [CrossRef]
- Tu, C.T.; Han, B.; Liu, H.C.; Zhang, S.C. Curcumin protects mice against concanavalin A-induced hepatitis by inhibiting intrahepatic intercellular adhesion molecule-1 (ICAM-1) and CXCL10 expression. Mol. Cell Biochem. 2011, 358, 53–60. [Google Scholar] [CrossRef] [PubMed]
- El-Agamy, D.S.; Shaaban, A.A.; Almaramhy, H.H.; Elkablawy, S.; Elkablawy, M.A. Pristimerin as a novel hepatoprotective agent against experimental autoimmune hepatitis. Front. Pharmacol. 2018, 9, 292. [Google Scholar] [CrossRef] [PubMed]
- Mo, W.; Wang, C.; Li, J.; Chen, K.; Xia, Y.; Li, S.; Xu, L.; Lu, X.; Wang, W.; Guo, C. Fucosterol protects against concanavalin A-induced acute liver injury: Focus on P38 MAPK/NF-κB pathway activity. Gastroenterol. Res. Pract. 2018, 2018, 2824139. [Google Scholar] [CrossRef]
- Mao, Y.; Wang, J.; Yu, F.; Cheng, J.; Li, H.; Guo, C.; Fan, X. Ghrelin reduces liver impairment in a model of concanavalin A-induced acute hepatitis in mice. Drug Des. Devel. Ther. 2015, 9, 5385–5396. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.; Pattanaik, A.K.; Das, J.; Bhutia, S.K.; Behera, B.; Singh, P.; Maiti, T.K. Role of PI3K/Akt/mTOR and MEK/ERK pathway in Concanavalin A induced autophagy in HeLa cells. Chem. Biol. Interact. 2014, 210, 96–102. [Google Scholar] [CrossRef]
- Meng, Z.; Wang, J.; Yuan, Y.; Cao, G.; Fan, S.; Gao, C.; Wang, L.; Li, Z.; Wu, X.; Wu, Z.; et al. γδ T cells are indispensable for interleukin-23-mediated protection against Concanavalin A-induced hepatitis in hepatitis B virus transgenic mice. Immunology 2017, 151, 43–55. [Google Scholar] [CrossRef]
- Roth, R.A.; Cassell, D.J.; Maddux, B.A.; Goldfine, I.D. Regulation of insulin receptor kinase activity by insulin mimickers and an insulin antagonist. Biochem. Biophys. Res. Commun. 1983, 115, 245–252. [Google Scholar] [CrossRef]
- Asherson, G.L.; Ferluga, J.; Janossy, G. Non-specific cytotoxicity by T cells activated with plant mitogens in vitro and the requirement for plant agents during the killing reaction. Clin. Exp. Immunol. 1973, 15, 573–589. [Google Scholar]
- Heymann, F.; Hamesch, K.; Weiskirchen, R.; Tacke, F. The concanavalin A model of acute hepatitis in mice. Lab. Anim. 2015, 49, 12–20. [Google Scholar] [CrossRef]
- Qi, C.; Tan, X.; Shi, Z.; Feng, H.; Sun, L.; Hu, Z.; Chen, G.; Zhang, Y. Discovery of an oxepine-containing diketopiperazine derivative active against concanavalin A-induced hepatitis. J. Nat. Prod. 2020, 83, 2672–2678. [Google Scholar] [CrossRef]
- Wang, Y.; Zhou, L.; Li, Y.; Guo, L.; Zhou, Z.; Xie, H.; Hou, Y.; Wang, B. The effects of berberine on concanavalin A-induced autoimmune hepatitis (AIH) in mice and the adenosine 5’-monophosphate (AMP)-activated protein kinase (AMPK) pathway. Med. Sci. Monit. 2017, 23, 6150–6161. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Zhu, Y.N.; Feng, J.Q.; Chen, H.J.; Zhang, R.J.; Ni, J.; Chen, Z.H.; Hou, L.F.; Liu, Q.F.; Zhang, J.; et al. Periplocoside A, a pregnane glycoside from Periploca sepium Bge, prevents concanavalin A-induced mice hepatitis through inhibiting NKT-derived inflammatory cytokine productions. Int. Immunopharmacol. 2008, 8, 1248–1256. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wang, J.; Liu, R.; Wang, Z.; Li, Y.; Zhang, Y.; Hao, X.; Huang, Y.; Xie, W.; Wei, H. Amelioration of concanavalin A-induced autoimmune hepatitis by magnesium isoglycyrrhizinate through inhibition of CD4(+)CD25(-)CD69(+) subset proliferation. Drug Des. Devel. Ther. 2016, 10, 443–453. [Google Scholar] [PubMed]
- Manns, M.P.; Lohse, A.W.; Vergani, D. Autoimmune hepatitis—Update 2015. J. Hepatol. 2015, 62, S100–S111. [Google Scholar] [CrossRef] [PubMed]
- Christen, U.; Hintermann, E. Immunopathogenic mechanisms of autoimmune hepatitis: How much do we know from animal models? Int. J. Mol. Sci. 2016, 17, 2007. [Google Scholar] [CrossRef]
- Christen, U. Animal models of autoimmune hepatitis. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 970–981. [Google Scholar] [CrossRef]
- AlSaadi, B.H.; AlHarbi, S.H.; Ibrahim, S.R.M.; El-Kholy, A.A.; El-Agamy, D.S.; Mohamed, G.A. Hepatoprotective activity of Costus speciosus against paracetamol-induced liver injury in mice. Afr. J. Tradit. Complement Altern. Med. 2018, 15, 35–41. [Google Scholar]
- Syed, S.H.; Namdeo, A.G. Current status of natural products for the treatment of liver disease-A review. Int. J. Phytopharm. 2014, 4, 37–43. [Google Scholar]
- Zhang, A.; Sun, H.; Wang, X. Recent advances in natural products from plants for treatment of liver diseases. Eur. J. Med. Chem. 2013, 63, 570–577. [Google Scholar] [CrossRef]
- Ibrahim, S.R.M.; El-Agamy, D.S.; Abdallah, H.M.; Ahmed, N.; Elkablawy, M.A.; Mohamed, G.A. Protective activity of tovophyllin A, a xanthone isolated from Garcinia mangostana pericarps, against acetaminophen-induced liver damage: Role of Nrf2 activation. Food Funct. 2018, 9, 3291–3300. [Google Scholar] [CrossRef]
- Qian, H.; Shi, J.; Fan, T.T.; Lv, J.; Chen, S.W.; Song, C.Y.; Zheng, Z.W.; Xie, W.F.; Chen, Y.X. Sophocarpine attenuates liver fibrosis by inhibiting the TLR4 signaling pathway in rats. World J. Gastroenterol. 2014, 20, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Mei, Y.; Wang, Y.; Zhang, B.; Wang, C.; Xu, L. Tetrandrine protects mice from concanavalin A-induced hepatitis through inhibiting NF-kappaB activation. Immunol. Lett. 2008, 121, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, J.; Wang, J.; Wang, L.; Yin, H.; Tan, R.; Xu, Q. Fumigaclavine C improves concanavalin A-induced liver injury in mice mainly via inhibiting TNF-alpha production and lymphocyte adhesion to extracellular matrices. J. Pharm. Pharmacol. 2004, 56, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zhang, B.; Shen, R.W.; Liu, J.B.; Gao, M.H.; Li, Y.; Li, Y.Y.; Zhang, W. Preventive effect of halofuginone on concanavalin A-induced liver fibrosis. PLoS ONE 2013, 8, e82232. [Google Scholar] [CrossRef]
- Tan, X.; Sun, L.; Li, Q.; Qi, C.; Fu, C.; Zhu, H.; Yang, X.; Feng, H.; Li, Y.; Zhang, Y.; et al. Secoemestrin C inhibits activation of NKT/conventional T cells and protects against concanavalin A-induced autoimmune hepatitis in mice. Am. J. Transl. Res. 2020, 12, 3389–3401. [Google Scholar]
- Zhao, J.; Park, S.; Kim, J.W.; Qi, J.; Zhou, Z.; Lim, C.W.; Kim, B. Nicotine attenuates concanavalin A-induced liver injury in mice by regulating the α7-nicotinic acetylcholine receptor in Kupffer cells. Int. Immunopharmacol. 2020, 78, 106071. [Google Scholar] [CrossRef]
- Wang, T.; Zhao, J.; Li, X.; Xu, Q.; Liu, Y.; Khan, I.A.; Yang, S. New alkaloids from green vegetable soybeans and their inhibitory activities on the proliferation of concanavalin A-activated lymphocytes. J. Agric. Food Chem. 2016, 64, 1649–1656. [Google Scholar] [CrossRef]
- Chen, M.; Cao, L.; Luo, Y.; Feng, X.; Sun, L.; Wen, M.; Peng, S. Paeoniflorin protects against concanavalin A-induced hepatitis in mice. Int. Immunopharmacol. 2015, 24, 42–49. [Google Scholar] [CrossRef]
- Wang, D.; Wang, H.; Fu, S.; Cheng, X.; Yang, F.; Zhang, Q.; Li, Y.; Xue, Z.; Zhang, L.; Huang, W.; et al. Parthenolide ameliorates concanavalin A-induced acute hepatitis in mice and modulates the macrophages to an anti-inflammatory state. Int. Immunopharmacol. 2016, 38, 132–138. [Google Scholar] [CrossRef]
- Zhou, Y.Q.; Weng, X.F.; Dou, R.; Tan, X.S.; Zhang, T.T.; Fang, J.B.; Wu, X.W. Betulin from Hedyotis hedyotidea ameliorates concanavalin A-induced and T cell-mediated autoimmune hepatitis in mice. Acta Pharmacol. Sin. 2017, 38, 201–210. [Google Scholar] [CrossRef]
- Zhou, R.; Tang, W.; Ren, Y.X.; He, P.L.; Yang, Y.F.; Li, Y.C.; Zuo, J.P. Preventive effects of (5R)-5-hydroxytriptolide on concanavalin A-induced hepatitis. Eur. J. Pharmacol. 2006, 537, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.T.; Li, J.; Wang, F.P.; Li, L.; Wang, J.Y.; Jiang, W. Glycyrrhizin regulates CD4+T cell response during liver fibrogenesis via JNK, ERK and PI3K/AKT pathway. Int. Immunopharmacol. 2012, 14, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Liu, Y.; Liu, X.; Gao, S.; Sun, X. Glycyrrhizic acid ammonium salt alleviates concanavalin A-induced immunological liver injury in mice through the regulation of the balance of immune cells and the inhibition of hepatocyte apoptosis. Biomed. Pharmacother. 2019, 120, 109481. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Zheng, L. Magnesium isoglycyrrhizinate protects against concanavalin A-induced immunological liver injury in a mouse model. Rev. Română Med. Lab. 2019, 27, 281–290. [Google Scholar] [CrossRef]
- Klein, C.; Wüstefeld, T.; Heinrich, P.C.; Streetz, K.L.; Manns, M.P.; Trautwein, C. ME3738 protects from concanavalin A-induced liver failure via an IL-6-dependent mechanism. Eur. J. Immunol. 2003, 33, 2251–2261. [Google Scholar] [CrossRef]
- Kuzuhara, H.; Nishiyama, S.; Minowa, N.; Sasaki, K.; Omoto, S. Protective effects of soyasapogenol A on liver injury mediated by immune response in a concanavalin A-induced hepatitis model. Eur. J. Pharmacol. 2000, 391, 175–181. [Google Scholar] [CrossRef]
- Sun, C.; Jiang, M.; Zhang, L.; Yang, J.; Zhang, G.; Du, B.; Ren, Y.; Li, X.; Yao, J. Cycloastragenol mediates activation and proliferation suppression in concanavalin A-induced mouse lymphocyte pan-activation model. Immunopharmacol. Immunotoxicol. 2017, 39, 131–139. [Google Scholar] [CrossRef]
- Sang, R.; Yu, Y.; Ge, B.; Xu, L.; Wang, Z.; Zhang, X. Taraxasterol from Taraxacum prevents concanavalin A-induced acute hepatic injury in mice via modulating TLRs/NF-κB and Bax/Bc1-2 signalling pathways. Artif. Cells Nanomed. Biotechnol. 2019, 47, 3929–3937. [Google Scholar] [CrossRef]
- Xie, Y.; Sun, H.X.; Li, D. Platycodin D is a potent adjuvant of specific cellular and humoral immune responses against recombinant hepatitis B antigen. Vaccine 2009, 27, 757–764. [Google Scholar] [CrossRef]
- Xie, Y.; He, S.W.; Sun, H.X.; Li, D. Platycodin D2 improves specific cellular and humoral responses to hepatitis B surface antigen in mice. Chem. Biodivers. 2010, 7, 178–185. [Google Scholar] [CrossRef]
- Chen, F.Y.; Zhou, L.F.; Li, X.Y.; Zhao, J.W.; Xu, S.F.; Huang, W.H.; Gao, L.J.; Hao, S.J.; Ye, Y.P.; Sun, H.X. Stephanthraniline A suppressed CD4+ T cell-mediated immunological hepatitis through impairing PKCθ function. Eur. J. Pharmacol. 2016, 789, 370–384. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, K.; Iwaki, T.; Kinoshita, S.; Koyama, M.; Fukunari, A.; Tanaka, H.; Tsurufuji, M.; Sakata, K.; Maeda, Y.; Imada, T.; et al. Ursodeoxycholic acid protects concanavalin A-induced mouse liver injury through inhibition of intrahepatic tumor necrosis factor-alpha and macrophage inflammatory protein-2 production. Eur. J. Pharmacol. 2008, 578, 57–64. [Google Scholar] [CrossRef]
- Chen, Y.F.; Wang, S.H.; Chang, S.J.; Shiau, A.L.; Her, L.S.; Shieh, G.S.; Chen, C.F.; Chang, C.C.; Su, Y.C.; Wu, C.L.; et al. Zhankuic acid A as a novel JAK2 inhibitor for the treatment of concanavalin A-induced hepatitis. Biochem. Pharmacol. 2014, 91, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zou, Y.; Liu, S.; Wang, J.; Zhu, J.; Li, J.; Bo, L.; Deng, X. Salidroside attenuates concanavalin A-induced hepatitis via modulating cytokines secretion and lymphocyte migration in mice. Mediat. Inflamm. 2014, 2014, 314081. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Chen, K.; He, L.; Xia, Y.; Dai, W.; Wang, F.; Li, J.; Li, S.; Liu, T.; Zheng, Y.; et al. The protective effect of resveratrol on concanavalin-A-induced acute hepatic injury in mice. Gastroenterol. Res. Pract. 2015, 2015, 506390. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Nie, H.; Li, K.; Zhang, Y.X.; Yang, F.; Li, C.B.; Wang, C.F.; Gong, Q. Curcumin inhibits HMGB1 releasing and attenuates concanavalin A-induced hepatitis in mice. Eur. J. Pharmacol. 2012, 697, 152–157. [Google Scholar] [CrossRef]
- Li, G.; Chen, M.J.; Wang, C.; Nie, H.; Huang, W.J.; Yuan, T.D.; Sun, T.; Shu, K.G.; Wang, C.F.; Gong, Q.; et al. Protective effects of hesperidin on concanavalin A-induced hepatic injury in mice. Int. Immunopharmacol. 2014, 21, 406–411. [Google Scholar] [CrossRef]
- Li, X.; Liu, H.C.; Yao, Q.Y.; Xu, B.L.; Zhang, S.C.; Tu, C.T. Quercetin protects mice from cona-induced hepatitis by inhibiting HMGB1-TLR expression and down-regulating the nuclear factor kappa B pathway. Inflammation 2016, 39, 96–106. [Google Scholar] [CrossRef]
- Liu, L.L.; Gong, L.K.; Wang, H.; Xiao, Y.; Wu, X.F.; Zhang, Y.H.; Xue, X.; Qi, X.M.; Ren, J. Baicalin protects mouse from concanavalin A-induced liver injury through inhibition of cytokine production and hepatocyte apoptosis. Liver Int. 2007, 27, 582–591. [Google Scholar] [CrossRef]
- Zhang, Y.; Shan, L.; Hua, Y.; Wang, D.; Zeng, H.; Liu, R.; Zhang, W.; Hu, Z. Baicalein selectively induces apoptosis in activated lymphocytes and ameliorates concanavalin a-induced hepatitis in mice. PLoS ONE 2013, 8, e69592. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, X.; Jiang, L.; Guo, Y.; Zheng, C. Epigallocatechin-3-gallate (EGCG) attenuates concanavalin A-induced hepatic injury in mice. Acta Histochem. 2014, 116, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Zhu, L.; Ding, J.; Zhuang, X.; Xu, L.; Chen, F. Protective effect of galangin in Concanavalin A-induced hepatitis in mice. Drug Des. Devel. Ther. 2015, 10, 2983–2992. [Google Scholar]
- Tan, Z.H.; Yu, L.H.; Wei, H.L.; Liu, G.T. The protective action of scutellarin against immunological liver injury induced by concanavalin A and its effect on pro-inflammatory cytokines in mice. J. Pharm. Pharmacol. 2007, 59, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhao, Y.; Xu, Q. Astilbin prevents concanavalin A-induced liver injury by reducing TNF-alpha production and T lymphocytes adhesion. J. Pharm. Pharmacol. 2004, 56, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Chen, J.; Zhu, P.; Fujino, M.; Takaharam, T.; Toyama, S.; Tomita, A.; Zhao, L.; Yang, Z.; Hei, M.; et al. Dihydroquercetin (DHQ) ameliorated concanavalin A-induced mouse experimental fulminant hepatitis and enhanced HO-1 expression through MAPK/Nrf2 antioxidant pathway in RAW cells. Int. Immunopharmacol. 2015, 28, 938–944. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Xia, Y.; Li, J.; Li, S.; Feng, J.; Wu, L.; Zhang, R.; Xu, S.; Cheng, K.; Zhou, Y.; et al. Shikonin attenuates concanavalin A-induced acute liver injury in mice via inhibition of the JNK pathway. Mediat. Inflamm. 2016, 2016, 2748367. [Google Scholar] [CrossRef]
- Zhang, H.; Ju, B.; Zhang, X.; Zhu, Y.; Nie, Y.; Xu, Y.; Lei, Q. Magnolol attenuates concanavalin A-induced hepatic fibrosis, inhibits CD4+ T helper 17 (Th17) cell differentiation and suppresses hepatic stellate cell activation: Blockade of Smad3/Smad4 signalling. Basic Clin. Pharmacol. Toxicol. 2017, 120, 560–570. [Google Scholar] [CrossRef]
- Wu, J.; Li, M.; He, J.; Lv, K.; Wang, M.; Guan, W.; Liu, J.; Tao, Y.; Li, S.; Ho, C.T.; et al. Protective effect of pterostilbene on concanavalin A-induced acute liver injury. Food Funct. 2019, 10, 7308–7314. [Google Scholar] [CrossRef]
- Xu, X.; Hu, Y.; Zhai, X.; Lin, M.; Chen, Z.; Tian, X.; Zhang, F.; Gao, D.; Ma, X.; Lv, L.; et al. Salvianolic acid A preconditioning confers protection against concanavalin A-induced liver injury through SIRT1-mediated repression of p66shc in mice. Toxicol. Appl. Pharmacol. 2013, 273, 68–76. [Google Scholar] [CrossRef]
- Yuan, Y.; Gong, X.; Zhang, L.; Jiang, R.; Yang, J.; Wang, B.; Wan, J. Chlorogenic acid ameliorated concanavalin A-induced hepatitis by suppression of Toll-like receptor 4 signaling in mice. Int. Immunopharmacol. 2017, 44, 97–104. [Google Scholar] [CrossRef]
- Luo, Q.; Ding, J.; Zhu, L.; Chen, F.; Xu, L. Hepatoprotective effect of wedelolactone against concanavalin A-induced liver injury in mice. Am. J. Chin. Med. 2018, 46, 819–833. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Yoshida, S.; Kobayashi, T.; Okabe, S. Inhibition of concanavalin A-induced mice hepatitis by coumarin derivatives. Jpn. J. Pharmacol. 2001, 85, 95–97. [Google Scholar] [CrossRef] [PubMed][Green Version]
- You, L.; Feng, S.; An, R.; Wang, X. Osthole: A promising lead compound for drug discovery from a traditional Chinese medicine (TCM). Nat. Prod. Commun. 2009, 4, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Lin, L.; Cui, B.; Zhao, T.; Mao, L.; Song, Y.; Wang, X.; Feng, H.; Qingxiang, Y.; Zhang, J.; et al. Therapeutic potential of genipin in various acute liver injury, fulminant hepatitis, NAFLD and other non-cancer liver diseases: More friend than foe. Pharmacol. Res. 2020, 159, 104945. [Google Scholar] [CrossRef] [PubMed]
- Fei, M.; Xie, Q.; Zou, Y.; He, R.; Zhang, Y.; Wang, J.; Bo, L.; Li, J.; Deng, X. Alpha-lipoic acid protects mice against concanavalin A-induced hepatitis by modulating cytokine secretion and reducing reactive oxygen species generation. Int. Immunopharmacol. 2016, 35, 53–60. [Google Scholar] [CrossRef]
- Bruck, R.; Aeed, H.; Brazovsky, E.; Noor, T.; Hershkoviz, R. Allicin, the active component of garlic, prevents immune-mediated, concanavalin A-induced hepatic injury in mice. Liver Int. 2005, 25, 613–621. [Google Scholar] [CrossRef]
- Ren, J.; Meng, S.; Yan, B.; Yu, J.; Liu, J. Protectin D1 reduces concanavalin A-induced liver injury by inhibiting NF-κB-mediated CX3CL1/CX3CR1 axis and NLR family, pyrin domain containing 3 inflammasome activation. Mol. Med. Rep. 2016, 13, 3627–3638. [Google Scholar] [CrossRef]
- Sun, Q.; Xu, X.; Yang, X.; Weng, D.; Wang, J.; Zhang, J. Salecan protected against concanavalin A-induced acute liver injury by modulating T cell immune responses and NMR-based metabolic profiles. Toxicol. Appl. Pharmacol. 2017, 317, 63–72. [Google Scholar] [CrossRef]
- Saito, A.; Yoneda, M.; Yokohama, S.; Okada, M.; Haneda, M.; Nakamura, K. Fucoidan prevents concanavalin A-induced liver injury through induction of endogenous IL-10 in mice. Hepatol. Res. 2006, 35, 190–198. [Google Scholar] [CrossRef][Green Version]
- Huang, T.H.; Chen, C.C.; Liu, H.M.; Lee, T.Y.; Shieh, S.H. Resveratrol pretreatment attenuates concanavalin A-induced hepatitis through reverse of aberration in the immune response and regenerative capacity in aged mice. Sci. Rep. 2017, 7, 2705. [Google Scholar] [CrossRef]
- Tu, C.T.; Han, B.; Yao, Q.Y.; Zhang, Y.A.; Liu, H.C.; Zhang, S.C. Curcumin attenuates concanavalin A-induced liver injury in mice by inhibition of toll-like receptor (TLR) 2, TLR4 and TLR9 expression. Int. Immunopharmacol. 2012, 12, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Tu, C.T.; Yao, Q.Y.; Xu, B.L.; Zhang, S.C. Curcumin protects against concanavalin A-induced hepatitis in mice through inhibiting the cytoplasmic translocation and expression of high mobility group box 1. Inflammation 2013, 36, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Mase, A.; Makino, B.; Tsuchiya, N.; Yamamoto, M.; Kase, Y.; Takeda, S.; Hasegawa, T. Active ingredients of traditional Japanese (kampo) medicine, inchinkoto, in murine concanavalin A-induced hepatitis. J. Ethnopharmacol. 2010, 127, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Song, Y.H.; Xia, W.J.; Jin, M.W. Aqueous extract of Yin-Chen-Hao decoction, a traditional Chinese prescription, exerts protective effects on concanavalin A-induced hepatitis in mice through inhibition of NF-kappaB. J. Pharm. Pharmacol. 2006, 58, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Yamashiki, M.; Mase, A.; Arai, I.; Huang, X.X.; Nobori, T.; Nishimura, A.; Sakaguchi, S.; Inoue, K. Effects of the Japanese herbal medicine ‘Inchinko-to’ (TJ-135) on concanavalin A-induced hepatitis in mice. Clin. Sci. 2000, 99, 421–431. [Google Scholar] [CrossRef]
- Wang, K.; Song, Z.; Wang, H.; Li, Q.; Cui, Z.; Zhang, Y. Angelica sinensis polysaccharide attenuates concanavalin A-induced liver injury in mice. Int. Immunopharmacol. 2016, 31, 140–148. [Google Scholar] [CrossRef]
- Wang, F.; Xue, Y.; Yang, J.; Lin, F.; Sun, Y.; Li, T.; Wu, C. Hepatoprotective effect of apple polyphenols against concanavalin A-induced immunological liver injury in mice. Chem. Biol. Interact. 2016, 258, 159–165. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; States, V.A.; Li, S.; Zhang, X.; Martin, R.C. The effect of black raspberry extracts on MnSOD activity in protection against concanavalin A induced liver injury. Nutr Cancer. 2014, 66, 930–937. [Google Scholar] [CrossRef]
- Herath, K.H.I.N.M.; Bing, S.J.; Cho, J.; Kim, A.; Shin, S.; Kim, G.O.; Lee, J.C.; Jee, Y. Anti-inflammatory activities of Dangyuja (Citrus grandis Osbeck) in concanavalin A stimulated murine splenocytes and 12-O-tetradecanoylphorbol-13-acetate-induced murine skin edema. Biomed. Pharmacother. 2016, 83, 1353–1364. [Google Scholar] [CrossRef]
- Yim, T.K.; Ko, K.M. Effects of fructus Ligustri lucidi extracts on concanavalin A-stimulated proliferation of isolated murine splenocytes. Pharm. Biol. 2001, 39, 146–151. [Google Scholar] [CrossRef]
- Mounieb, F.; Ramadan, L.; Akool, E.S.; Balah, A. Propolis alleviates concanavalin A-induced hepatitis by modulating cytokine secretion and inhibition of reactive oxygen species. Naunyn Schmiedebergs Arch. Pharmacol. 2017, 390, 1105–1115. [Google Scholar] [CrossRef] [PubMed]
- Zhai, K.F.; Duan, H.; Cao, W.G.; Gao, G.Z.; Shan, L.L.; Fang, X.M.; Zhao, L. Protective effect of Rabdosia amethystoides (Benth) Hara extract on acute liver injury induced by concanavalin A in mice through inhibition of TLR4-NF-κB signaling pathway. J. Pharmacol. Sci. 2016, 130, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.; Zhang, Y.; Zhao, J.; He, H.; Hou, T. Selenium-biofortified corn peptides: Attenuating concanavalin A-Induced liver injury and structure characterization. J. Trace Elem. Med. Biol. 2019, 51, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Ji, W.; Peng, X.; Lou, T.; Wang, J.; Qiu, W. Total flavonoids from Tetrastigma hemsleyanum ameliorates inflammatory stress in concanavalin A-induced autoimmune hepatitis mice by regulating Treg/Th17 immune homeostasis. Inflammopharmacology 2019, 27, 1297–1307. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Fan, J.; Ouyang, Z.; Ning, R.; Guo, W.; Shenm, Y.; Wu, X.; Sun, Y.; Xu, Q. Tupistra chinensis extract attenuates murine fulminant hepatitis with multiple targets against activated T lymphocytes. J. Pharm. Pharmacol. 2014, 66, 453–465. [Google Scholar] [CrossRef]
- Tian, M.; Liu, W.; You, H.; Zhao, Q.; Ouyang, L.; Gao, B.; Zhang, X.; Che, N. Protective effect of Yiguanjian decoction against DNA damage on concanavalin A-induced liver injury mice model. J. Tradit. Chin. Med. 2016, 36, 471–478. [Google Scholar]
- Tang, H.H.; Li, H.L.; Li, Y.X.; You, Y.; Guan, Y.Y.; Zhang, S.L.; Liu, L.X.; Bao, W.L.; Zhou, Y.; Shen, X.Y. Protective effects of a traditional Chinese herbal formula Jiang-Xian HuGan on concanavalin A-induced mouse hepatitis via NF-κB and Nrf2 signaling pathways. J. Ethnopharmacol. 2018, 217, 118–125. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Class | Mol. Formula | Mol. Weight | Plant/Fungus Name (Family) | Reference | ||
|---|---|---|---|---|---|---|---|
| Alkaloids | |||||||
| Sophocarpine | Matrine-type alkaloid | C16H24N2O | 260 | Sophorae Subprostrata = Sophora tonkinensis (Fabaceae) | [10] | ||
| Demethyleneberberine | Isoquinoline alkaloid | C19H18NO4+ | 325 | Phellodendron chinense (Rutaceae) | [9] | ||
| Berberine | Isoquinoline alkaloid | C20H18NO4+ | 337 | Coptis chinensis (Ranunculaceae) Phellodendron amurense (Rutaceae) | [21] | ||
| Capsaicin (8-methyl-N-vanillyl-6-nonenamide) * | Phenolic amide alkaloid | C18H27NO3 | 305 | - | [4] | ||
| Tetrandrine | bis-Benzylisoquinoline alkaloid | C38H42N2O6 | 622 | Stephania tetrandra (Menispermaceae) | [32] | ||
| Fumigaclavine C | Ergoline alkaloid | C23H30N2O2 | 366 | Aspergillus fumigatus (strain No. CY018) (Aspergillaceae) | [33] | ||
| Halofuginone | Quinazoline alkaloid | C16H17BrClN3O3 | 413 | Dichroa febrifuga (Hydrangeaceae) | [34] | ||
| Secoemestrin C | Epitetrathiodioxopiperazine alkaloid | C27H22N2O10S | 566 | Aspergillus nidulans (B, C) (Aspergillaceae) | [35] | ||
| Varioxepine B | Diketopiperazine alkaloid | C27H31N3O5 | 477 | Aspergillus terreus (strain No. CY018) (Aspergillaceae) | [20] | ||
| Nicotine * | Pyridine alkaloid | C10H14N2 | 162 | - | [36] | ||
| Soyalkaloid A | Indole alkaloid | C13H10N2O2 | 226 | Glycine max (Fabaceae) | [37] | ||
| Isoginsenine | Azepino-indole alkaloid | C13H14N2O2 | 230 | Glycine max (Fabaceae) | [37] | ||
| Ginsenine | Azepino-indole alkaloid | C13H14N2O2 | 230 | Glycine max (Fabaceae) | [37] | ||
| (1S,3S)-1-Methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid | Tetrahydro-β-carboline alkaloid | C13H14N2O2 | 230 | Glycine max (Fabaceae) | [37] | ||
| (1R,3S)-1-Methyl-1,2,3,4-tetrahydro-β-carboline-3-carboxylic acid | Tetrahydro-β-carboline alkaloid | C13H14N2O2 | 230 | Glycine max (Fabaceae) | [37] | ||
| Indole-3-carboxylic acid | Indole alkaloid | C9H7NO2 | 161 | Glycine max (Fabaceae) | [37] | ||
| Terpenes | |||||||
| Eugenol * | Phenolic monoterpene | C10H12O2 | 164 | - | [8] | ||
| Paeoniflorin | Monoterpene glucoside | C23H28O11 | 480 | Paeonia lactiflora (Paeoniaceae) | [38] | ||
| Parthenolide | Sesquiterpene lactone | C15H20O3 | 248 | Tanacetum parthenium (Asteraceae) | [39] | ||
| Betulin | Triterpenoid | C30H50O2 | 442 | Hedyotis hedyotidea (Rubiaceae) | [40] | ||
| (5R)-5-hydroxytriptolide | Diterpenoid epoxide | C20H24O7 | 376 | Tripterygium wilfordii (Celastraceae) | [41] | ||
| Pristimerin | Quinonemethide triterpenoid | C30H40O4 | 464 | - | [12] | ||
| Glycyrrhizin * | Triterpenoid | C42H62O16 | 822 | - | [5,42] | ||
| Glycyrrhizic acid ammonium salt * | Triterpenoidal acid salt | C42H62O16.NH3 | 839 | - | [43] | ||
| Magnesium isoglycyrrhizinate * | Triterpenoidal acid magnesium salt | C42H60MgO16 | 844 | - | [44] [23] | ||
| ME3738 * | Triterpenoid | C31H52O3 | 472 | - | [45] | ||
| Soyasapogenol A | Triterpenoid | C30H50O4 | 474 | Glycine max (Fabaceae) | [46] | ||
| Cycloastragenol * | Cycloartane triterpenoid | C30H50O5 | 490 | - | [47] | ||
| Taraxasterol | Triterpenoid | C30H50O | 426 | Taraxacum officinale (Asteraceae) | [48] | ||
| Periplocoside A | Pregnane glycoside | C73H116O27 | 1424 | Periploca sepium (Apocynaceae) | [22] | ||
| Platycodin D | Triterpenoid glycoside | C57H92O27 | 1208 | Platycodon grandiflorum (Campanulaceae) | [49] | ||
| Platycodin D2 | Triterpenoid glycoside | C63H102O33 | 1386 | Platycodon grandiflorum (Campanulaceae) | [50] | ||
| Stephanthraniline A | Polyoxypregnane | C31H43NO8 | 557 | Stephanotis mucronata (Asclepiadoideae) | [51] | ||
| Sterols | |||||||
| Fucosterol * | Steroid | C29H48O | 412 | - | [13] | ||
| Ursodeoxycholic * acid | Steroid | C24H40O4 | 392 | - | [52] | ||
| Zhankuic acid | Steroid acid | C29H42O6 | 478 | Taiwanofungus camphoratus | [53] | ||
| Phenolic compounds | |||||||
| Salidroside | Phenylpropanoid glycoside | C14H20O7 | 300 | Rhodiola rosea (Crassulaceae) | [54] | ||
| Resveratrol * | Stilbene | C14H12O3 | 228 | - | [55] | ||
| Curcumin * | Diarylheptanoid | C21H20O6 | 368 | - | [56] | ||
| Hesperidin * | Flavanone glycoside | C28H34O15 | 610 | - | [57] | ||
| Quercetin * | Flavone | C15H10O7 | 302 | - | [58] | ||
| Baicalin | Flavone | C21H18O11 | 446 | Scutellaria baicalensis (Lamiaceae) | [59] | ||
| Baicalein | Flavone | C15H10O5 | 270 | Scutellaria baicalensis (Lamiaceae) | [60] | ||
| (-)-Epigallocatechin-3-gallate | Flavan | C22H18O11 | 458 | Camellia sinensis (Theaceae) | [61] | ||
| Galangin | Flavonol | C15H10O5 | 270 | Alpinia officinarum (Zingiberaceae) | [62] | ||
| Scutellarin * | Glycosyloxyflavone | C21H18O12 | 462 | - | [63] | ||
| Astilbin | Flavanonol glucoside | C21H22O11 | 450 | Smilacis Glabra (Smilacaceae) | [64] | ||
| Dihydroquercetin * | Flavanonol | C15H12O7 | 290 | - | [65] | ||
| Shikonin * | Naphthoquinone | C16H16O5 | 288 | - | [66] | ||
| Magnolol | Lignan | C18H18O2 | 266 | Magnolia officinalis (Magnoliaceae) | [67] | ||
| Pterostilbene * | Stilbene | C16H16O3 | 256 | - | [68] | ||
| Salvianolic acid | Phenolic acid | C26H22O10 | 494 | Salvia miltiorrhiza (Lamiaceae) | [69] | ||
| Chlorogenic acid * | Phenolic acid | C16H18O9 | 354 | - | [70] | ||
| Coumarins and coumarin derivatives | |||||||
| Wedelolactone | Coumarin | C16H10O7 | 314 | Eclipta prostrata (Asteraceae) | [71] | ||
| Osthole | Coumarin | C15H16O3 | 224 | Cnidium monnieri (Apiaceae) | [72] | ||
| [73] | |||||||
| Imperatorin | Psoralen | C16H14O4 | 270 | Cnidium monnieri (Apiaceae) | [73] | ||
| Pd-Ia | Coumarin | C21H22O7 | 386 | Angelica decursiva (Apiaceae) | [72] | ||
| Pd-II | Coumarin | C24H26O7 | 426 | Angelica decursiva (Apiaceae) | [72] | ||
| Pd-III | Coumarin | C24H28O7 | 428 | Angelica decursiva (Apiaceae) | [72] | ||
| Other compounds | |||||||
| Genipin | Iridoid | C11H14O5 | 226 | Gardenia jasminoides (Rubiaceae) | [74] | ||
| α-Lipoic acid * | Organosulfur compound | C8H14O2S2 | 206 | - | [75] | ||
| Allicin | Organosulfur compound | C6H10OS2 | 162 | Allium sativum (Amaryllidaceae) | [76] | ||
| Protectin D1 * | Dihydroxydocosahexaenoic acid | C22H32O4 | 360 | - | [77] | ||
| Salecan | Polysaccharide | 2 × 106 Da | Agrobacterium sp.ZX09 (Rhizobiaceae) | [78] | |||
| Fucoidan | Sulfated polysaccharide | 13 kDa to 950 kDa | Fucus vesiculosus (Fucaceae) | [79] | |||
| Compound Name | Measured Parameter | Results of Compound on Con A-Induced Change (Conc.) | Positive Control and/or Con A Model | Ref. |
|---|---|---|---|---|
| Sophocarpine (SC) | ALT | 398.02 U/L SC (30 mg/kg) 171.25 U/L SC (60 mg/kg) | 628.12 U/L Bicyclol (100 mg/kg) 957.92 U/L Con A (15 mg/kg) | [10] |
| AST | 157.69 U/L SC (30 mg/kg) 84.46 U/L SC (60 mg/kg) | 219.81 U/L Bicyclol (100 mg/kg) 344.98 U/L Con A (15 mg/kg) | ||
| TBIL | 5.05 U/L SC (30 mg/kg) 3.21 U/L SC (60 mg/kg) | 5.08 U/L Bicyclol (100 mg/kg) 9.99 U/L Con A (15 mg/kg) | ||
| % TUNEL-positive cells | 8.43% SC (30 mg/kg) 4.33% SC (60 mg/kg) | 6.23% Bicyclol (100 mg/kg) 13.53% Con A (15 mg/kg) | ||
| Relative MIP-1α expression | 16.80-fold SC (30 mg/kg) 3.32-fold SC (60 mg/kg) | 41.47-fold Con A (15 mg/kg) | ||
| Relative CXCL10 expression | 57.05-fold SC (30 mg/kg) 18.56-fold SC (60 mg/kg) | 123.72-fold Con A (15 mg/kg) | ||
| Relative ICAM-1 expression | 20.69-fold SC (30 mg/kg) 3.35-fold SC (60 mg/kg) | 43.44-fold Con A (15 mg/kg) | ||
| IFN-γ | 2831.00 pg/mL SC (30 mg/kg) 2030.00 pg/mL SC (60 mg/kg) | 3248.00 pg/mL Con A (15 mg/kg) | ||
| TNF-α | 1009.78 pg/mL SC (30 mg/kg) 540.99 pg/mL SC (60 mg/kg) | 998.35 pg/mL Con A (15 mg/kg) | ||
| % CD4+IFN-γ+ | 1.87% SC (30 mg/kg) 1.41% SC (60 mg/kg) | 2.89% Con A (15 mg/kg) | ||
| Expression of T-bet | 36.81 SC (30 mg/kg) 27.99 SC (60 mg/kg) | 67.74 pg/mL Con A (15 mg/kg) | ||
| Activation of STAT1 | 75.72 SC (30 mg/kg) 8.21 SC (60 mg/kg) | 205.49 pg/mL Con A (15 mg/kg) | ||
| Overexpression of SOCS1 | 2.21-fold SC (30 mg/kg) 3.83-fold SC (60 mg/kg) | 1.76 pg/mL Con A (15 mg/kg) | ||
| Demethyleneberberine (DMB) | ALT | ↓ 91% DMB (30 mg/kg) | ↑ 75-fold Con A (20 mg/kg) | [9] |
| AST | ↓ 85% DMB (30 mg/kg) | ↑ 19-fold Con A (20 mg/kg) | ||
| LDH | ↓ 48% DMB (30 mg/kg) | ↑ 8-fold Con A (20 mg/kg) | ||
| MDA | ↓ 29% DMB (7.5 mg/kg) | ↑ 127% Con A (20 mg/kg) | ||
| ↓ 40% DMB (15 mg/kg) | ||||
| ↓ 48% DMB (30 mg/kg) | ||||
| ALB | ↑ 16% DMB (15 mg/kg) | ↓ 33% Con A (20 mg/kg) | ||
| GSH | ↑ 45% DMB (7.5 mg/kg) | ↓ 39% Con A (20 mg/kg) | ||
| ↑ 61% DMB (15 mg/kg) | ||||
| ↑ 90% DMB (30 mg/kg) | ||||
| TNF-α | ↓ 64% DMB (15 mg/kg) | ↑ 22-fold Con A (20 mg/kg) | ||
| IL-6 | ↓ 75% DMB (15 mg/kg) | ↑ 42-fold Con A (20 mg/kg) | ||
| IL-1β | ↓ 40% DMB (15 mg/kg) | ↑ 21-fold Con A (20 mg/kg) | ||
| IFN-γ | ↓ 85% DMB (15 mg/kg) | ↑ 58-fold Con A (20 mg/kg) | ||
| Fumigaclavine C (FCC) | ALT | 3387 Karmen unit FCC (5 mg/kg) | 650 Karmen unit Ciclosporin (10 mg/kg) 6288 Karmen unit Con A (18 mg/kg) | [33] |
| 1078 Karmen unit FCC (10 mg/kg) | ||||
| 447 Karmen unit FCC (20 mg/kg) | ||||
| AST | 4213 Karmen unit FCC (5 mg/kg) | 1509 Karmen unit Ciclosporin (10 mg/kg) 8632 Karmen unit Con A (18 mg/kg) | ||
| 1423 Karmen unit FCC (10 mg/kg) | ||||
| 921 Karmen unit FCC (20 mg/kg) | ||||
| Hepatocyte necrosis * | 1 (0–2) FCC (5 mg/kg) | 0 (0–2) Ciclosporin (10 mg/kg) 2 (1–2) Con A (18 mg/kg) | ||
| 0 (0–1) FCC (10 mg/kg) | ||||
| 0 (0–1) FCC (20 mg/kg) | ||||
| Hepatocyte degeneration * | 2 (1–3) FCC (5 mg/kg) | 1 (0–3) Ciclosporin (10 mg/kg) 3 (1–3) Con A (18 mg/kg) | ||
| 1 (0–2) FCC (10 mg/kg) | ||||
| 1 (0–1) FCC (20 mg/kg) | ||||
| Inflammatory infiltration * | 1 (1–2) FCC (5 mg/kg) | 1 (1–2) Ciclosporin (10 mg/kg) 2 (1–3) Con A (18 mg/kg) | ||
| 1 (1–2) FCC (10 mg/kg) | ||||
| 1 (0–2) FCC (20 mg/kg) | ||||
| Kupffer cell hyperplasia * | 1 (1–2) FCC (5 mg/kg) | 2 (1–2) Ciclosporin (10 mg/kg) 1 (0–2) Con A (18 mg/kg) | ||
| 1 (1–2) FCC (10 mg/kg) | ||||
| 1 (0–1) FCC (20 mg/kg) | ||||
| Spleen weights | 162.7 mg FCC (5 mg/kg) | 124.4 mg Ciclosporin (10 mg/kg) 210.9 mg Con A (18 mg/kg) | ||
| 121.4 mg FCC (10 mg/kg) | ||||
| 111.6 mg FCC (20 mg/kg) | ||||
| Halofuginone (HF) | Liver-to-bodyweight ratio (LBWR) | 4.86 HF (10 ppm) | 5.42 Con A (17.5mg/kg) | [34] |
| ALT | 59.29 IU/L HF (10 ppm) | 73.37 IU/L Con A (17.5 mg/kg) | ||
| AST | 193.02 IU/L HF (10 ppm) | 274.64 IU/L Con A (17.5 mg/kg) | ||
| ALB | 31.17 g/L HF (10 ppm) | 27.58 g/L Con A (17.5 mg/kg) | ||
| Semi-quantitative scoring system (SSS) value | 13 HF (10 ppm) | 10 Con A (17.5 mg/kg) | ||
| Soyalkaloid A (SAA) | Proliferation of Con A-activated lymphocytes | 0.40 SAA (0.25 µM) | 0.28 Cyclosporine A (1 µM) 0.43 Con A (1 µg/mL) | [37] |
| 0.36 SAA (0.5 µM) | 0.28 Cyclosporine A (1 µM) 0.43 Con A (1 µg/mL) | |||
| 0.32 SAA (1 µM) | 0.28 Cyclosporine A (1 µM) 0.43 Con A (1 µg/mL) | |||
| 0.29 SAA (2 µM) | 0.28 Cyclosporine A (1 µM) 0.43 Con A (1 µg/mL) | |||
| (5R)-5-Hydroxytriptolide (LLDT-8) | Survival rat | 83% LLDT-8 (0.5 mg/kg) | 40% Con A (30 mg/kg) | [41] |
| 86% LLDT-8 (1 mg/kg) | ||||
| 100% LLDT-8 (2 mg/kg) | ||||
| ALT | 952 U/L LLDT-8 (2 mg/kg) | 2304 U/L Con A (30 mg/kg) | ||
| IFN-γ | 1508 pg/mL LLDT-8 (2 mg/kg) | 4375 pg/mL Con A (30 mg/kg) | ||
| Magnesium isoglycyrrhizinate (MIG) | ALT | 126.73 U/L MIG (50 mg/kg) | 82.38 U/L Dexamethasone (2.5 mg/kg) 341.19 U/L Con A (20 mg/kg) | [44] |
| 156.78 U/L MIG (25 mg/kg) | ||||
| 212.40 U/L MIG (12.5 mg/kg) | ||||
| AST | 227.50 U/L MIG (50 mg/kg) | 169.2 U/L Dexamethasone (2.5 mg/kg) 477.71 U/L Con A (20 mg/kg) | ||
| 279.39 U/L MIG (25 mg/kg) | ||||
| 336.0 U/L MIG (12.5 mg/kg) | ||||
| MDA | 59.68 mmol/mg Prot MIG (50 mg/kg) | 57.40 mmol/mg Prot Dexamethasone (2.5 mg/kg) 92.81 mmol/mg Prot Con A (20 mg/kg) | ||
| 57.48 mmol/mg Prot MIG (25 mg/kg) | ||||
| 70.31 mmol/mg Prot MIG (12.5 mg/kg) | ||||
| SOD | 287.01 U/mg Prot MIG (50 mg/kg) | 280.60 U/mg Prot Dexamethasone (2.5 mg/kg) 168.87 U/mg Prot Con A (20 mg/kg) | ||
| 295.97 U/mg Prot MIG (25 mg/kg) | ||||
| 279.84 U/mg Prot MIG (12.5 mg/kg | ||||
| TNF-α | 348.69 pg/mL MIG (50 mg/kg) | 183.44 pg/mL Dexamethasone (2.5 mg/kg) 498.65 pg/mL Con A (20 mg/kg) | ||
| 301.77 pg/mL MIG (25 mg/kg) | ||||
| 311.40 pg/mL MIG (12.5 mg/kg) | ||||
| IFN-γ | 577.70 pg/mL MIG (50 mg/kg) | 1786.73 pg/mL Dexamethasone (2.5 mg/kg) 182.33 pg/mL Con A (20 mg/kg) | ||
| 466.53 pg/mL MIG (25 mg/kg) | ||||
| 792.51 pg/mL MIG (12.5 mg/kg) | ||||
| Cycloastragenol (CAG) | CD69+ expression | 50.22% CAG (5.0 µM) | 68.86% Con A (5 µg/mL) | [47] |
| 49.39% CAG (10 µM) | ||||
| 38.33% CAG (20 µM) | ||||
| CD25+ expression | 67.29 % CAG (5.0 µM) | 90.45% Con A (5 µg/mL) | ||
| 49.23% CAG (10 µM) | ||||
| 36.66% CAG (20 µM) | ||||
| Con A-stimulated lymphocyte proliferation (PI) | 1.35 CAG (5.0 µM) | 1.43 Con A (5 µM) | ||
| 1.29 CAG (10 µM) | ||||
| 1.07 CAG (20 µM) | ||||
| Eugenol | SOD | 0.39 U/mg protein Eugenol (5 mg/kg) | 0.25 U/mg protein Con A (12 mg/kg) | [8] |
| CAT | 44.5 nmol H2O2 utilized/mg protein Eugenol (5 mg/kg) | 31.4 nmol H2O2 utilized/mg protein Con A (12 mg/kg) | ||
| GPx | 6.41 nmol GSH utilized/mg protein Eugenol (5 mg/kg) | 4.11 nmol GSH utilized/mg protein Con A (12 mg/kg) | ||
| GR | 19 nmol NADPH oxidized/min/mg protein Eugenol (5 mg/kg) | 11 nmol NADPH oxidized/min/mg protein Con A (12 mg/kg) | ||
| G6PD | 1.46 nmol inorganic phosphorus liberated/min/mg protein Eugenol (5 mg/kg) | 1.14 nmol inorganic phosphorus liberated/min/mg protein Con A (12 mg/kg) | ||
| Paeoniflorin | % CD4+ infiltration | 6.37% Paeoniflorin (50 mg/kg) | 17.4% Con A (15 mg/kg) | [38] |
| % CD8+ infiltration | 6.56% Paeoniflorin (50 mg/kg) | 13.3% Con A (15 mg/kg) | ||
| Amount NKT infiltration | 8.89 × 105 Paeoniflorin (50 mg/kg) | 14.56 × 105 Con A (15 mg/kg) | ||
| Resveratrol (RSV) | ALT | 351.98 IU/L RSV (30 mg/kg) | 853.13 IU/L Con A (15 mg/kg) | [80] |
| AST | 295.86 IU/L RSV (30 mg/kg) | 753.98 IU/L Con A (15 mg/kg) | ||
| TNF-α | 159.73 pg/mL RSV (30 mg/kg) | 314.98 pg/mL Con A (15 mg/kg) | ||
| IL-6 | 215.87 pg/mL RSV (30 mg/kg) | 513.47 pg/mL Con A (15 mg/kg) | ||
| IFN-γ | 137.75 pg/mL RSV (30 mg/kg) | 285.95 pg/mL Con A (15 mg/kg) | ||
| MCP-1 | 203.13 pg/mL RSV (30 mg/kg) | 837.98 pg/mL Con A (15 mg/kg) | ||
| ALT | 253.73 IU/L RSV (10 mg/kg) | 389.40 IU/L Con A (20 mg/kg) | [55] | |
| 239.40 IU/L RSV (20 mg/kg) | ||||
| 205.52 IU/L RSV (30 mg/kg) | ||||
| AST | 250.49 IU/L RSV (10 mg/kg) | 450.74 IU/L Con A (20 mg/kg) | ||
| 259.89 IU/L RSV (20 mg/kg) | ||||
| 165.36 IU/L RSV (30 mg/kg) | ||||
| Knodell scores | 4.67 RSV (10 mg/kg) | 5.51 Con A (20 mg/kg) | ||
| 3.33 RSV (20 mg/kg) | ||||
| 3.0 RSV (30 mg/kg) | ||||
| Curcumin (CM) | ALT 2 h | 123.8 IU/L CM (200 mg/kg) | 155.0 IU/L Con A (20 mg/kg) | [11] |
| ALT 8 h | 629.2 IU/L CM (200 mg/kg) | 833.7 IU/L Con A (20 mg/kg) | ||
| ALT 24 h | 263.0 IU/L CM (200 mg/kg) | 435.7 IU/L Con A (20 mg/kg) | ||
| AST 2 h | 475.2 IU/L CM (200 mg/kg) | 469.8 IU/L Con A (20 mg/kg) | ||
| AST 8 h | 881.3 IU/L CM (200 mg/kg) | 1143.8 IU/L Con A (20 mg/kg) | ||
| AST 24 h | 377.8 IU/L CM (200 mg/kg) | 629.3 IU/L Con A (20 mg/kg) | ||
| Hesperidin (HDN) | T-cell activation ratio in vivo (after 6 h) | 70.05 HDN (1000 mg/kg) | 90.21 Con A (15 mg/kg) | [57] |
| T-cell activation ratio in vitro (after 12 h) | 64.59 HDN (1000 mg/kg) | 83.39 Con A (15 mg/kg) | ||
| Scutellarin | ALT | 410.7 U/L Scutellarin (50 mg/kg) | 550.0 U/L Con A (25 mg/kg) | [63] |
| 402.2 U/L Scutellarin (100 mg/kg) | ||||
| AST | 224.4 U/L Scutellarin (50 mg/kg) | 448.0 U/L Con A (25 mg/kg) | ||
| 229.9 U/L Scutellarin (100 mg/kg) | ||||
| Histological grade of liver injury | 1.6 Scutellarin (100 mg/kg) | 2.6 Con A (25 mg/kg) | ||
| TNF-α | 1082.6 pg/mL Scutellarin (50 mg/kg) | 1067.9 pg/mL Con A (25 mg/kg) | ||
| 477.1 pg/mL Scutellarin (100 mg/kg) | ||||
| NO2-/NO3- levels | 21.1 µM/L Scutellarin (50 mg/kg) | 24.8 µM/L Con A (25 mg/kg) | ||
| 13.5 µM/L Scutellarin (100 mg/kg) | ||||
| iNOS/β-actin | 1.48 Scutellarin (50 mg/kg) | 2.29 Con A (25 mg/kg) | ||
| 0.95 Scutellarin (100 mg/kg) | ||||
| IFN-γ/β-actin | 1.94 Scutellarin (50 mg/kg) | 2.50 Con A (25 mg/kg) | ||
| 1.85 Scutellarin (100 mg/kg) | ||||
| Fas/β-actin | 2.05 Scutellarin (50 mg/kg) | 2.82 Con A (25 mg/kg) | ||
| 1.81 Scutellarin (100 mg/kg) | ||||
| TNF-α/β-actin | 2.03 Scutellarin (50 mg/kg) | 3.00 Con A (25 mg/kg) | ||
| 1.13 scutellarin (100 mg/kg) | ||||
| Astilbin | Hepatocyte necrosis | 0.75 Scutellarin (20 mg/kg) | 0 Ciclosporin (10 mg/kg) 1.13 Con A (20 mg/kg) | [64] |
| 0.63 Astilbin (40 mg/kg) | ||||
| Hepatocyte degeneration | 1.5 Astilbin (20 mg/kg) | 0.25 Ciclosporin (10 mg/kg) 2.25 Con A (20 mg/kg) | ||
| 0.88 Astilbin (40 mg/kg) | ||||
| Inflammatory infiltration | 1.13 Astilbin (20 mg/kg) | 0.88 Ciclosporin (10 mg/kg) 1.5 Con A (20 mg/kg) | ||
| 0.75 Astilbin (40 mg/kg) | ||||
| Kupffer cell hyperplasia | 1 Astilbin (20 mg/kg) | 0.75 Ciclosporin (10 mg/kg) 1.38 Con A (20 mg/kg) | ||
| 0.75 Astilbin (40 mg/kg) | ||||
| Pterostilbene (PTE) | ALT | 390 U/L PTE (10 mg/kg) | 1400 U/L Con A (10 mg/kg) | [68] |
| 130 U/L PTE (40 mg/kg) | ||||
| AST | 500 U/L PTE (10 mg/kg) | 1700 U/L Con A (10 mg/kg) | ||
| 230 U/L PTE (40 mg/kg) | ||||
| TUNEL positive cells | ≈14% PTE (10 mg/kg) | ≈41% Con A (10 mg/kg) | ||
| ≈6% PTE (40 mg/kg) | ||||
| Ki67 positive area | ≈10% PTE (10 mg/kg) | ≈1% Con A (10 mg/kg) | ||
| ≈13% PTE (40 mg/kg) | ||||
| mRNA IFN-γ level | ↓≈2.7% PTE (10 mg/kg) ↓≈3.3% PTE (40 mg/kg) | - | ||
| mRNA TNF-α level | ↓≈3.1 PTE (10 mg/kg) ↓≈4.5% PTE (40 mg/kg) | - | ||
| TF | ↓≈19% PTE (10 mg/kg) | ↑≈43% Con A (10 mg/kg) | ||
| ↓≈6% PTE (40 mg/kg) | ||||
| Fibrin | ↓≈3.8 PTE (10 mg/kg) | - | ||
| ↓≈6.7% PTE (40 mg/kg) | ||||
| F4/80 staining | ↓≈11% PTE (10 mg/kg) | ↑≈42% Con A (10 mg/kg) | ||
| ↓≈5% PTE (40 mg/kg) | ||||
| Ursodeoxycholic acid (UDCA) | UDCA | 38.5 nmol/g liver UDCA (50 mg/kg) | 2.4 nmol/g liver Con A (20 mg/kg) | [52] |
| 87.8 nmol/g liver UDCA (150 mg/kg) | ||||
| α-MCA | 11.1 nmol/g liver UDCA (50 mg/kg) | 13.1 nmol/g liver Con A (20 mg/kg) | ||
| 18.7 nmol/g liver UDCA (150 mg/kg) | ||||
| β-MCA | 58.7 nmol/g liver UDCA (50 mg/kg) | 72.3 nmol/g liver Con A (20 mg/kg) | ||
| 49.7 nmol/g liver UDCA (150 mg/kg) | ||||
| HDCA | <1 nmol/g liver UDCA (50 mg/kg) | <1 nmol/g liver Con A (20 mg/kg) | ||
| <1 nmol/g liver UDCA (150 mg/kg) | ||||
| CA | 107.8 nmol/g liver UDCA (50 mg/kg) | 141.1 nmol/g liver Con A (20 mg/kg) | ||
| 105.5 nmol/g liver UDCA (150 mg/kg) | ||||
| CDCA | <1 nmol/g liver UDCA (50 mg/kg) | <1 nmol/g liver Con A (20 mg/kg) | ||
| <1 nmol/g liver UDCA (150 mg/kg) | ||||
| DCA | 6.5 nmol/g liver UDCA (50 mg/kg) | 10.0 nmol/g liver Con A (20 mg/kg) | ||
| 12.4 nmol/g liver UDCA (150 mg/kg) | ||||
| LCA | <1 nmol/g liver UDCA (50 mg/kg) | <1 nmol/g liver Con A (20 mg/kg) | ||
| <1 nmol/g liver UDCA (150 mg/kg) | ||||
| Total bile acids | 223.6 nmol/g liver UDCA (50 mg/kg) | 239.6 nmol/g liver Con A (20 mg/kg) | ||
| 275.7 nmol/g liver UDCA (150 mg/kg) |
| Name of Plant (Organ, Family)//Preparation | Tested Extract/Fraction (Major Constituents) | Extract/Fraction/Con A (Conc.) | Pharmacological Outcomes/Effects | Reference |
|---|---|---|---|---|
| Yin-Chen-Hao (YCH) prescription in China inchinkoto (ICKT) and Inchinko-To (TJ-135) in Japan Artemisia capillaris (Inchinko) (Herb, Asteraceae) Gardenia jasminoides (Sanshishi) (Fruit, Rubiaceae) Rheum palmatum (Daio) (Rhizome, Polygonaceae) | Aqueous extract (ICKTD) (capillarisin, genipin, and 6,7-dimethylesculetin) | ICKTD (500 mg/10 mL/kg) ICKTD (1000 mg/10 mL/kg) ICKTD (2000 mg/10 mL/kg) Con A (20 mg/kg) | ICKTD ↓ ALT, AST, and IFN-γ concentrations. ICKTD ↓ production of IFN-γ in con A-stimulated splenocyte culture in vitro. ICKTD suppressed IL-1β, IL-6, and IL-12p70 synthesis. ICKT ↓ nitrite release from IFN-γ-stimulated macrophages. | [83] |
| Aqueous extract (decoction) (YCHD) (0.32% scoparone, 0.96% geniposide and 0.25% rhein) | YCH (150 mg/kg) YCH (300 mg/kg) YCH (600 mg/kg) Con A (22 mg/kg) | YCHD ↓ elevation in ALT, AST, and LDH activity. YCHD ↓ liver DNA fragmentation and caspase-3 levels. YCHD inhibited in vitro and in vivo TNF-α production. YCHD blocked the activation of NF-κB. YCHD effect depended on ↓TNF-α production via inhibition of NF-κB activation. | [84] | |
| Hot-water extract (TJ-135) | TJ-135 (500 mg/10 mL/kg) TJ-135 (1000 mg/10 mL/kg) TJ-135 (2000 mg/10 mL/kg) Con A (14 mg/kg) | TJ-135 significantly ↓serum AST, ALT, and LDH levels. TJ-135 suppressed sub-massive hepatic necrosis accompanying inflammatory cell infiltration. TJ-135 ↓ serum levels of IL-12, IFN-γ, and IL-12. TJ-135 ↑ IL-10 serum and intrasplenic levels. TJ-135 inhibited the production of inflammatory cytokine and enhanced the production of anti-inflammatory cytokines. | [85] | |
| Angelica sinensis (Roots, Apiaceae) | Polysaccharide (arabinose, glucose, and galactose with molar ratio 1:2.5:7.5) (ASP) | ASP (1.6 mg/kg) ASP (6 mg/kg) Con A (15 mg/kg) | ASP attenuated Con A-induced liver injury through its anti-inflammatory and anti-oxidant actions. ASP ↓ ALT and AST levels. ASP ↓ pro-inflammatory cytokines: TNF-α, IFN-γ, IL-2 and IL-6 levels. ASP alleviated oxidative stress by ↓ MDA and ROS levels and ↑ SOD activity. ASP ↓ caspase-3-dependent apoptosis by Caspase-8 and JNK-mediated pathway. ASP inhibited IL-6/STAT3 and NF-κB signaling pathways activation. | [86] |
| Malus domestica (Fruits, Rosaceae) | Polyphenols (AP) (procyanidins, quercetin, (-)-epicatechin, (+)-catechin, phloretin, phloridzin, and chlorogenic acid) | AP (200 mg/kg) AP (400 mg/kg) AP (800 mg/kg) Con A (200 mg/kg) | AP ↓ ALT, AST, TP and Alb levels and A/G ratio. AP ↓ TNF-α and IFN-γ, ↓ NO content serum levels. AP inhibited oxidative DNA single-strand breaks. AP improved the abnormalities of MDA content, SOD activity, and GSH level. AP exerted a protective effect through activation of the antioxidant system and suppression of pro-inflammatory cytokines. | [87] |
| Rubus occidentalis (Fruit, Rosaceae) | Ethanol/H2O soluble extract (BRB-A) Hexane extract (BRB-B) Ethanol/H2O insoluble extract (BRB-C) | BRB-A (1.6 mg/kg) BRB-B (1.6 mg/kg) BRB-C (1.6 mg/kg) Con A (30 mg/kg) | BRB-A and BRB-C inhibited Con A induced liver injury. The BRB-A and BRB-C ↓ lipid peroxidation and NDA oxidative damage. BRB-A and BRB-C ↑ MnSOD activity but not the Cu/ZnSOD. BRB extracts protective action resulted from their antioxidant action. | [88] |
| Citrus grandis (Fruit, Rutaceae) | EtOH extract (ECGO) | ECGO (3.9, 7.8, 15.6, 31.3, 62.5,125, 250, 500, and 1000 µg/mL) Con A (10 µg/mL) | ECGO significantly ↓ CD44/CD62L+ T cell population and ↓ production of IL-2, IFN-γ and IL-4 in Con A-stimulated splenocytes. | [89] |
| Ligustrum lucidum (Fruit, Oleaceae) (FLL) | Aqueous fraction (FLL-Aq) Butanol fraction (FLL-Bu) Chloroform fraction (FLL-Ch) Petroleum ether fraction (FLL-Pe) | FLL-Aq (2 g/kg) FLL-Bu (1 g/kg) FLL-Ch (1 g/kg) FLL-Pe (1 g/kg) Con A (1, 2, 4 μg/mL) | FLL-Pe produced an immunostimulatory effect, as evidenced by the significant enhancement of the Con A-stimulated mitogenic response of splenocytes in in vitro culture. FLL-Bu and FLL-Aq inhibited the in vitro Con A-stimulated splenocyte proliferation. FLL-Pe or FLL-Ch in ex vivo experiments, enhanced the extent of Con A-stimulated splenocyte proliferation. FLL-Pe and FLL-Ch produced immuno-stimulatory action under ex vivo assay conditions, but only FLL-Pe produced a similar effect under in vitro conditions. | [90] |
| Propolis (resinous natural product from honeybees) | phenolic acids or their esters, flavonoids, terpenes, fatty acids, stilbenes, and β-steroids | Propolis (300 mg/kg) Con A (12 mg/kg) | Propolis significantly ↑ Alb level and ↓ ALT, AST, and total bilirubin levels. Propolis attenuated lipid peroxidation and ↑ GSH, SOD, and CAT activities in liver tissue. Propolis ↓ serum levels of TNF-α and IL-6. Propolis ↓ TGF-β activation and Smad phosphorylation. | [91] |
| Rabdosia amethystoides (Root, Lamiaceae) | EtOH extract (ERA) | ERA (50 mg/kg) ERA (100 mg/kg) ERA (150 mg/kg) Con A (15 mg/mL/kg) | ERA significantly ↓ the elevated ALT and AST levels and liver necrosis in Con A-induced hepatitis. ERA significantly ↓ MPO and MDA levels and ↑ SOD level in the liver tissue. ERA significantly ↓ the expression level of TLR4 mRNA or protein in liver tissues. ERA attenuated the activation of the NF-kB pathway by inhibiting IkBα kinase and p65 phosphorylation. ERA effect may be mediated by the downregulation of TLR4 expression and inhibition of NF-kB activation. | [92] |
| Zea mays (Seeds, Poaceae) | Selenium-bio-fortified Peptides (SeCPs) | SeCPs (200 mg/kg) Con A (15 mg/kg) | SeCPs significantly ↓ AST, ALT, TNF-α, IFN-γ contents in serum, and MDA contents in liver. SeCPs significantly ↑ SOD and GPx activities. SeCPs effect was related to its antioxidant capacity, reduction of lipid peroxidation, and inhibiting the release of immune factors (TNF-α and IFN-γ) | [93] |
| Tetrastigma hemsleyanum (Roots, Vitaceae) | Total flavonoids (TFT) | TFT (1 mg/kg) TFT (2 mg/kg) TFT (4 mg/kg) Con A (20 mg/kg) | TFT significantly ↓ ALT and AST serum levels and attenuated histopathological alterations in Con A-induced liver injury. TFT ↓ increased serum levels of IL-17 and IL-6, Th17 cells proportions in spleen and the expression of RORγt in hepatic tissues. TFT enhanced % of Treg cells in spleen, the expression of Foxp3 in hepatic tissues and levels TGF-β1 and IL-10 in serum. | [94] |
| Tupistra chinensis (Rhizomes, Asparagaceae) | EtOH extract (TCE) (saponins) | TCE (75 mg/kg) TCE (150 mg/kg) TCE (300 mg/kg) Con A (15 mg/kg) | TCE significantly ↓ ALT, AST, and LDH serum levels. TCE ↓ IFN-γ and TNF-α levels. TCE suppressed MAPK and NF-κB-signaling in liver. TCE blocked STAT1/NF-κB-signaling and induced apoptosis in activated T cells. | [95] |
| Yiguanjian decoction (YD) Glehnia littoralis (Roots, Apiaceae) Ophiopogon japonicus (Roots, Asparagaceae) Angelica sinensis (Roots, Apiaceae) Rehmannia glutinosa (Roots, Scrophulariaceae) Lycium barbarum (Fruits, Solanaceae) Melia toosendan (Fruits, Meliaceae) | Aqueous extract | YD (0.13 g/mL) YD (0.26 g/mL) YD (0.52 g/mL) Con A (2 mg/mL) | YD improved the degree of liver inflammation and fibrosis in the liver of chronic hepatitis mice. YD ↓ liver cell DNA damage. YD ↓ protein expression of TNF-α in liver tissue. YD ↑ mRNA expression of Bax and MTH1. | [96] |
| Jiang-Xian HuGan (JXHG) herbal formula Corbicula fluminea (Clams, Cyrenidae) Curcuma longa (Rhizomes, Zingiberaceae) Ligustrum lucidum (Leaves, Oleaceae) Eclipta prostrata (Herb, Asteraceae) Paeonia lactiflora (Roots, Paeoniaceae) | EtOH extract | JXHG (700 mg/kg) JXHG (1400 mg/kg) Con A (20 mg/kg) | JXHG ↓ serum levels of ALT and AST, and ↓ hepatocyte apoptosis and mortality. JXHG significantly ↓ the serum levels of IFN-γ, TNF-α, IL-4, and IL-6. JXHG ↓ the mRNA expression of IL-6 and IFN-γ and ↑ IL-10, SOD1, GSR, and GPX2 mRNA in the liver tissues. JXHG dramatically suppressed NF-κB p65 phosphorylation, ↑ Nrf2 expression, and ↓ expression ratio of cleaved caspase-3/caspase-3 in liver tissues. | [97] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibrahim, S.R.M.; Sirwi, A.; Eid, B.G.; Mohamed, S.G.A.; Mohamed, G.A. Summary of Natural Products Ameliorate Concanavalin A-Induced Liver Injury: Structures, Sources, Pharmacological Effects, and Mechanisms of Action. Plants 2021, 10, 228. https://doi.org/10.3390/plants10020228
Ibrahim SRM, Sirwi A, Eid BG, Mohamed SGA, Mohamed GA. Summary of Natural Products Ameliorate Concanavalin A-Induced Liver Injury: Structures, Sources, Pharmacological Effects, and Mechanisms of Action. Plants. 2021; 10(2):228. https://doi.org/10.3390/plants10020228
Chicago/Turabian StyleIbrahim, Sabrin R. M., Alaa Sirwi, Basma G. Eid, Shaimaa G. A. Mohamed, and Gamal A. Mohamed. 2021. "Summary of Natural Products Ameliorate Concanavalin A-Induced Liver Injury: Structures, Sources, Pharmacological Effects, and Mechanisms of Action" Plants 10, no. 2: 228. https://doi.org/10.3390/plants10020228
APA StyleIbrahim, S. R. M., Sirwi, A., Eid, B. G., Mohamed, S. G. A., & Mohamed, G. A. (2021). Summary of Natural Products Ameliorate Concanavalin A-Induced Liver Injury: Structures, Sources, Pharmacological Effects, and Mechanisms of Action. Plants, 10(2), 228. https://doi.org/10.3390/plants10020228