
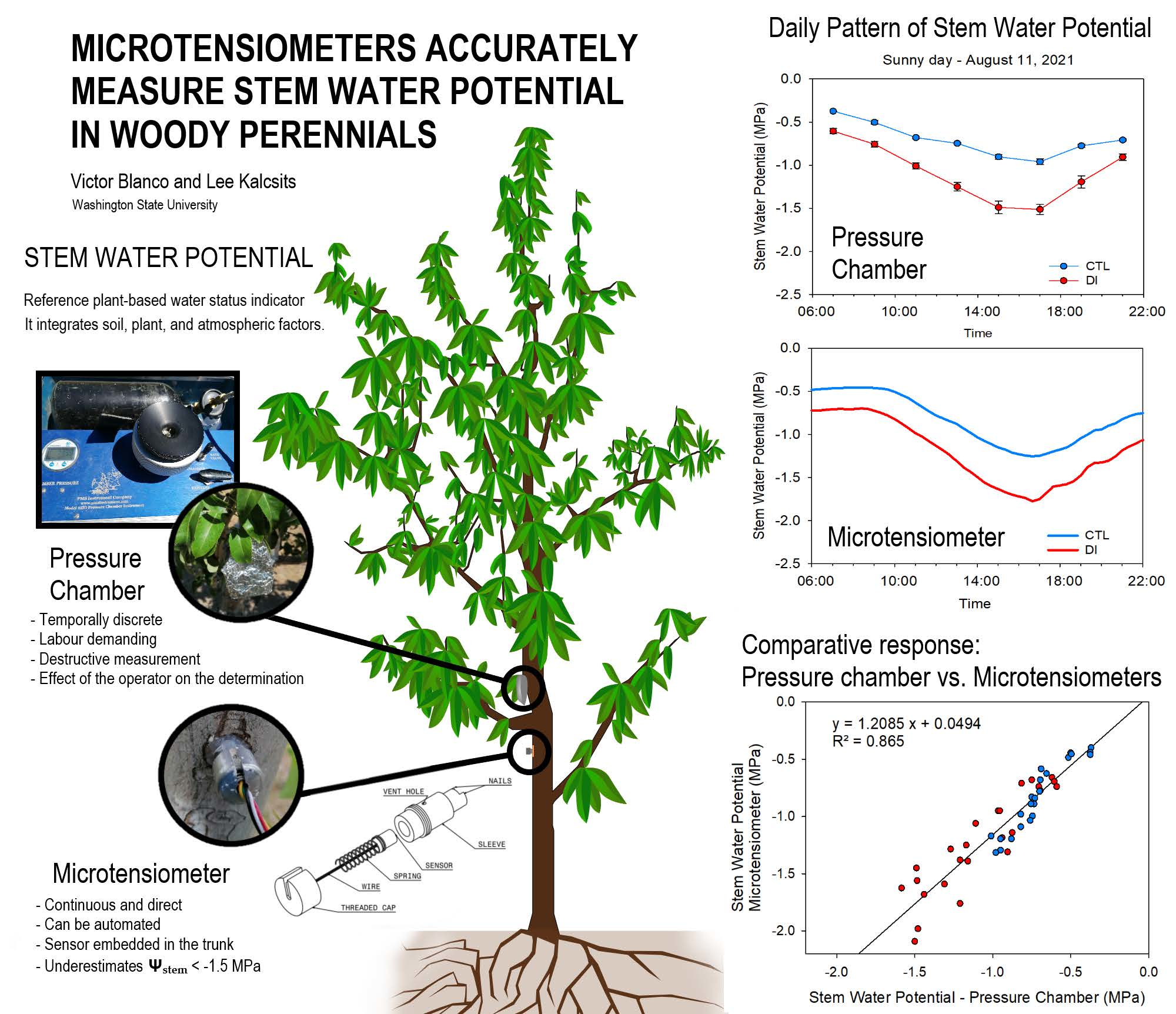
Microtensiometers Accurately Measure Stem Water Potential in Woody Perennials


Abstract
:
1. Introduction
2. Results
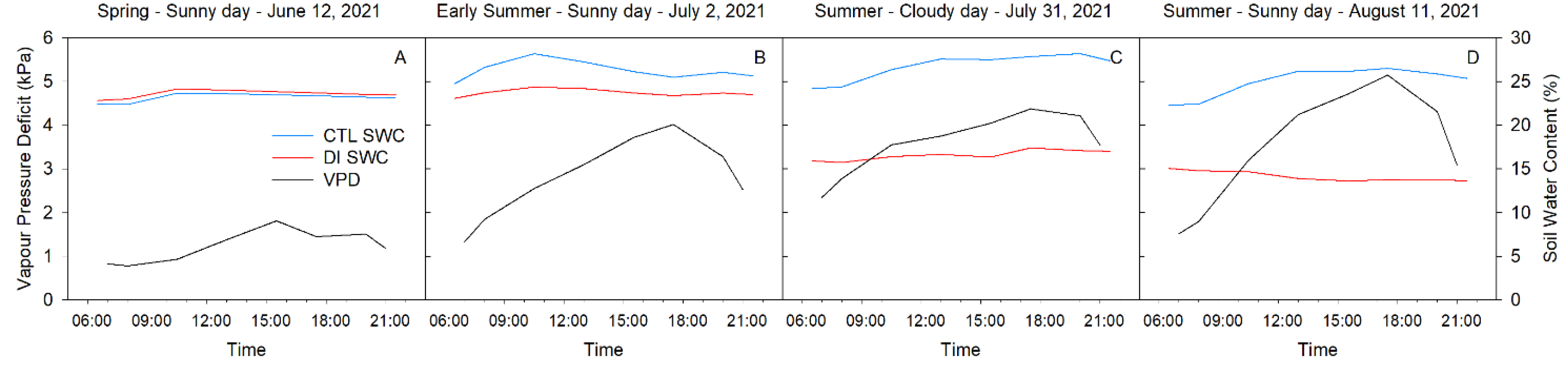
2.1. Weather and Soil Conditions
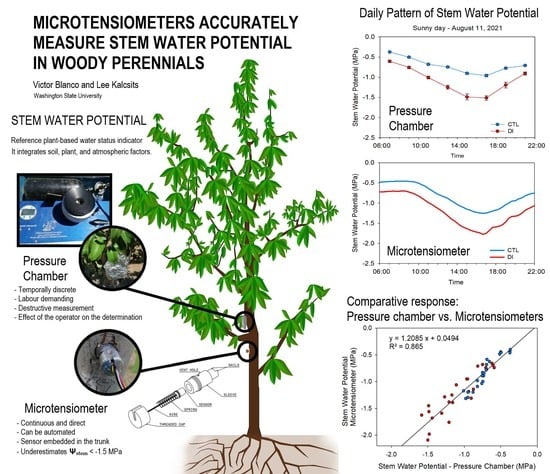
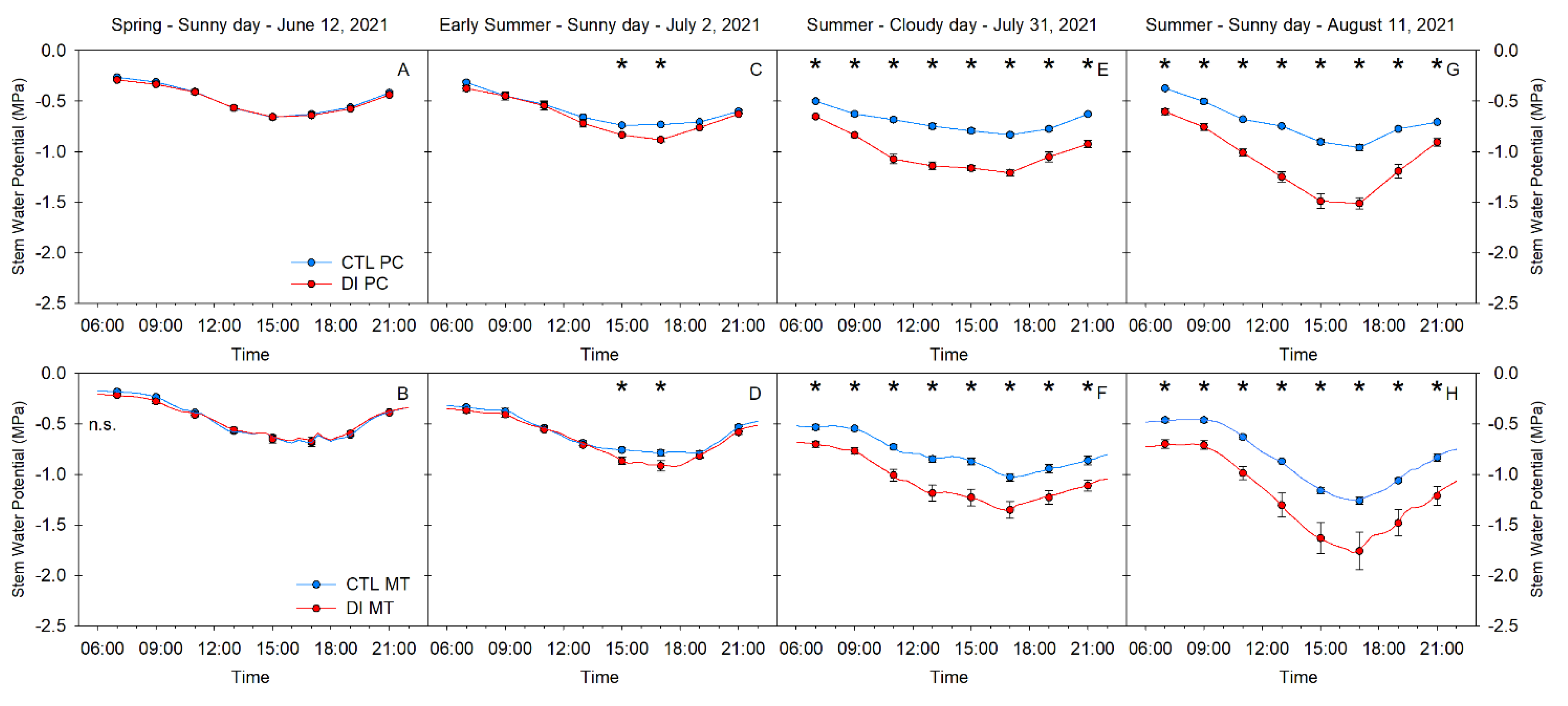
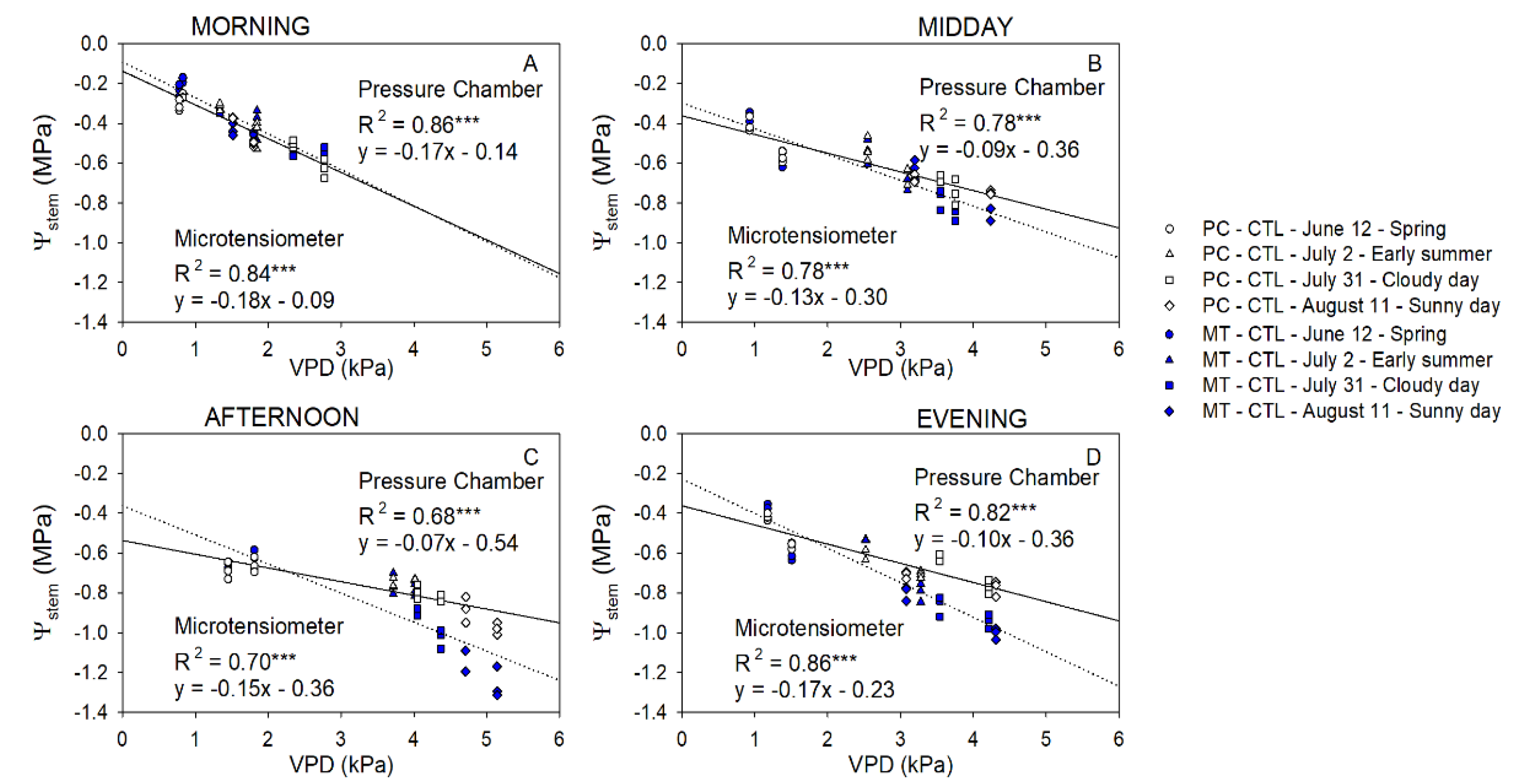
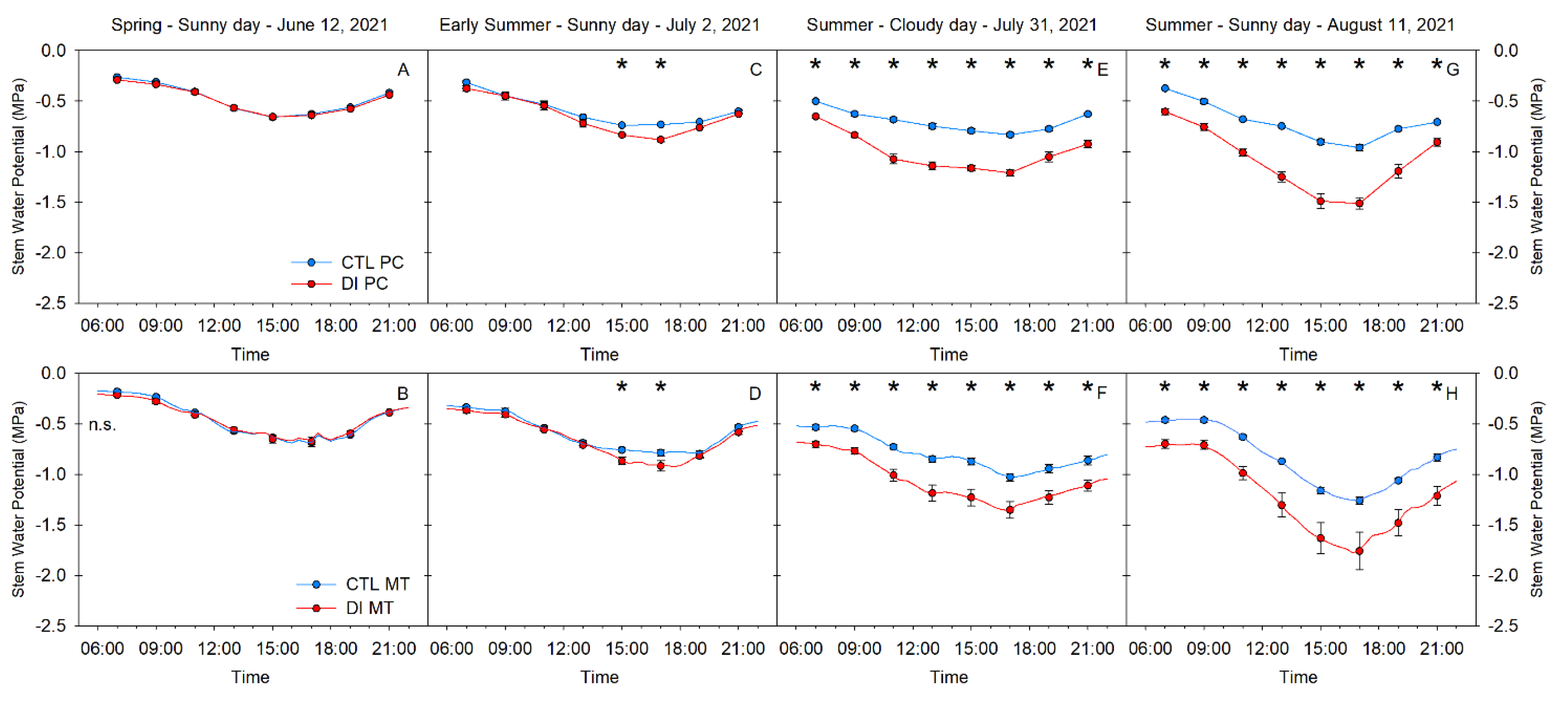
2.2. Stem Water Potential. Diurnal Pattern
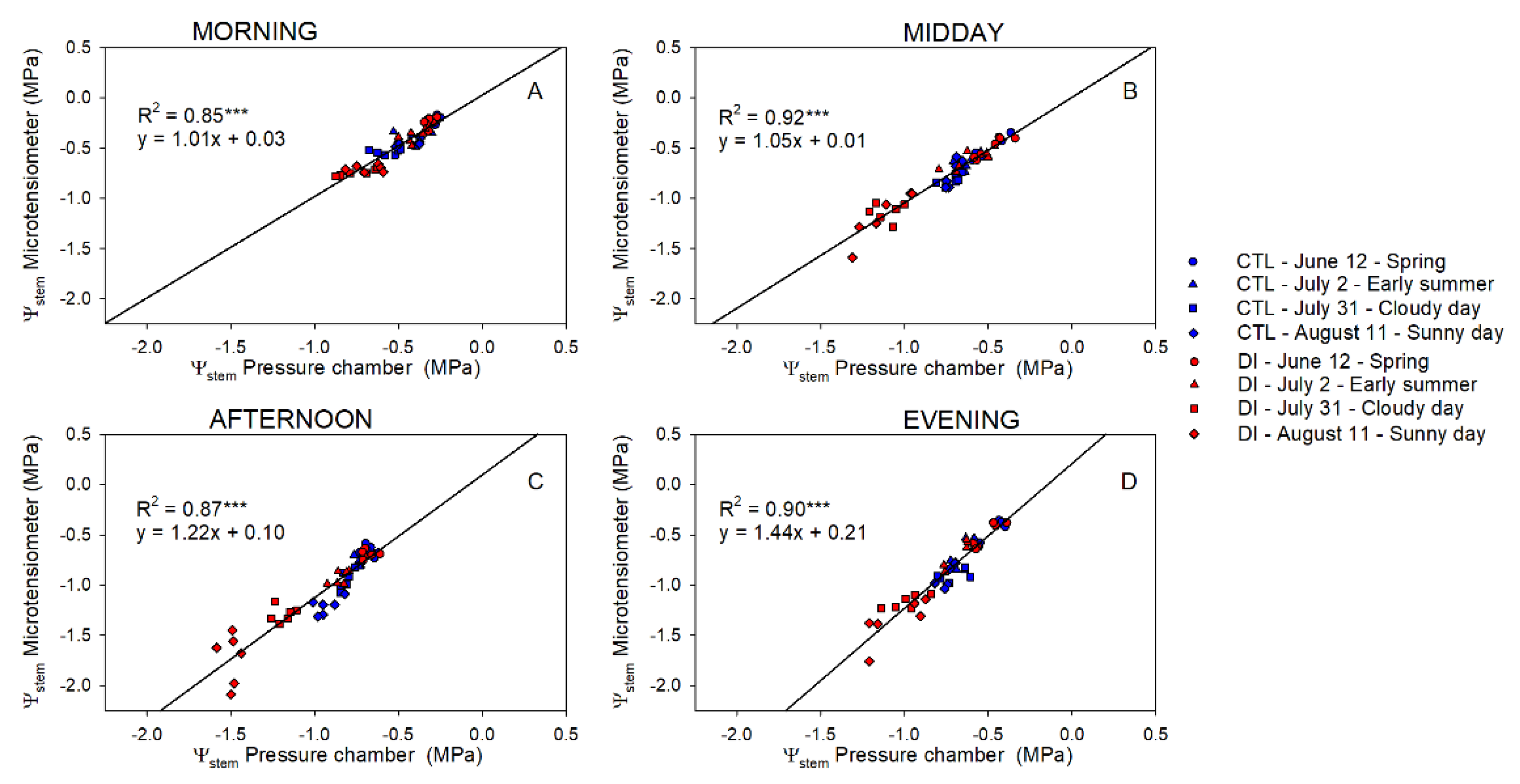
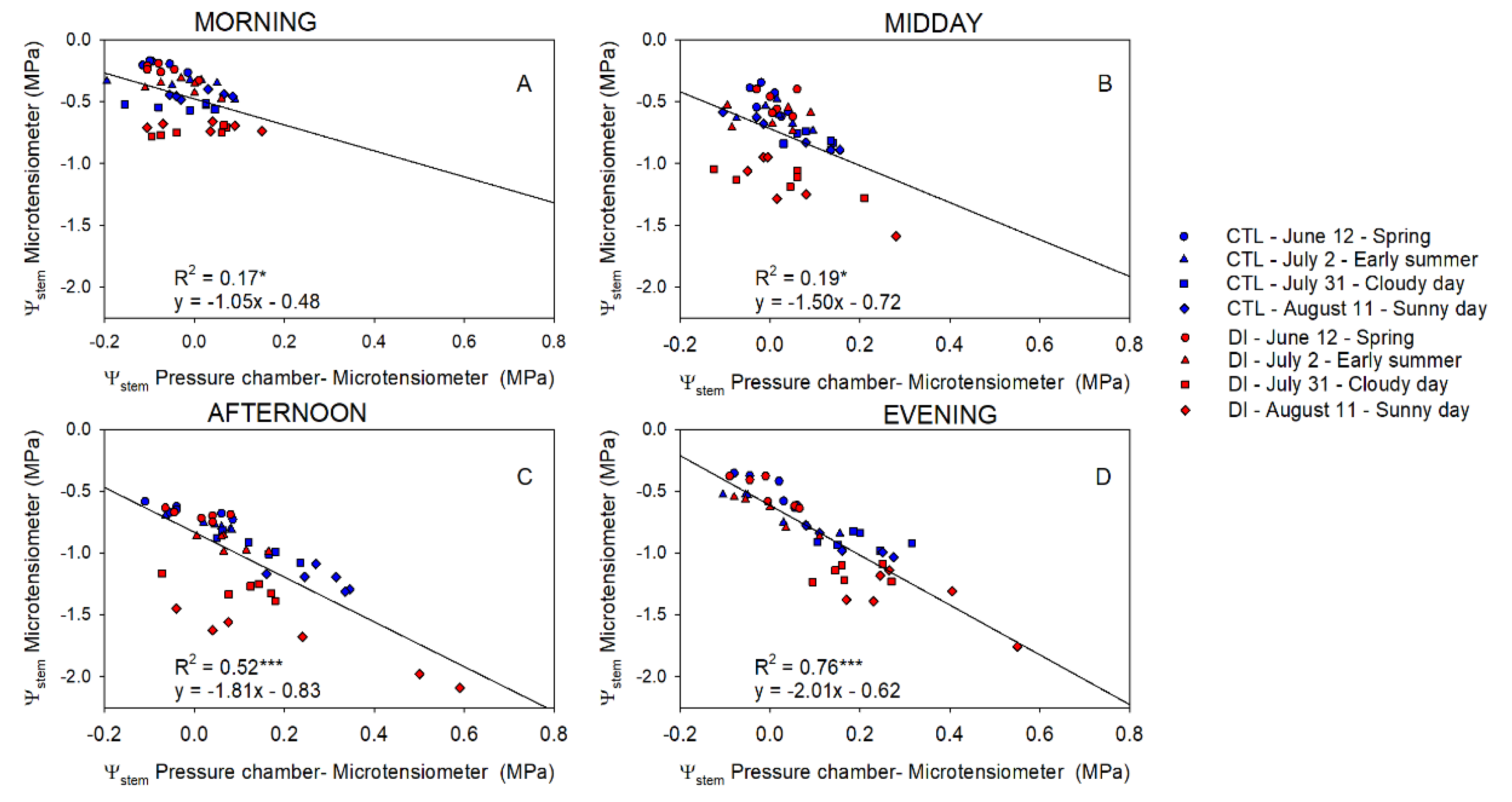
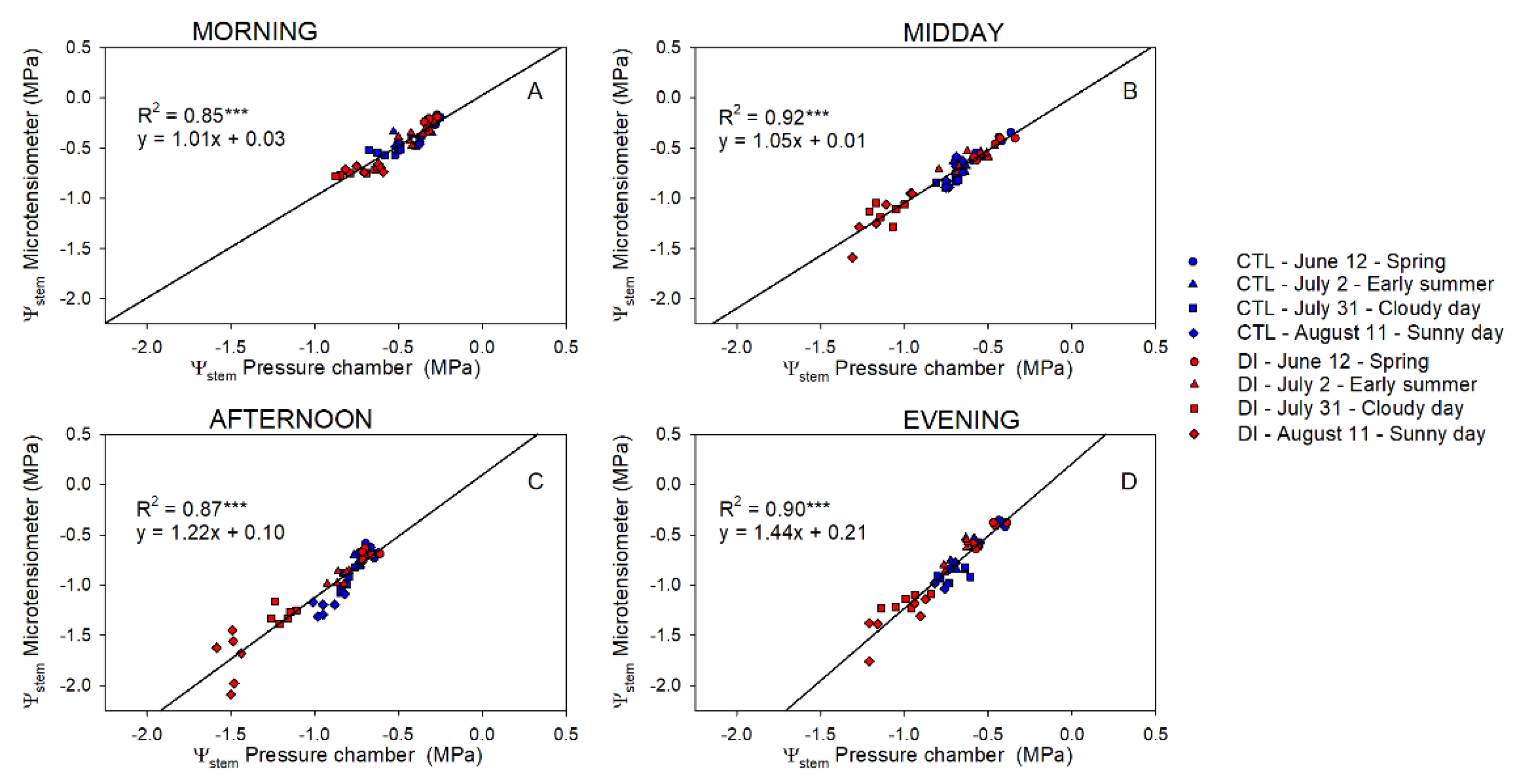
2.3. Stem Water Potential. Comparative Response: Pressure Chamber vs. Microtensiometers
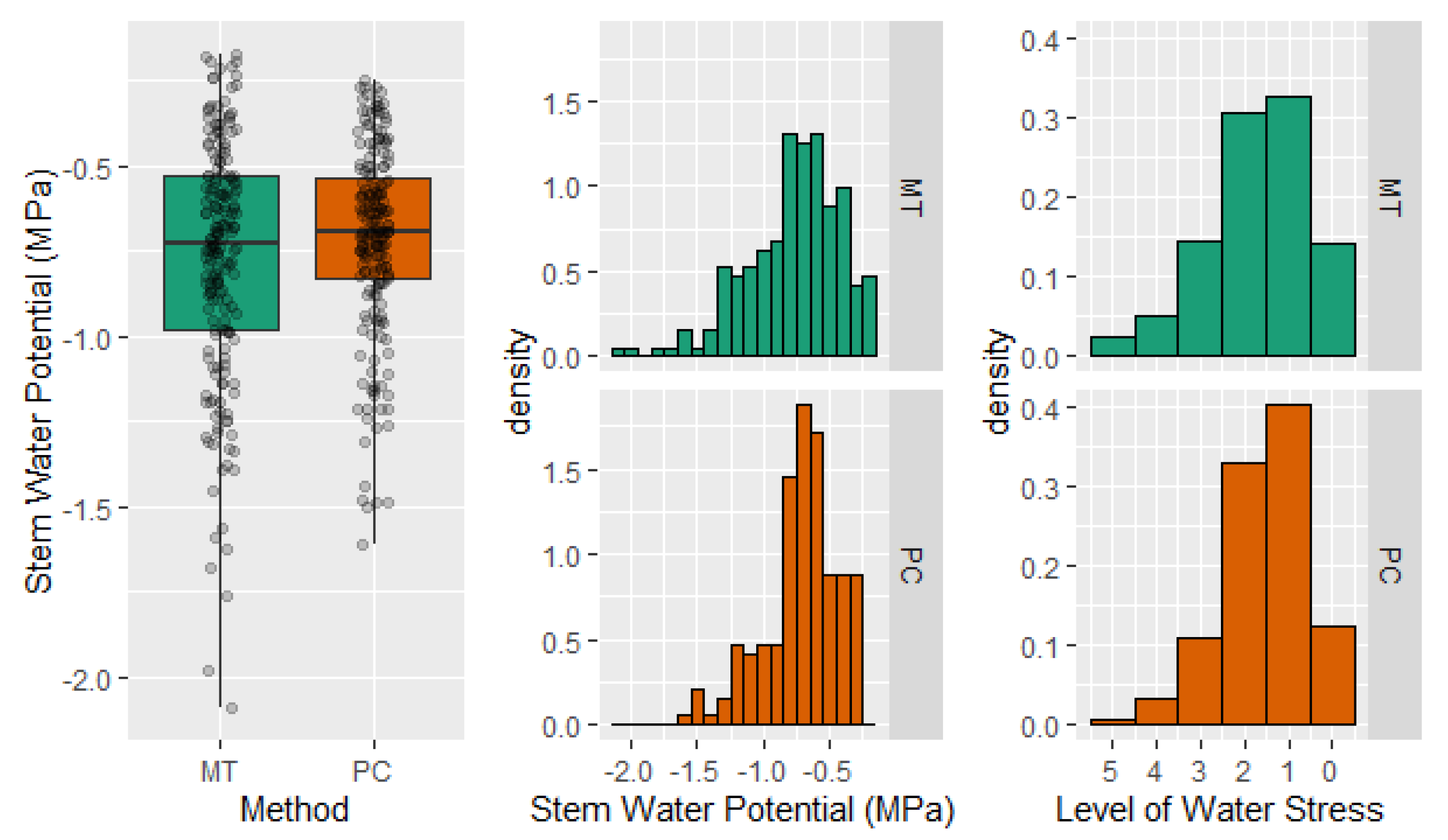
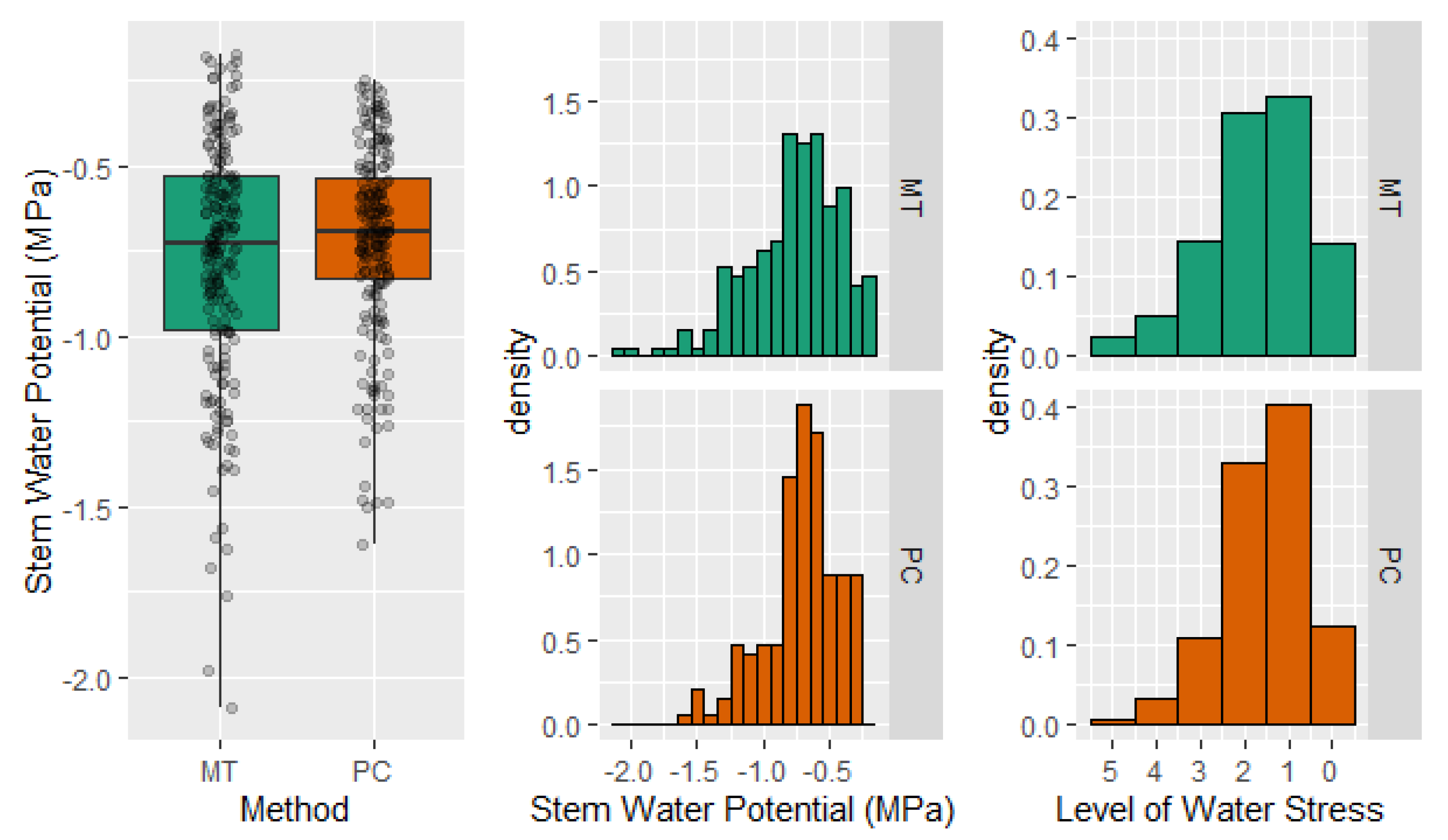
2.4. Stem Water Potential. Level of Tree Water Stress
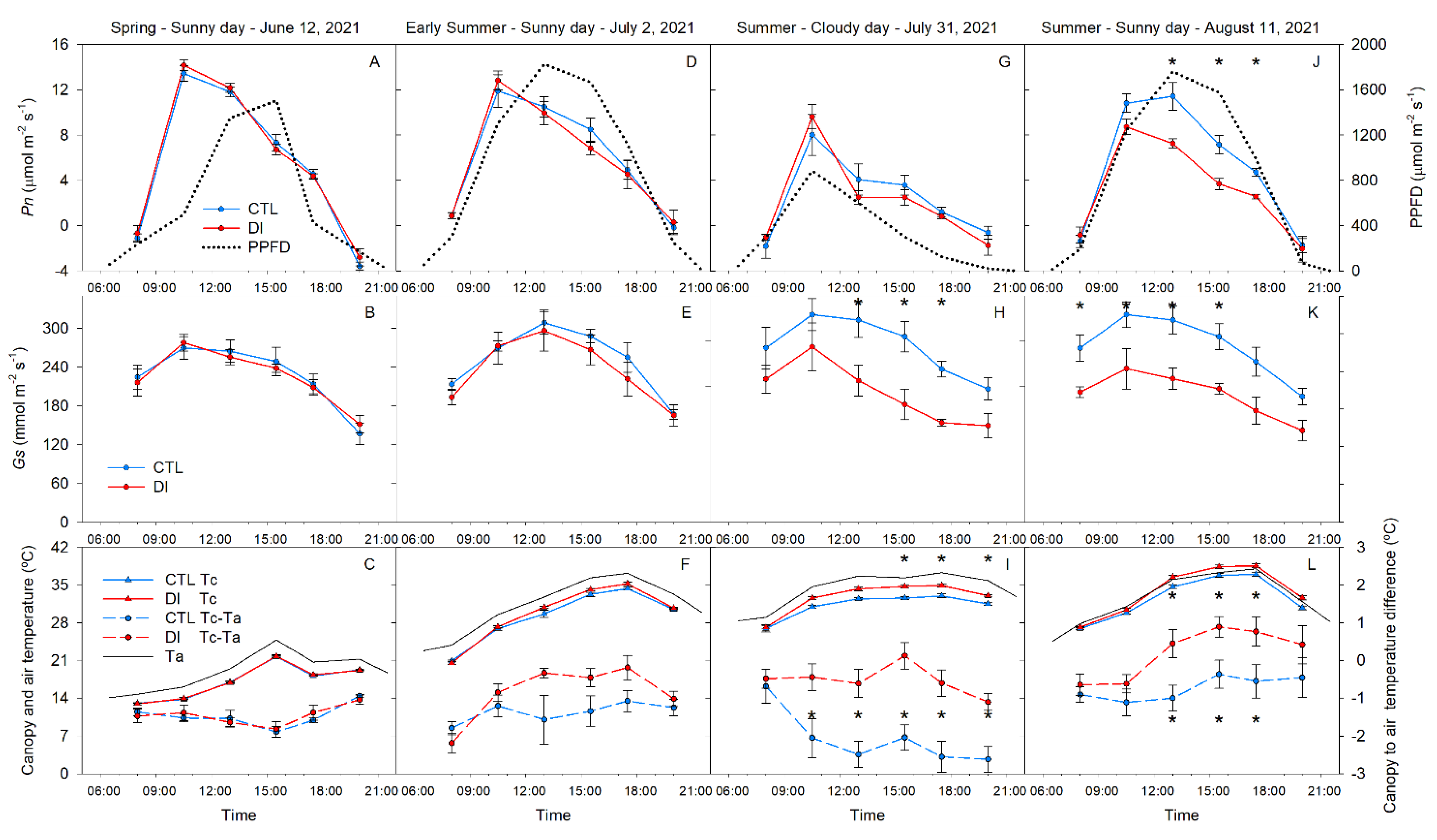
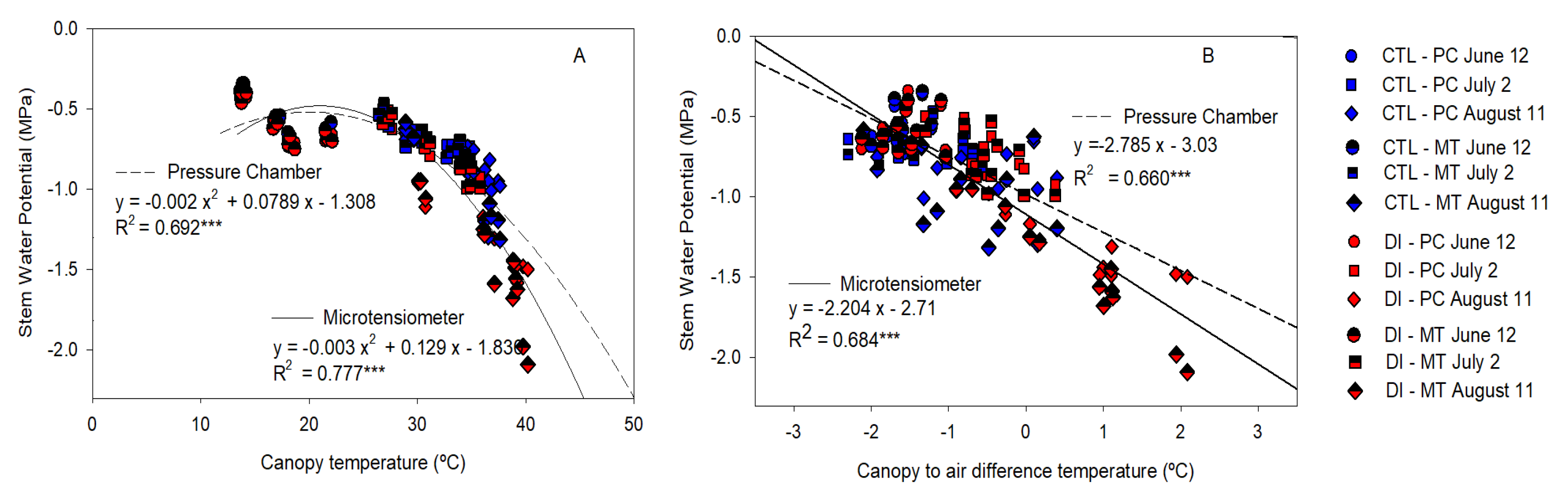
2.5. Gas Exchange and Canopy Temperature. Diurnal Pattern
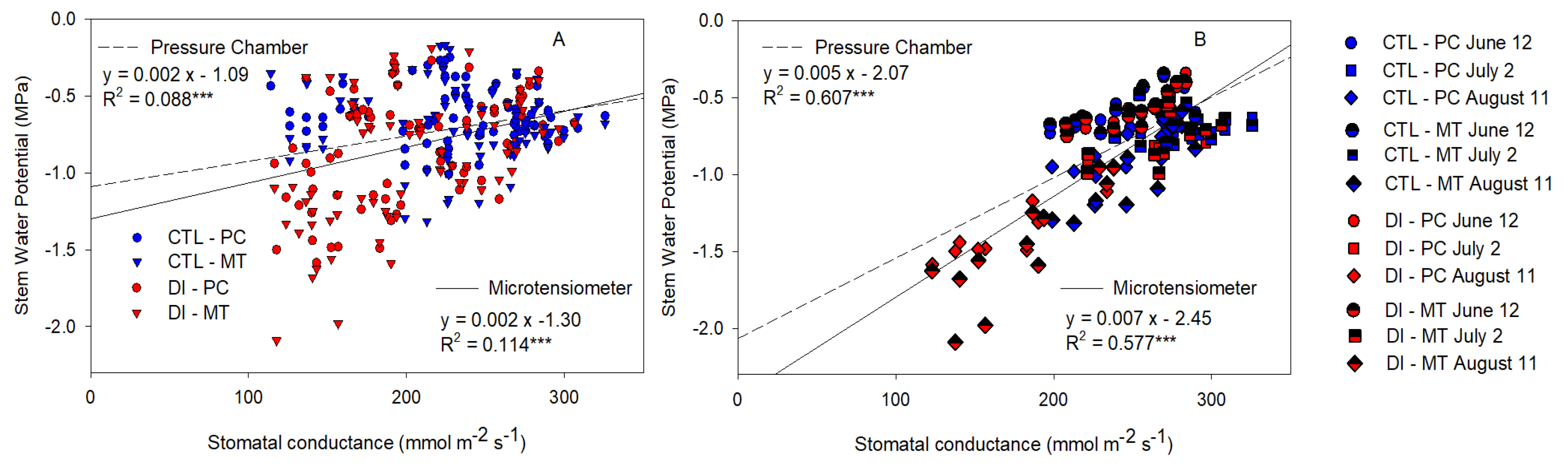
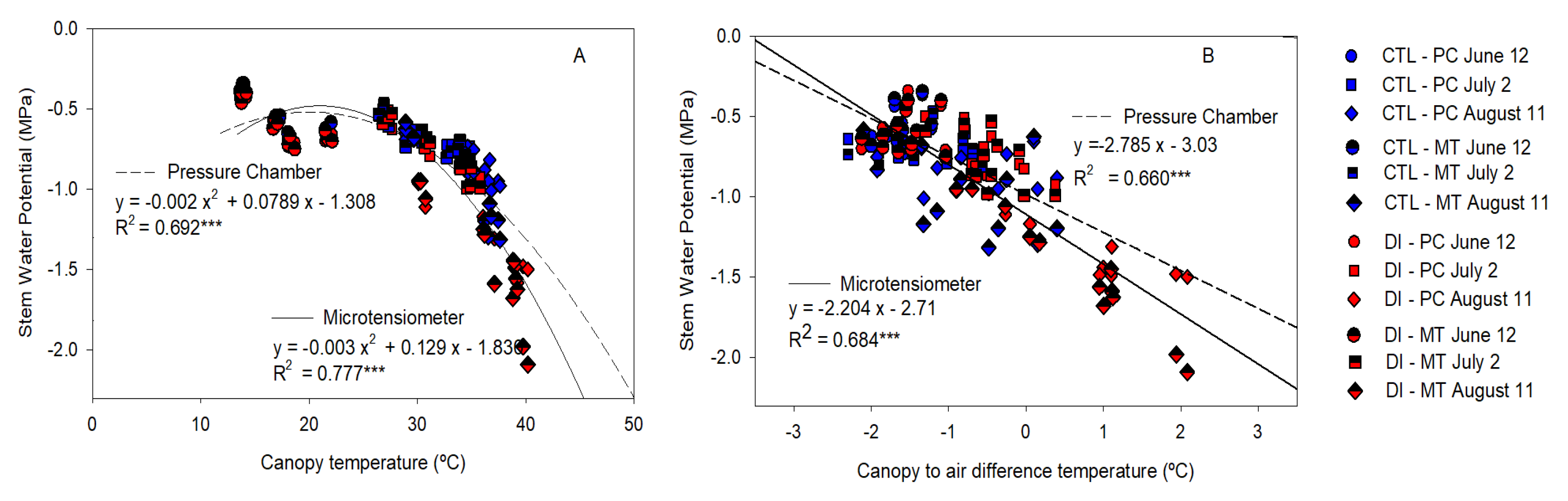
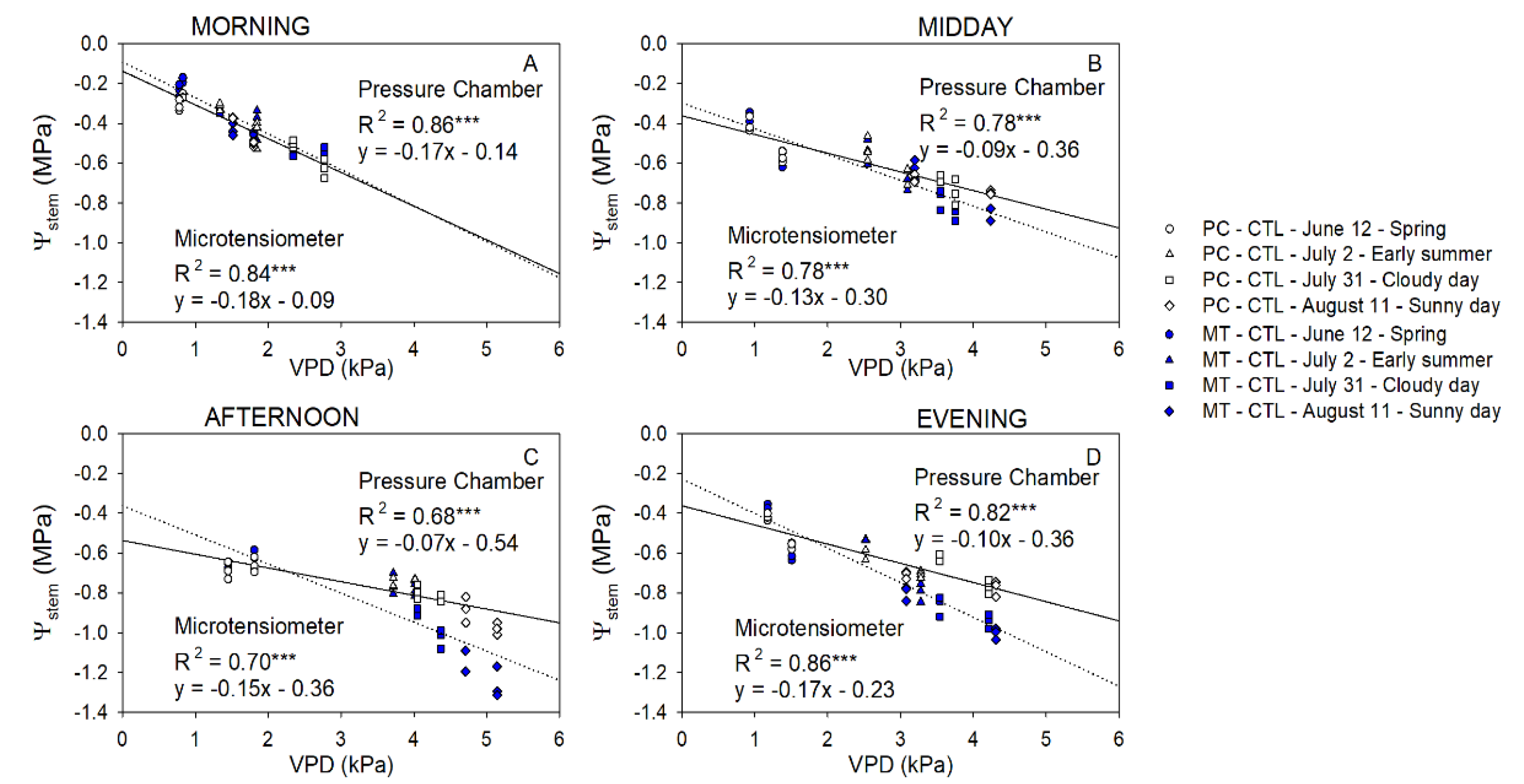
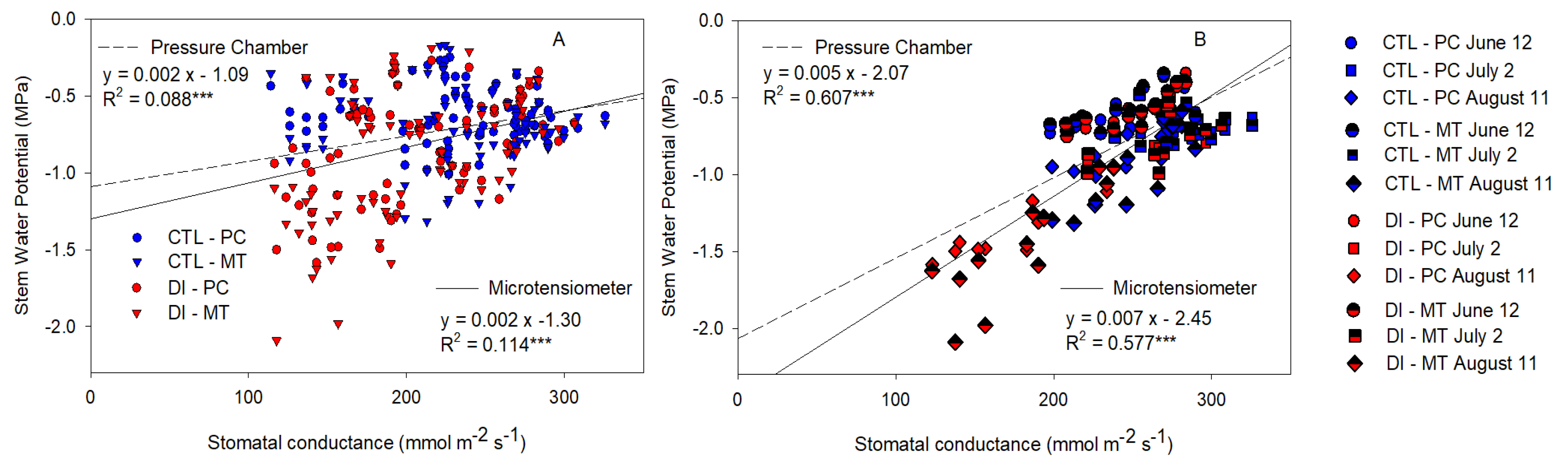
2.6. Stem Water Potential Relationship with Stomatal Conductance and Canopy Temperature
2.7. Sensitivity Analysis
3. Discussion
4. Materials and Methods
4.1. Study Site and Irrigation Treatments
4.2. Measurements
4.2.1. Environmental Data and Soil Water Content
4.2.2. Gas Exchange and Canopy Temperature
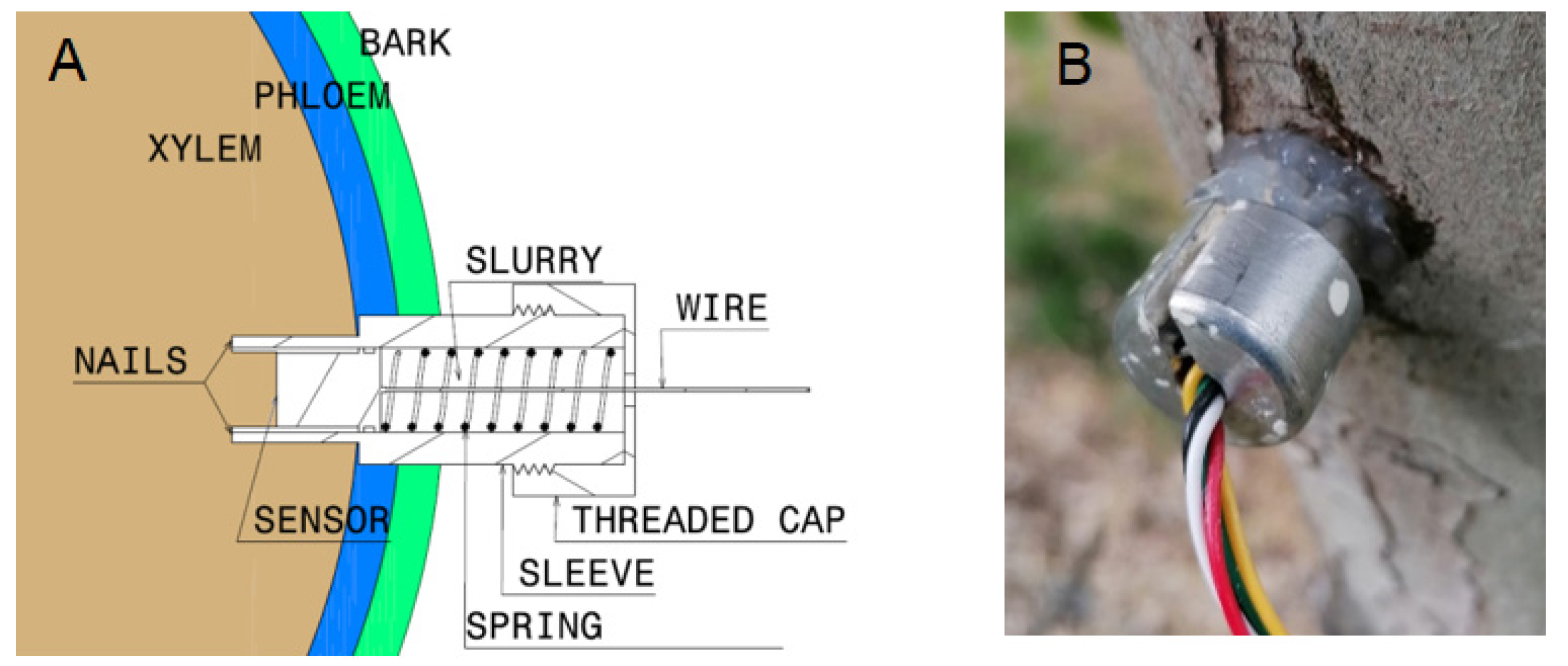
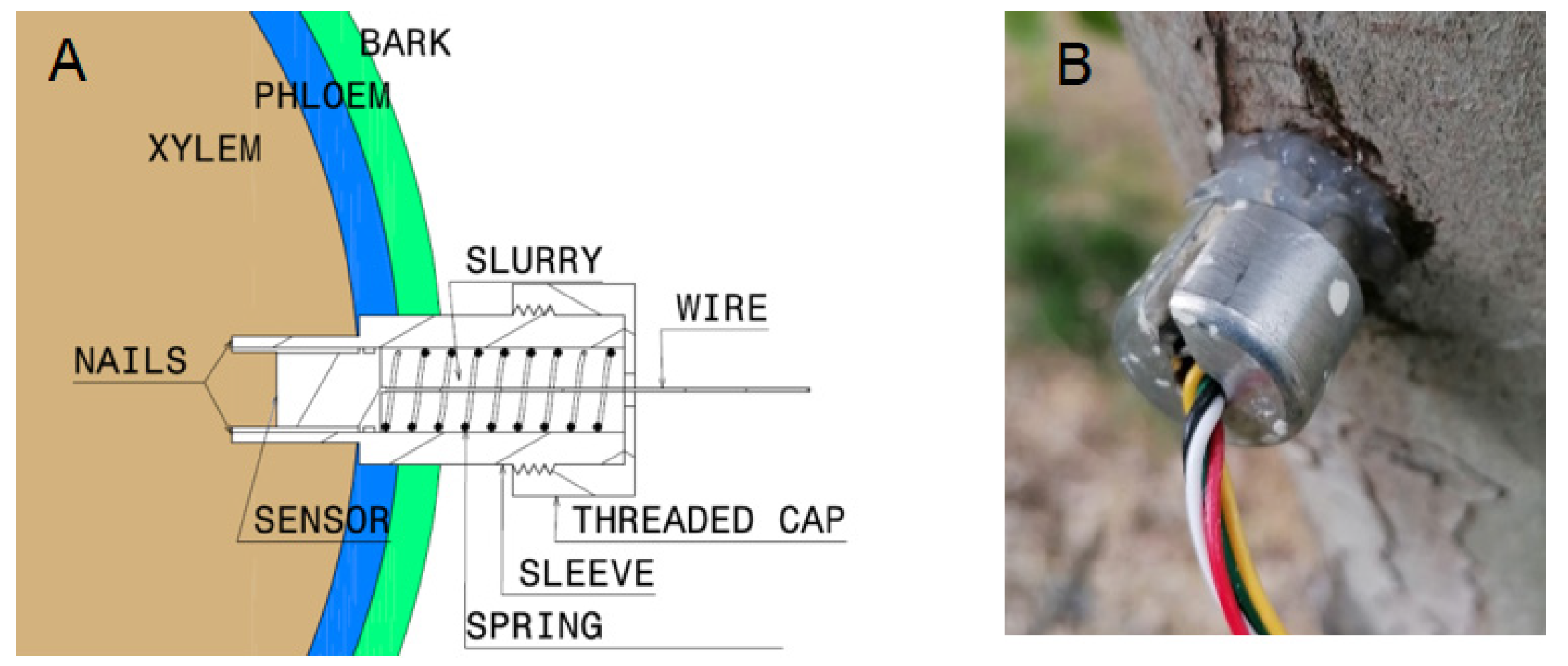
4.2.3. Stem Water Potential
4.3. Sensitivity and Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Jones, H.G. Irrigation scheduling—Comparison of soil, plant and atmosphere monitoring approaches. Acta Hortic. 2008, 792, 391–403. [Google Scholar] [CrossRef]
- Sui, R. Irrigation Scheduling Using Soil Moisture Sensors. J. Agric. Sci. 2017, 10, 1–11. [Google Scholar] [CrossRef]
- Pereira, L.S.; Paredes, P.; Jovanovic, N. Soil Water Balance Models for Determining Crop Water and Irrigation Requirements and Irrigation Scheduling Focusing on the FAO56 Method and the Dual Kc Approach. Agric. Water Manag. 2020, 241, 106357. [Google Scholar] [CrossRef]
- Fernández, J.E. Plant-Based Methods for Irrigation Scheduling of Woody Crops. Horticulturae 2017, 3, 35. [Google Scholar] [CrossRef] [Green Version]
- Shackel, K. A Plant-Based Approach to Deficit Irrigation in Trees and Vines. HortScience 2011, 46, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Naor, A. Midday stem water potential as a plant water stress indicator for irrigation scheduling in fruit trees. Acta Hortic. 2000, 447–454. [Google Scholar] [CrossRef]
- McCutchan, H.; Shackel, K.A. Stem-Water Potential as a Sensitive Indicator of Water Stress in Prune Trees (Prunus domestica L. Cv. French). J. Am. Soc. Hortic. Sci. 1992, 117, 607–611. [Google Scholar] [CrossRef] [Green Version]
- Blanco, V.; Domingo, R.; Pérez-Pastor, A.; Blaya-Ros, P.J.; Torres-Sánchez, R. Soil and Plant Water Indicators for Deficit Irrigation Management of Field-Grown Sweet Cherry Trees. Agric. Water Manag. 2018, 208, 83–94. [Google Scholar] [CrossRef]
- Scholander, P.F.; Bradstreet, E.D.; Hemmingsen, E.A.; Hammel, H.T. Sap Pressure in Vascular Plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.C. Measurement of Plant Water Status by the Pressure Chamber Technique. Irrig. Sci. 1988, 9, 289–308. [Google Scholar] [CrossRef]
- Kume, T.; Takizawa, H.; Yoshifuji, N.; Tanaka, K.; Tantasirin, C.; Tanaka, N.; Suzuki, M. Impact of Soil Drought on Sap Flow and Water Status of Evergreen Trees in a Tropical Monsoon Forest in Northern Thailand. For. Ecol. Manag. 2007, 238, 220–230. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Martel, A.B.; Dixon, S.L. Environmental Factors Influence Plant Vascular System and Water Regulation. Plants 2019, 8, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortuño, M.F.; Conejero, W.; Moreno, F.; Moriana, A.; Intrigliolo, D.S.; Biel, C.; Mellisho, C.D.; Pérez-Pastor, A.; Domingo, R.; Ruiz-Sánchez, M.C.; et al. Could Trunk Diameter Sensors Be Used in Woody Crops for Irrigation Scheduling? A Review of Current Knowledge and Future Perspectives. Agric. Water Manag. 2010, 97, 1–11. [Google Scholar] [CrossRef]
- Mancha, L.A.; Uriarte, D.; Prieto, M.d.H. Characterization of the Transpiration of a Vineyard under Different Irrigation Strategies Using Sap Flow Sensors. Water 2021, 13, 2867. [Google Scholar] [CrossRef]
- Martínez-Gimeno, M.A.; Castiella, M.; Rüger, S.; Intrigliolo, D.S.; Ballester, C. Evaluating the Usefulness of Continuous Leaf Turgor Pressure Measurements for the Assessment of Persimmon Tree Water Status. Irrig. Sci. 2017, 35, 159–167. [Google Scholar] [CrossRef] [Green Version]
- Mira-García, A.B.; Conejero, W.; Vera, J.; Ruiz-Sánchez, M.C. Leaf Water Relations in Lime Trees Grown under Shade Netting and Open-Air. Plants 2020, 9, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, H.; Turner, N.C.; Aogu, K.; Dyck, M.; Feng, H.; Si, B.; Wang, J.; Lv, J. Time and Frequency Domain Reflectometry for the Measurement of Tree Stem Water Content: A Review, Evaluation, and Future Perspectives. Agric. For. Meteorol. 2021, 306, 108442. [Google Scholar] [CrossRef]
- Pagay, V.; Santiago, M.; Sessoms, D.A.; Huber, E.J.; Vincent, O.; Pharkya, A.; Corso, T.N.; Lakso, A.N.; Stroock, A.D. A Microtensiometer Capable of Measuring Water Potentials below −10 MPa. Lab Chip 2014, 14, 2806–2817. [Google Scholar] [CrossRef]
- Pagay, V. Evaluating a Novel Microtensiometer for Continuous Trunk Water Potential Measurements in Field-Grown Irrigated Grapevines. Irrig. Sci. 2021. [Google Scholar] [CrossRef]
- Girona, J.; Marsal, J.; Lopez Velasco, G. Establishment of Stem Water Potential Thresholds for the Response of “O’Henry” Peach Fruit Growth to Water Stress during Stage III of Fruit Development. Acta Hortic. 2006, 713, 197–201. [Google Scholar] [CrossRef]
- Corell, M.; Martín-Palomo, M.J.; Girón, I.; Andreu, L.; Galindo, A.; Centeno, A.; Pérez-López, D.; Moriana, A. Stem Water Potential-Based Regulated Deficit Irrigation Scheduling for Olive Table Trees. Agric. Water Manag. 2020, 242, 106418. [Google Scholar] [CrossRef]
- Abrisqueta, I.; Conejero, W.; Valdes-Vela, M.; Vera, J.; Ortuño, M.F.; Ruiz-Sánchez, M.C. Stem Water Potential Estimation of Drip-Irrigated Early-Maturing Peach Trees under Mediterranean Conditions. Comput. Electron. Agric. 2015, 114, 7–13. [Google Scholar] [CrossRef]
- Suter, B.; Triolo, R.; Pernet, D.; Dai, Z.; Van Leeuwen, C. Modeling Stem Water Potential by Separating the Effects of Soil Water Availability and Climatic Conditions on Water Status in Grapevine (Vitis vinifera L.). Front. Plant Sci. 2019, 10, 1485. [Google Scholar] [CrossRef] [Green Version]
- De Bei, R.; Cozzolino, D.; Sullivan, W.; Cynkar, W.; Fuentes, S.; Dambergs, R.; Pech, J.; Tyerman, S. Non-Destructive Measurement of Grapevine Water Potential Using near Infrared Spectroscopy. Aust. J. Grape Wine Res. 2011, 17, 62–71. [Google Scholar] [CrossRef]
- Blaya-Ros, P.J.; Blanco, V.; Domingo, R.; Soto-Valles, F.; Torres-Sánchez, R. Feasibility of Low-Cost Thermal Imaging for Monitoring Water Stress in Young and Mature Sweet Cherry Trees. Appl. Sci. 2020, 10, 5461. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Ortega-Arévalo, C.J.; Iglesias-Contreras, M.; Moreno, J.M.; Souza, L.; Tavira, S.C.; Durán-Zuazo, V.H. Assessing the Crop-Water Status in Almond (Prunus dulcis Mill.) Trees via Thermal Imaging Camera Connected to Smartphone. Sensors 2018, 18, 1050. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, L.; Zweifel, R.; Kahmen, A. Daily Stem Diameter Variations Can Predict the Canopy Water Status of Mature Temperate Trees. Tree Physiol. 2018, 38, 941–952. [Google Scholar] [CrossRef]
- Scalisi, A.; Marino, G.; Marra, F.P.; Caruso, T.; Lo Bianco, R. A Cultivar-Sensitive Approach for the Continuous Monitoring of Olive (Olea europaea L.) Tree Water Status by Fruit and Leaf Sensing. Front. Plant Sci. 2020, 11, 340. [Google Scholar] [CrossRef] [PubMed]
- Bellvert, J.; Adeline, K.; Baram, S.; Pierce, L.; Sanden, B.L.; Smart, D.R. Monitoring Crop Evapotranspiration and Crop Coefficients over an Almond and Pistachio Orchard Throughout Remote Sensing. Remote Sens. 2018, 10, 2001. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Dugo, V.; Zarco-Tejada, P.; Nicolás, E.; Nortes, P.A.; Alarcón, J.J.; Intrigliolo, D.S.; Fereres, E. Using High Resolution UAV Thermal Imagery to Assess the Variability in the Water Status of Five Fruit Tree Species within a Commercial Orchard. Precis. Agric. 2013, 14, 660–678. [Google Scholar] [CrossRef]
- Blanco, V.; Blaya-Ros, P.J.; Castillo, C.; Soto-Vallés, F.; Torres-Sánchez, R.; Domingo, R. Potential of UAS-Based Remote Sensing for Estimating Tree Water Status and Yield in Sweet Cherry Trees. Remote Sens. 2020, 12, 2359. [Google Scholar] [CrossRef]
- De la Rosa, J.M.; Conesa, M.R.; Domingo, R.; Pérez-Pastor, A. A New Approach to Ascertain the Sensitivity to Water Stress of Different Plant Water Indicators in Extra-Early Nectarine Trees. Sci. Hortic. 2014, 169, 147–153. [Google Scholar] [CrossRef]
- Intrigliolo, D.; Bonet Perez de León, L.; Ferrer, P.; Reig, C.; Mesejo, C.; Soler, E. Usefulness of Stem Dendrometers as Continuous Water Stress Indicators of Loquat Tree Water Status. Acta Hortic. 2011, 887, 149–154. [Google Scholar] [CrossRef]
- Tuccio, L.; Lo Piccolo, E.; Battelli, R.; Matteoli, S.; Massai, R.; Scalabrelli, G.; Remorini, D. Physiological Indicators to Assess Water Status in Potted Grapevine (Vitis vinifera L.). Sci. Hortic. 2019, 255, 8–13. [Google Scholar] [CrossRef]
- Gaudin, R.; Roux, S.; Tisseyre, B. Linking the Transpirable Soil Water Content of a Vineyard to Predawn Leaf Water Potential Measurements. Agric. Water Manag. 2017, 182, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Puerto, P.; Domingo, R.; Torres, R.; Pérez-Pastor, A.; García-Riquelme, M. Remote Management of Deficit Irrigation in Almond Trees Based on Maximum Daily Trunk Shrinkage. Water Relations and Yield. Agric. Water Manag. 2013, 126, 33–45. [Google Scholar] [CrossRef]
- Conesa, M.R.; Conejero, W.; Vera, J.; Ramírez-Cuesta, J.M.; Ruiz-Sánchez, M.C. Terrestrial and Remote Indexes to Assess Moderate Deficit Irrigation in Early-Maturing Nectarine Trees. Agronomy 2019, 9, 630. [Google Scholar] [CrossRef] [Green Version]
- Naor, A.; Cohen, S. Sensitivity and Variability of Maximum Trunk Shrinkage, Midday Stem Water Potential, and Transpiration Rate in Response to Withholding Irrigation from Field-Grown Apple Trees. HortScience 2003, 38, 547–551. [Google Scholar] [CrossRef] [Green Version]
- Reid, M.; Kalcsits, L. Water Deficit Timing Affects Physiological Drought Response, Fruit Size, and Bitter Pit Development for “Honeycrisp” Apple. Plants 2020, 9, 874. [Google Scholar] [CrossRef]
- Valverdi, N.A.; Kalcsits, L. Apple Rootstock Genotype Affects Scion Responses to Water Limitations under Field Conditions. Acta Physiol. Plant. 2021, 43, 97. [Google Scholar] [CrossRef]
- Remorini, D.; Massai, R. Comparison of Water Status Indicators for Young Peach Trees. Irrig. Sci. 2003, 22, 39–46. [Google Scholar] [CrossRef]
- Santesteban, L.G.; Miranda, C.; Marín, D.; Sesma, B.; Intrigliolo, D.S.; Mirás-Avalos, J.M.; Escalona, J.M.; Montoro, A.; de Herralde, F.; Baeza, P.; et al. Discrimination Ability of Leaf and Stem Water Potential at Different Times of the Day through a Meta-Analysis in Grapevine (Vitis vinifera L.). Agric. Water Manag. 2019, 221, 202–210. [Google Scholar] [CrossRef]
- Moriana, A.; Pérez-López, D.; Prieto, M.H.; Ramírez-Santa-Pau, M.; Pérez-Rodriguez, J.M. Midday Stem Water Potential as a Useful Tool for Estimating Irrigation Requirements in Olive Trees. Agric. Water Manag. 2012, 112, 43–54. [Google Scholar] [CrossRef]
- Romero-Trigueros, C.; Gambín, J.M.B.; Nortes Tortosa, P.A.; Cabañero, J.J.A.; Nicolás Nicolás, E. Isohydricity of Two Different Citrus Species under Deficit Irrigation and Reclaimed Water Conditions. Plants 2021, 10, 2121. [Google Scholar] [CrossRef]
- Steduto, P.; Hsiao, T.C.; Fereres, E.; Raes, D. Crop Yield Response to Water. In FAO Irrigation and Drainage; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; ISBN 978-92-5-107274-5. [Google Scholar]
- Marsal, J.; Girona, J.; Naor, A. Crop Yield Response to Water. Pear. In FAO Irrigation and Drainage; Food and Agriculture Organization of the United Nations: Rome, Italy, 2012; ISBN 978-92-5-107274-5. [Google Scholar]
- Naor, A.; Stern, R.; Flaishman, M.; Gal, Y.; Peres, M. Effects of Post-Harvest Water Stress on Autumnal Bloom and Subsequent-Season Productivity in Mid-Season ‘Spadona’ Pear. J. Hortic. Sci. Biotechnol. 2006, 81, 365–370. [Google Scholar] [CrossRef]
- Marsal, J.; Mata, M.; Arbonàs, A.; Rufat, J.; Girona, J. Water stress limits for vegetative and reproductive growth of barlett pears. Acta Hortic. 2002, 596, 659–663. [Google Scholar] [CrossRef]
- Vélez-Sánchez, J.E.; Balaguera-López, H.E.; Alvarez-Herrera, J.G. Effect of Regulated Deficit Irrigation (RDI) on the Production and Quality of Pear Triunfo de Viena Variety under Tropical Conditions. Sci. Hortic. 2021, 278, 109880. [Google Scholar] [CrossRef]
- Venturi, M.; Manfrini, L.; Perulli, G.D.; Boini, A.; Bresilla, K.; Corelli Grappadelli, L.; Morandi, B. Deficit Irrigation as a Tool to Optimize Fruit Quality in Abbé Fetél Pear. Agronomy 2021, 11, 1141. [Google Scholar] [CrossRef]
- Caspari, H.W.; Behboudian, M.H.; Chalmers, D.J. Water Use, Growth, and Fruit Yield of `Hosui’ Asian Pears under Deficit Irrigation. J. Am. Soc. Hortic. Sci. 1994, 119, 383–388. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Wolf, A.; Arango-Velez, A.; Choat, B.; Chmura, D.J.; Jansen, S.; Kolb, T.; Li, S.; Meinzer, F.; Pita, P.; et al. Plant Water Potential Improves Prediction of Empirical Stomatal Models. PLoS ONE 2017, 12, e0185481. [Google Scholar] [CrossRef] [Green Version]
- Galindo, A.; Rodríguez Hernández, P.; Mellisho, C.D.; Torrecillas, E.; Moriana, A.; Cruz Pérez, Z.; Conejero, W.; Moreno, F.; Torrecillas, A. Assessment of Discretely Measured Indicators and Maximum Daily Trunk Shrinkage for Detecting Water Stress in Pomegranate Trees. Agric. For. Meteorol. 2013, 180, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Levin, A.D. Re-Evaluating Pressure Chamber Methods of Water Status Determination in Field-Grown Grapevine (Vitis spp.). Agric. Water Manag. 2019, 221, 422–429. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage, Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; ISBN 92-5-104219-5. [Google Scholar]
- Fereres, E.; Martinich, D.A.; Aldrich, T.M.; Castel, J.; Holzapfel, E.; Schulbach, H. Drip Irrigation Saves Money in Young Almond Orchards. Calif. Agric. 1982, 36, 12–13. [Google Scholar]
- Goldhamer, D.A.; Fereres, E. Irrigation Scheduling Protocols Using Continuously Recorded Trunk Diameter Measurements. Irrig. Sci. 2001, 20, 115–125. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morning | Midday | |||||||
| Ψstem | Gs | Tc-Ta | Ψstem | Gs | Tc-Ta | |||
| PC | MT | PC | MT | |||||
| SI 1 | 1.26 | 1.30 | 0.90 | 0.71 | 1.30 | 1.26 | 0.83 | 0.45 |
| CV | 0.06 | 0.09 | 0.10 | 0.68 | 0.07 | 0.07 | 0.13 | 0.45 |
| S | 19.48 | 14.44 | 9.06 | 1.05 | 17.65 | 18.16 | 6.53 | 1.02 |
| Afternoon | Evening | |||||||
| Ψstem | Gs | Tc-Ta | Ψstem | Gs | Tc-Ta | |||
| PC | MT | PC | MT | |||||
| SI 1 | 1.31 | 1.26 | 0.82 | 0.62 | 1.23 | 1.23 | 1.08 | 0.72 |
| CV | 0.04 | 0.07 | 0.12 | 0.37 | 0.04 | 0.05 | 0.20 | 0.30 |
| S | 34.02 | 18.16 | 7.00 | 1.67 | 29.93 | 23.89 | 5.52 | 2.42 |
| 12 June | 2 July | |||||||
| Ψstem | Gs | Tc-Ta | Ψstem | Gs | Tc-Ta | |||
| PC | MT | PC | MT | |||||
| SI 1 | 1.03 | 1.08 | 1.01 | 0.98 | 1.09 | 1.09 | 0.96 | 0.66 |
| CV | 0.06 | 0.08 | 0.13 | 0.39 | 0.04 | 0.05 | 0.15 | 0.73 |
| S | 17.06 | 13.81 | 8.03 | 2.53 | 27.60 | 21.14 | 6.55 | 0.91 |
| 31 July | 11 August | |||||||
| Ψstem | Gs | Tc-Ta | Ψstem | Gs | Tc-Ta | |||
| PC | MT | PC | MT | |||||
| SI 1 | 1.43 | 1.36 | 0.79 | 0.31 | 1.54 | 1.52 | 0.87 | 0.56 |
| CV | 0.06 | 0.08 | 0.11 | 0.41 | 0.05 | 0.07 | 0.16 | 0.27 |
| S | 22.64 | 16.47 | 7.24 | 0.75 | 28.30 | 22.31 | 5.51 | 2.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco, V.; Kalcsits, L. Microtensiometers Accurately Measure Stem Water Potential in Woody Perennials. Plants 2021, 10, 2780. https://doi.org/10.3390/plants10122780
Blanco V, Kalcsits L. Microtensiometers Accurately Measure Stem Water Potential in Woody Perennials. Plants. 2021; 10(12):2780. https://doi.org/10.3390/plants10122780
Chicago/Turabian StyleBlanco, Victor, and Lee Kalcsits. 2021. "Microtensiometers Accurately Measure Stem Water Potential in Woody Perennials" Plants 10, no. 12: 2780. https://doi.org/10.3390/plants10122780
APA StyleBlanco, V., & Kalcsits, L. (2021). Microtensiometers Accurately Measure Stem Water Potential in Woody Perennials. Plants, 10(12), 2780. https://doi.org/10.3390/plants10122780