
Weather Variables Associated with Spore Dispersal of Lecanosticta acicola Causing Pine Needle Blight in Northern Spain

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Spore Traps Location
2.2. Design and Measurements of Spore Traps
2.3. Meteorological Data
2.4. Statistical Analysis
3. Results
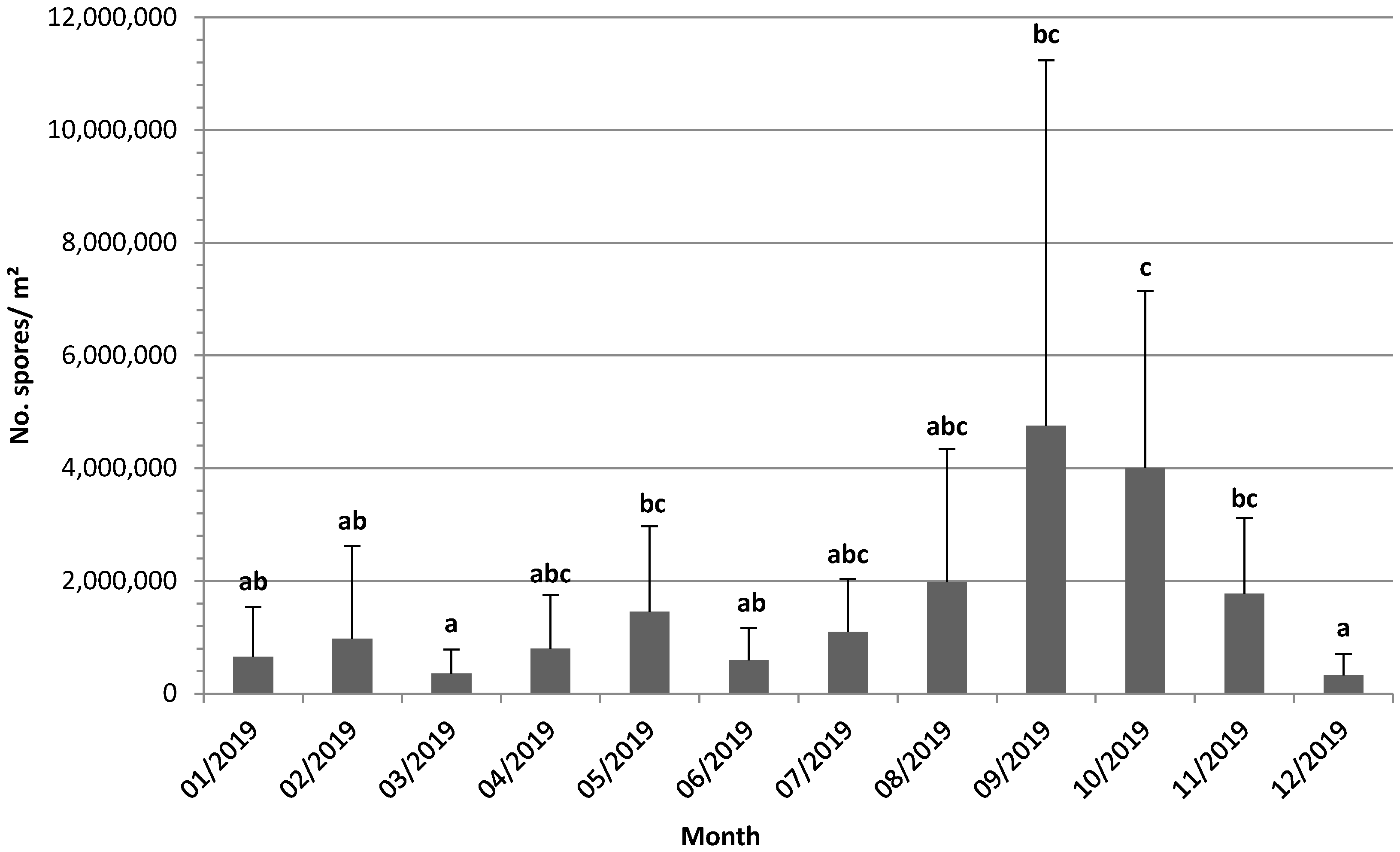
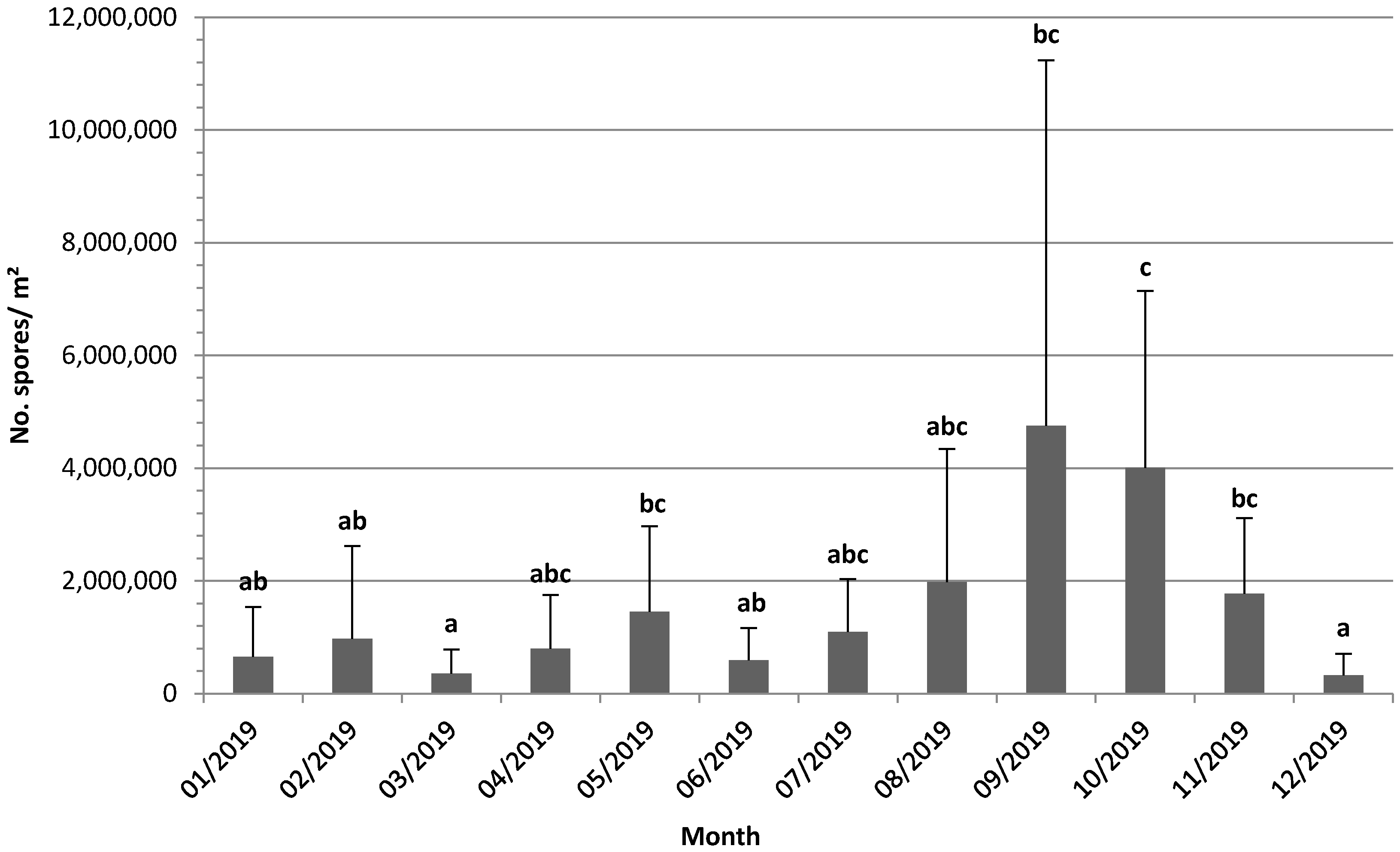
3.1. Measures of Spore Dispersal
3.2. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ortíz de Urbina, E.; Mesanza, N.; Aragonés, A.; Raposo, R.; Elvira-Recuenco, M.; Boqué, R.; Patten, C.; Aitken, J.; Iturritxa, E. Emerging needle blight diseases in Atlantic pinus ecosystems of Spain. Forests 2017, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Van der Nest, A.; Wingfield, M.J.; Janoušek, J.; Barnes, I. Lecanosticta acicola: A growing threat to expanding global pine forests and plantations. Mol. Plant Pathol. 2019, 20, 1327–1364. [Google Scholar] [CrossRef] [Green Version]
- Broders, K.; Munck, I.A.; Wyka, S.A.; Iriarte, G.; Beaudoin, E. Characterization of fungal pathogens associated with white pine needle damage (WPND) in northeastern North America. Forests 2015, 6, 4088–4104. [Google Scholar] [CrossRef]
- Fitt, B.D.L.; McCartney, H.A.; Walklate, P.J. The role of rain in dispersal of pathogen inoculum. Annu. Rev. Phytopathol. 1989, 27, 241–270. [Google Scholar] [CrossRef]
- Wyka, S.A.; Smith, C.; Munck, I.A.; Rock, B.N.; Ziniti, B.L.; Broders, K. Emergence of white pine needle damage in the northeastern United States is associated with changes in pathogen pressure in response to climate change. Glob. Chang. Biol. 2017, 23, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Wyka, S.A.; McIntire, C.D.; Smith, C.; Munck, I.A.; Rock, B.N.; Asbjornsen, H.; Broders, K.D. Effect of climatic variables on abundance and dispersal of Lecanosticta acicola spores and their impact on defoliation on eastern white pine. Phytopathology 2018, 108, 374–383. [Google Scholar] [CrossRef] [Green Version]
- Suto, Y. Seasonal development of symptoms and conidial production and dispersal of Lecanosticta acicola in Pinus thunbergii. Appl. For. Sci. 2002, 11, 17–22. [Google Scholar]
- Skilling, D.D.; Nicholls, T.H. Brown spot needle disease—Biology and control in Scotch pine plantations. USDA For. Serv. Res. Pap. 1974, NC-109, 1–19. [Google Scholar]
- Sadiković, D.; Piškur, B.; Barnes, I.; Hauptman, T.; Diminić, D.; Wingfield, M.J.; Jurc, D. Genetic diversity of the pine pathogen Lecanosticta acicola in Slovenia and Croatia. Plant Pathol. 2019, 68, 1120–1131. [Google Scholar] [CrossRef]
- Siggers, P.V. The brown spot needle blight of pine seedlings. USDA Tech. Bull. 1944, 870, 1–36. [Google Scholar]
- Janoušek, J.; Wingfield, M.J.; Monsivais, J.G.; Jankovský, L.; Stauffer, C.; Konečný, A.; Barnes, I. Genetic analyses suggest separate introductions of the pine pathogen Lecanosticta acicola into Europe. Phytopathology 2016, 106, 1413–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesanza, N.; Hernández, M.; Raposo, R.; Iturritxa, E. First report of Mycosphaerella dearnessii Rostrup, teleomorph of Lecanosticta acicola (Thüm.) Syd., in Europe. Plant Health Prog. 2021, 22, 565–566. [Google Scholar] [CrossRef]
- EPPO. PM 7/46 (3) Lecanosticta acicola (formerly Mycosphaerella dearnessii), Dothistroma septosporum (formerly Mycosphaerella pini) and Dothistroma pini. EPPO Bull. 2015, 45, 163–182. [Google Scholar] [CrossRef]
- Sinclair, W.A.; Lyon, H.H.; Johnson, W.T. Diseases of Trees and Shrubs; Cornell University Press: Ithaca, NY, USA, 1987. [Google Scholar]
- Cordell, C.E.; Gramling, C.; Lowman, B.; Brown, D. A precision seed sower for longleaf pine bareroot nursery seedlings. Tree Plant. Notes 1990, 41, 33–38. [Google Scholar]
- Tainter, F.H.; Baker, F.A. Brown spot. In Principles of Forest Pathology; John Wiley: New York, NY, USA, 1996; pp. 467–492. [Google Scholar]
- McIntire, C.D.; Munck, I.A.; Ducey, M.J.; Asbjornsen, H. Thinning treatments reduce severity of foliar pathogens in eastern white pine. For. Ecol. Manag. 2018, 423, 106–113. [Google Scholar] [CrossRef]
- Michel, A.; Seidling, W. Forest condition in Europe: 2016 technical report of ICP forests. In UNECE Convention on Long-Range Transboundary air Pollution (CLRTAP); BFW Dokumentation 23/2016; BFWAustrian Research Centre for Forests: Vienna, Austria, 2016; p. 206. [Google Scholar]
- Iturritxa, E.; Ganley, R. Dispersión por vía aérea de esporas de Diplodia pinea en tres localidades de la cornisa cantábrica. Bol. San. Veg. Plagas 2007, 33, 383–390. [Google Scholar]
- Meuten, D.J.; Moore, F.M.; George, J.W. Mitotic Count and the Field of View Area: Time to Standardize. Vet. Pathol. 2016, 53, 7–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruskal, W.H.; Wallis, W.A. Use of ranks in one-criterion variance analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- IBM Corp. Released 2020; IBM SPSS Statistics for Windows, Version 27.0; IBM Corp.: Armonk, NY, USA, 2020. [Google Scholar]
- Euskalmet. Available online: https://opendata.euskadi.eus/catalogo/-/estaciones-meteorologicas-lecturas-recogidas-en-2019/ (accessed on 17 August 2021).
- R Core Team. R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. 2017. Available online: https://www.R-project.org/ (accessed on 20 June 2021).
- Wood, S.N. Generalized Additive Models: An Introduction with R; Chapman and Hall/CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Multimodel inference: Understanding AIC and BIC in model selection. Sociol. Methods Res. 2004, 33, 261–304. [Google Scholar] [CrossRef]
- Dvorak, M.; Jankovsky, L.; Drapela, K. Dothistroma septosporum: Spore production and weather conditions. For. Syst. 2012, 21, 323–328. [Google Scholar] [CrossRef] [Green Version]
- Kais, A.G. Variation between southern and northern isolates of Scirrhia acicola. Phytopathology 1972, 62, 768. [Google Scholar]
- Ramos, P.; Petisco, E.; Martín, J.M.; Rodríguez, E. Downscaled climate change projections over Spain: Application to water resources. Internat. J. Water Resour. Dev. 2012, 29, 201–218. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Trap ID | Province | X Coordinates | Y Coordinates | Orientation | Slope (%) | Age | Defoliation Level of Site (%) |
|---|---|---|---|---|---|---|---|
| Albina | Araba | 531,468 | 4,762,368 | Southeast | 5 to 10 | 13 | 25 |
| Oleta | Araba | 531,448 | 4,765,973 | Southwest | 20 to 30 | 9 | 30 |
| Idiazabal Larraegi | Gipuzkoa | 563,439 | 4,760,042 | Southwest | 30 to 50 | 4 | 30 |
| Azpeitia Igarate | Gipuzkoa | 557,840 | 4,777,305 | Northwest | 30 to 50 | 9 | 70 |
| Mallabia | Bizkaia | 535,952 | 4,785,234 | Northeast | 20 to 30 | <15 | >30 |
| Muxika | Bizkaia | 523,110 | 4,787,806 | Northeast | 30 to 50 | <15 | >30 |
| Igorre | Bizkaia | 516,044 | 4,780,811 | South | 30 to 50 | <15 | >30 |
| Güeñes | Bizkaia | 493,228 | 4,783,097 | Northeast | 50 to 100 | <15 | >30 |
| Karrantza | Bizkaia | 475,875 | 4,785,737 | West | 10 to 20 | <15 | >30 |
| Elorrio | Bizkaia | 539,801 | 4,778,037 | Northwest | 10 to 20 | 4 | 50 |
| Pagatza | Gipuzkoa | 540,597 | 4,776,579 | North | 10 to 20 | 12 | 55 |
| Lezama1 | Bizkaia | 515,746 | 4,793,192 | South | 20 to 30 | 5 | 55 |
| Lezama2 | Bizkaia | 515,746 | 4,793,192 | South | 20 to 30 | 5 | 55 |
| Umbe1 | Bizkaia | 506,024 | 4,799,627 | North | 5 to 10 | 14 | 50 |
| Umbe2 | Bizkaia | 506,024 | 4,799,627 | North | 5 to 10 | 14 | 50 |
| Idiazabal | Azpeitia | Karrantza | Güeñes | Igorre | Muxika | Mallabia | Unbe1 | Unbe2 | Lezama1 | Lezama2 | Elorrio | Pagatza | Olaeta | Albina | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 07/01/2019 | ND | ND | ND | ND | ND | ND | ND | 50,295 | 6035 | ND | ND | 18,106 | 42,247 | 4024 | 2012 |
| 21/01/2019 | ND | ND | ND | ND | ND | ND | ND | 116,683 | 54,318 | 261,532 | 74,436 | 114,672 | 86,507 | 21,906 | 0 |
| 04/02/2019 | ND | ND | 0 | 0 | 18,505 | 14,235 | 39,858 | 26,746 | 10,698 | 77,564 | 40,119 | 18,722 | 2675 | 21,906 | 0 |
| 18/02/2019 | 6404 | 6404 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 35,524 | 0 | 0 | 582,592 | 19,824 | 88,105 |
| 04/03/2019 | 0 | 0 | 2496 | 0 | 17,474 | 7489 | 24,963 | 28,419 | 7105 | 10,657 | 17,762 | 28,419 | 3552 | 7105 | 0 |
| 18/03/2019 | 1949 | 1949 | 23,793 | 1322 | 7931 | 10,574 | 15,862 | 17,762 | 7105 | 0 | 0 | 0 | 0 | 0 | 0 |
| 01/04/2019 | 8966 | 0 | 7931 | 1322 | 5287 | 6609 | 6609 | 202,020 | 15,151 | 75,757 | 35,354 | 10,101 | 0 | 0 | 0 |
| 15/04/2019 | 0 | 25,617 | 52,459 | 34,973 | 31,475 | 13,989 | 13,989 | 202,020 | 15,152 | 75,758 | 35,354 | 10,101 | 0 | 0 | 102,973 |
| 29/04/2019 | 25,617 | 6404 | 3264 | 3264 | 11,424 | 8160 | 8160 | 9946 | 2486 | 134,280 | 14,920 | 37,300 | 12,433 | 12,433 | 7460 |
| 13/05/2019 | 2989 | 13,185 | 5649 | 0 | 7533 | 5649 | 1883 | 31,971 | 3552 | 142,096 | 120,781 | 284,191 | 28,419 | 0 | 0 |
| 27/05/2019 | 44,830 | 110,036 | 22,732 | 3497 | 110,164 | 66,448 | 57,705 | 14,210 | 63,943 | 92,362 | 81,705 | 138,543 | 56,838 | 31,971 | 0 |
| 10/06/2019 | 50,104 | 5274 | 5649 | 0 | 28,247 | 5649 | 7533 | 29,840 | 6631 | 46,418 | 72,942 | 66,311 | 0 | 46,181 | 0 |
| 24/06/2019 | 16,302 | 0 | 0 | 0 | 36,721 | 40,219 | 0 | 0 | 11,477 | 3826 | 7651. | 0 | 0 | 92,362 | 3552 |
| 08/07/2019 | 0 | 0 | 4080 | 0 | 134,645 | 96,564 | 28,561 | 0 | 7105 | 39,076 | 85,257 | 3552 | 49,733 | 23,272 | 18,864 |
| 22/07/2019 | 3202 | 0 | 4080 | 0 | 134,645 | 96,564 | 28,561 | 53,049 | 33,156 | 102,782 | 62,996 | 62,996 | 43,102 | 23,272 | 18,864 |
| 05/08/2019 | 3202 | 0 | 0 | 0 | 54,837 | 75,401 | 67,567 | 9947 | 4973 | 39,787 | 34,813 | 74,600 | 0 | 169,094 | 29,840 |
| 19/08/2019 | 0 | 0 | 0 | 0 | 54,837 | 75,401 | 67,567 | 46,807 | 4973 | 32,180 | 1755 | 32,180 | 1170 | 2925 | 2925 |
| 02/09/2019 | 10,345 | 0 | 1749 | 0 | 117,159 | 138,142 | 96,175 | 144,679 | 226,061 | 1,446,791 | 578,716 | 149,200 | 9042 | 60,391 | 10,657 |
| 16/09/2019 | 0 | 0 | 0 | 0 | 132,896 | 78,689 | 36,721 | 418,346 | 198,934 | 854,245 | 424,197 | 359,836 | 17,553 | 7105 | 0 |
| 30/09/2019 | 0 | 0 | 0 | 0 | 342,733 | 35,780 | 11,299 | 294,848 | 63,943 | 291,296 | 209,591 | 191,829 | 60,391 | 177,619 | 10,657 |
| 14/10/2019 | 35,864 | 65,750 | 0 | 0 | 172,998 | 127,301 | 200,744 | 195,381 | 39,076 | 255,772 | 209,591 | 269,982 | 14,210 | 134,991 | 40,260 |
| 28/10/2019 | 32,021 | 64,754 | 0 | 1748 | 125,902 | 253,552 | 103,169 | 504,439 | 195,381 | 319,715 | 298,401 | 127,886 | 14,210 | 134,991 | 40,260 |
| 11/11/2019 | 22,415 | 84,678 | 0 | 0 | 131,820 | 86,625 | 122,405 | 165,778 | 175,725 | 62,996 | 16,578 | 179,040 | 3316 | 71,048 | 60,391 |
| 25/11/2019 | 6897 | 32,875 | 1632 | 0 | 29,377 | 44,066 | 26,113 | 0 | 0 | 95,914 | 49,733 | 110,124 | 7105 | 29,840 | 6631 |
| 09/12/2019 | 0 | 4981 | 0 | 0 | 5246 | 13,989 | 1749 | 8913 | 0 | 41,592 | 77,243 | 8913 | 0 | 17,825 | 0 |
| 23/12/2019 | 4483 | 16,302 | 0 | 1632 | 27,745 | 8160 | 37,537 | 17,361 | 0 | 14,205 | 6313 | 17,361 | 1578 | 2185 | 0 |
| Data from All Traps | Leaving out Pagatza and Lezama 1 | |||
|---|---|---|---|---|
| Variable | Coefficient | Std. Error | Coefficient | Std. Error |
| Daily maximum temperature * | 78,002 | 38,497 | 83,075 | 27,376 |
| Daily cumulative precipitation * | 47,580 | 26,445 | 44,785 | 19,572 |
| Daily maximum relative humidity | 12,280 | 19,662 | 10,527 | 13,340 |
| Daily mean irradiance | −2106 | 2386 | −1811 | 1650 |
| Daily average wind speed | −41,958 | 100,710 | −11,860 | 74,291 |
| Model | Temp | Rainfull | Humidity | Irrad | Wind | k | Δi |
|---|---|---|---|---|---|---|---|
| 1 | Daily maximum * | Cumulative precipitation * | Daily maximum | Daily mean | Average speed | 8 | 0 |
| 2 | Daily maximum * | Cumulative precipitation * | Daily mean | Daily mean | Average speed | 8 | 0.683 |
| 3 | Daily maximum * | Cumulative precipitation * | Daily maximum | Daily maximum | Average speed | 8 | 0.725 |
| 4 | Daily maximum * | Cumulative precipitation * | Daily maximum | Daily mean | Average speed | 10 | 1.163 |
| 5 | Daily maximum * | Cumulative precipitation * | Daily maximum | Daily mean | Average speed | 12 | 1.373 |
| 6 | Daily maximum * | Cumulative precipitation * | Daily mean | Daily maximum | Average speed | 8 | 1.41 |
| 7 | Daily maximum * | Cumulative precipitation * | Daily mean | Daily mean | Average speed | 10 | 1.795 |
| 8 | Daily maximum * | Cumulative precipitation * | Daily maximum | Daily maximum | Average speed | 10 | 1.897 |
| Final Model | Deviance Explained = 41.5% | k = 8 Basis Functions |
|---|---|---|
| Variable | Coefficient | Std. error |
| Daily maximum temperature * | 77,652 | 26,153 |
| Cumulative precipitation * | 50,438 | 18,987 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mesanza, N.; García-García, D.; Raposo, E.R.; Raposo, R.; Iturbide, M.; Pascual, M.T.; Barrena, I.; Urkola, A.; Berano, N.; Sáez de Zerain, A.; et al. Weather Variables Associated with Spore Dispersal of Lecanosticta acicola Causing Pine Needle Blight in Northern Spain. Plants 2021, 10, 2788. https://doi.org/10.3390/plants10122788
Mesanza N, García-García D, Raposo ER, Raposo R, Iturbide M, Pascual MT, Barrena I, Urkola A, Berano N, Sáez de Zerain A, et al. Weather Variables Associated with Spore Dispersal of Lecanosticta acicola Causing Pine Needle Blight in Northern Spain. Plants. 2021; 10(12):2788. https://doi.org/10.3390/plants10122788
Chicago/Turabian StyleMesanza, Nebai, David García-García, Elena R. Raposo, Rosa Raposo, Maialen Iturbide, Mª Teresa Pascual, Iskander Barrena, Amaia Urkola, Nagore Berano, Aitor Sáez de Zerain, and et al. 2021. "Weather Variables Associated with Spore Dispersal of Lecanosticta acicola Causing Pine Needle Blight in Northern Spain" Plants 10, no. 12: 2788. https://doi.org/10.3390/plants10122788
APA StyleMesanza, N., García-García, D., Raposo, E. R., Raposo, R., Iturbide, M., Pascual, M. T., Barrena, I., Urkola, A., Berano, N., Sáez de Zerain, A., & Iturritxa, E. (2021). Weather Variables Associated with Spore Dispersal of Lecanosticta acicola Causing Pine Needle Blight in Northern Spain. Plants, 10(12), 2788. https://doi.org/10.3390/plants10122788