
Cloning and Characterization of Two Putative P-Type ATPases from the Marine Microalga Dunaliella maritima Similar to Plant H+-ATPases and Their Gene Expression Analysis under Conditions of Hyperosmotic Salt Shock

 ,
,
Abstract
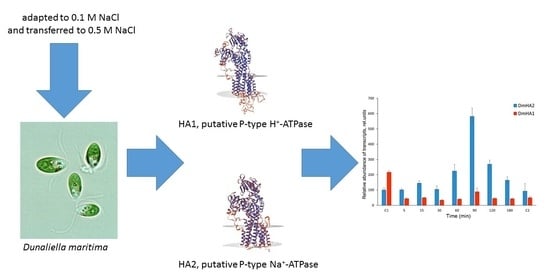
1. Introduction
2. Materials and Methods
2.1. The Object of the Study and Conditions of Cultivation
2.2. Isolation of Total RNA from D. maritima Cells
2.3. cDNA Synthesis on the Total RNA Template
2.4. Amplification of the DmHA2 Partial Coding Sequence
2.5. Amplification of the Full-Length Sequences Encoding the DmHA1 and DmHA2
2.6. Analysis of DmHA1 and DmHA2 Expression under Hyperosmotic Salt Shock
2.7. Determination of Na+ Content in D. maritima Cells under Hyperosmotic Salt Shock
2.8. Bioinformatic Methods
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oren, A. A hundred years of Dunaliella research: 1905–2005. Saline Syst. 2005, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Avron, M.; Ben-Amotz, A. (Eds.) Dunaliella: Physiology, Biochemistry, and Biotechnology; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Borowitzka, M.A.; Siva, C.J. The taxonomy of the genus Dunaliella (Chlorophyta, Dunaliella Les) with emphasis on the marine and halophilic species. J. Appl. Phycol. 2007, 19, 567–590. [Google Scholar] [CrossRef]
- Massjuk, N.P. Morphology, Taxonomy, Ecology and Geographic Distribution of the Genus Dunaliella Teod. and Prospects for Its Potential Utilization; Naykova Dumka: Kiev, Ukraine, 1973; p. 242. (In Russian) [Google Scholar]
- Ginzburg, M. Dunaliella: A green alga adapted to salt. Adv. Bot. Res. 1987, 14, 93–183. [Google Scholar]
- Ben-Amotz, A. Adaptation of the unicellular alga Dunaliella parva to a saline environment. J. Phycol. 1975, 11, 50–54. [Google Scholar] [CrossRef]
- Balnokin, Y.V.; Mazel, Y.Y. Permeability of the plasma membrane to sodium ions in halophilic algae of the genus Dunaliella. Soviet Plant Physiol. 1985, 32, 23–30. [Google Scholar]
- Wegmann, K. Osmotic regulation of photosynthetic glycerol production in Dunaliella. Biochim. Biophys. Acta 1971, 234, 317–323. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Avron, M. The role of glycerol in the osmotic regulation of the halophilic alga Dunaliella parva. Plant Physiol. 1973, 51, 875–878. [Google Scholar] [CrossRef]
- Borowitzka, L.J.; Brown, A.D. The salt relations of marine and halophilic species of the unicellular green alga, Dunaliella. The role of glycerol as a compatible solute. Arch. Microbiol. 1974, 96, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Gimmler, H.; Hartung, W. Low permeability of the plasma membrane of Dunaliella parva for solutes. J. Plant Physiol. 1988, 133, 165–172. [Google Scholar] [CrossRef]
- Johnson, M.K.; Johnson, E.J.; MacElroy, R.D.; Speer, H.L.; Bruff, B.S. Effects of salts on the halophilic alga Dunaliella viridis. J. Bacteriol. 1968, 95, 1461–1468. [Google Scholar] [CrossRef]
- Pollard, A.; Jones, W.G.W. Enzyme activities in concentrated solutions of glycinbetain and other solutes. Planta 1979, 144, 291–298. [Google Scholar] [CrossRef]
- Balnokin, Y.V.; Medvedev, A.V. Effects of ions on electron transport in chloroplasts of the halophilic alga Dunaliella. Soviet Plant Physiol. 1980, 27, 1229–1236. [Google Scholar]
- Flowers, T.J.; Dalmond, D. Protein synthesis in halophytes–the influence of potassium sodium, and magnesium in vitro. Plant Soil 1992, 146, 153–161. [Google Scholar] [CrossRef]
- Maathuis, F.J.M.; Amtmann, A. K+ nutrition and Na+ toxicity: The basis of cellular K+/Na+ ratios. Ann. Bot. 1999, 84, 123–133. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohner, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Bhandal, I.S.; Malik, C.P. Potassium estimation uptake, and its role in the physiology and metabolism of flowering plants. Int. Rev. Cytol. 1988, 110, 205–254. [Google Scholar] [CrossRef]
- Volkov, V.; Amtmann, A. Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, has specific root ion-channel features supporting K+/Na+ homeostasis under salinity stress. Plant J. 2006, 48, 342–353. [Google Scholar] [CrossRef]
- Huh, G.H.; Damsz, B.; Matsumoto, T.K.; Reddy, M.P.; Rus, A.M.; Ibeas, J.I.; Narasimhan, M.L.; Bressan, R.A.; Hasegawa, P.M. Salt causes ion disequilibrium-induced programmed cell death in yeast and plants. Plant J. 2002, 29, 649–659. [Google Scholar] [CrossRef]
- Shabala, S. Salinity and programmed cell death: Unraveling mechanisms for ion specific signaling. J. Exp. Bot. 2009, 60, 709–711. [Google Scholar] [CrossRef]
- Demidchik, V.; Cuin, T.A.; Svistunenko, D.; Smith, S.J.; Miller, A.J.; Shabala, S.; Sokolik, A.; Yurin, V. Arabidopsis root K+-efflux conductance activated by hydroxyl radicals: Single-channel properties, genetic basis and involvement in stress-induced cell death. J. Cell Sci. 2010, 123, 1468–1479. [Google Scholar] [CrossRef]
- Ehrenfeld, J.; Cousin, J.L. Ionic regulation of the unicellular green alga Dunaliella tertiolecta. J. Membr. Biol. 1982, 70, 47–57. [Google Scholar] [CrossRef]
- Bental, M.; Degani, H.; Avron, M. 23Na NMR studies of the intracellular sodium ion concentration in the halotolerant alga Dunaliella salina. Plant Physiol. 1988, 87, 813–817. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.; Avron, M. Determination of intracellular osmotic volume and sodium concentration in Dunaliella. Plant Physiol. 1985, 78, 817–820. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.; Pick, U.; Avron, M. Characterization and reconstitution of the Na+/H+ antiporter from the plasma membrane of the halophilic alga Dunaliella. Biochim. Biophys. Acta 2001, 1504, 423–431. [Google Scholar] [CrossRef]
- Pick, U.; Karni, L.; Avron, M. Determination of ion content and ion fluxes in the halotolerant alga Dunaliella salina. Plant Physiol. 1986, 81, 92–96. [Google Scholar] [CrossRef]
- Shumkova, G.A.; Popova, L.G.; Balnokin, Y.V. Export of Na+ from cells of the halotolerant microalga Dunaliella maritima: Na+/H+ antiporter or primary Na+-pump? Biochemistry 2000, 65, 1080–1087. [Google Scholar]
- Pardo, J.M.; Cubero, B.; Leidi, E.O.; Quintero, F.J. Alkali cation exchanger: Roles in cellular homeostasis and stress tolerance. J. Exp. Bot. 2006, 57, 1181–1199. [Google Scholar] [CrossRef]
- Rodriguez-Rosales, M.P.; Galvez, F.J.; Huertas, R.; Aranda, M.N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/proton antiporters. Plant Signal. Bahavior 2009, 4, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Hamamoto, S.; Uozumi, N. Sodium transport system in plant cells. Front. Plant Sci. 2013, 4, 410. [Google Scholar] [CrossRef]
- Gaxiola, R.A.; Palmgren, M.G.; Schumacher, K. Plant proton pumps. FEBS Lett. 2007, 581, 2204–2214. [Google Scholar] [CrossRef]
- Morsomme, P.; Boutry, M. The plant plasma membrane H+-ATPase: Structure, function and regulation. Biochim. Biophys. Acta 2000, 1465, 1–16. [Google Scholar] [CrossRef]
- Duby, G.; Boutry, M. The plant plasma membrane proton pump ATPase: A highly regulated P-type ATPase with multiple physiological roles. Pfluggers Arch.–Eur. J. Physiol. 2009, 457, 645–655. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Habets, M.; Breuninger, H.; Dolan, L.; Offringa, R.; van Duijn, B. Evolutionary and functional analysis of a Chara plasma membrane H+-ATPase. Front. Plan Sci. 2020, 10, 1707. [Google Scholar] [CrossRef]
- Ohta, H.; Shirakawa, H.; Uchida, K.; Yoshida, M.; Matuo, Y.; Enami, I. Cloning and sequencing of the gene encoding the plasma membrane H+-ATPase from an acidophilic red alga, Cyanidium caldarium. Biochim. Biophys. Acta 1997, 1319, 9–13. [Google Scholar] [CrossRef]
- Harada, A.; Fukuhara, T.; Takagi, S. Photosynthetic control of the plasma membrane H+-ATPase in Vallisneria leaves. II. Presence of putative isogenes and a protein equipped with a C-terminal autoinhibitory domain. Planta 2002, 214, 870–876. [Google Scholar] [CrossRef]
- Masashi, T. Cell physiological aspects of the plasma membrane electrogenic H+ pump. J. Plant Res. 2003, 116, 419–442. [Google Scholar] [CrossRef]
- Matsuzaki, M.; Misumi, O.; Shin-I, T.; Maruyama, S.; Takahara, M.; Miyagishima, S.Y.; Mori, T.; Nishida, K.; Yagisawa, F.; Nishida, K.; et al. Genome sequence of the ultrasmall unicellular red alga Cyanidioschyzon merolae 10D. Nature 2004, 428, 653–657. [Google Scholar] [CrossRef]
- Prochnik, S.E.; Umen, J.; Nedelcu, A.M.; Hallmann, A.; Miller, S.M.; Nishii, I.; Ferris, P.; Kuo, A.; Mitros, T.; Fritz-Laylin, L.K.; et al. Genomic analysis of organismal complexity in the multicellular green alga Volvox carteri. Science 2010, 329, 223–226. [Google Scholar] [CrossRef]
- Balnokin, Y.; Popova, L.; Gimmler, H. Further evidence for an ATP-driven sodium pump in the marine alga Tetraselmis (Platymonas) viridis. J. Plant Physiol. 1997, 150, 264–270. [Google Scholar] [CrossRef]
- Skou, J.C. Nobel Lecture. The identification of the sodium pump. Biosci. Rep. 1998, 18, 155–169. [Google Scholar] [CrossRef] [PubMed]
- Scheiner-Bobis, G. The sodium pump. Eur. J. Biochem. 2002, 269, 2424–2433. [Google Scholar] [CrossRef] [PubMed]
- Glinn, I.M. A hundred years of sodium pumping. Annu. Rev. Physiol. 2002, 64, 1–18. [Google Scholar] [CrossRef]
- Jorgensen, P.L.; Hakansson, K.O.; Karlish, J.D. Structure and mechanism of Na, K-ATPase: Functional site and their interactions. Annu. Rev. Physiol. 2003, 65, 817–849. [Google Scholar] [CrossRef] [PubMed]
- Benito, B.; Garciadeblas, B.; Rodriguez-Navarro, A. Potassium- or sodium-efflux ATPase, a key enzyme in the evolution of fungi. Microbiology 2002, 148, 933–941. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Navarro, A.; Benito, B. Sodium or potassium efflux ATPase. A fungal, bryophyte, and protozoal ATPase. Biochim. Biophys. Acta 2010, 1798, 1841–1853. [Google Scholar] [CrossRef] [PubMed]
- Scarborough, G.A. Structure and function of the P-type ATPases. Curr. Opin. Cell Biol. 1999, 11, 517–522. [Google Scholar] [CrossRef]
- Palmgren, M.G.; Nissen, P. P-type ATPases. Annu. Rev. Biophys. 2011, 40, 243–266. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, M.G.; Harper, J.F. Pumping with plant P-type ATPases. J. Exp. Bot. 1999, 50, 883–893. [Google Scholar] [CrossRef]
- Popova, L.G.; Shumkova, G.A.; Andreev, I.M.; Balnokin, Y.V. Functional identification of electrogenic Na+-translocating ATPase in the plasma membrane of the halotolerant microalga Dunaliella maritima. FEBS Lett. 2005, 579, 5002–5006. [Google Scholar] [CrossRef]
- Popova, L.G.; Kornilova, A.G.; Shumkova, G.A.; Andreev, I.M.; Balnokin, Y.V. Na+-transporting ATPase in the plasma membrane of halotolerant microalga Dunaliella maritima operates as a Na+-uniporter. Russ. J. Plant Physiol. 2006, 53, 474–480. [Google Scholar] [CrossRef]
- Wolf, A.H.; Slayman, C.W.; Gradmann, D. Primary structure of the plasma membrane H+-ATPase from the halotolerant alga Dunaliella bioculata. Plant Mol. Biol. 1995, 28, 657–666. [Google Scholar] [CrossRef]
- Weiss, M.; Pick, U. Primary structure and effect of pH on the expression of the plasma membrane H+-ATPase from Dunaliella acidophila and Dunaliella salina. Plant Physiol. 1996, 112, 1693–1702. [Google Scholar] [CrossRef] [PubMed]
- Raschke, B.C.; Wolf, A.H. Molecular cloning of a P-type Ca(2+)-ATPase from the halotolerant alga Dunaliella bioculata. Planta 1996, 200, 78–84. [Google Scholar] [CrossRef]
- Popova, L.G.; Belyaev, D.V.; Shuvalov, A.V.; Yurchenko, A.A.; Matalin, D.A.; Khramov, D.E.; Orlova, Y.V.; Balnokin, Y.V. In silico analysis of transcriptomes of the marine green microalga Dunaliella tertiolecta: Identification of sequences encoding P-type ATPases. Mol. Biol. 2018, 52, 520–531. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. Algae Base. World-Wide Electronic Publication, National University of Ireland, Galway. 2021. Available online: http://www.algaebase.org (accessed on 1 December 2021).
- Balnokin, Y.V.; Medvedev, A.V. Na+, K+, and H+ transport across the plasmalemma of K+-deficient cells of the halophilic alga Dunaliella maritima. Soviet Plant Physiol. 1984, 31, 805–809. [Google Scholar]
- Popova, L.G.; Shumkova, G.A.; Andreev, I.M.; Balnokin, Y.V. Na+-dependent electrogenic ATPase from the plasma membrane of the halotolerant microalga Dunaliella maritima. Dokl. Biochem. Biophys. 2000, 375, 235–238. [Google Scholar] [CrossRef]
- Massjuk, N.P. New taxa of the genus Dunaliella Teod. Part I. Ukr. Bot. J. 1973, 30, 175–178. [Google Scholar]
- de Vries, S.; Hoge, H.; Bisseling, T. Isolation of total and polysomal RNA from plant tissues. In Plant Molecular Biology Manual; Gelvin, S.B., Schilperoort, R.A., Verma, D.P.S., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1988; pp. 1–13. [Google Scholar]
- Ochman, H.; Gerber, A.S.; Hartl, D.L. Genetic applications of an inverse polymerase chain reaction. Genetics 1988, 120, 621–623. [Google Scholar] [CrossRef]
- Okamoto, H.; Suzuki, Z. Intracellular concentration of ions in a halophilic strain of Chlamydomonas. I. Concentration of Na+, K+ and Cl− in the cell. Z. Allg. Microbiol. 1964, 4, 350–357. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetic analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Axelsen, K.; Palmgren, M.G. Evolution and substrate specificities in the P-type ATPase superfamily. J. Mol. Evol. 1998, 46, 84–101. [Google Scholar] [CrossRef]
- Møller, J.V.; Juul, B.; le Maire, M. Structural organization, ion transport and energy transduction of P-type ATPases. Biochim. Biophys. Acta 1996, 1286, 1–51. [Google Scholar] [CrossRef]
- Thever, M.D.; Saier, M.H., Jr. Bioinformatic characterization of P-type ATPase encoded within the fully sequenced genomes of 26 eukaryotes. J. Membr. Biol. 2009, 229, 115–130. [Google Scholar] [CrossRef][Green Version]
- Kaim, G.; Dimroth, P. Construction, expression and characterization of a plasmid-encoded Na+-specific ATPase hybrid consisting of Propionigenium modestum F0-ATPase and Escherichia coli F1-ATPase. Eur. J. Biochem. 1994, 222, 615–623. [Google Scholar] [CrossRef]
- Wada, M.; Satoh, S.; Kasamo, K.; Fujii, T. Presence of a Na+-activated ATPase in the plasma membrane of the marine raphidophycean Heterosigma akashiwo. Plant Cell Physiol. 1989, 30, 923–928. [Google Scholar] [CrossRef]
- Shono, M.; Wada, M.; Fujii, T. Partial purification of a Na+-ATPase from the plasma membrane of the marine alga Heterosigma akashiwo. Plant Physiol. 1995, 108, 1615–1621. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Iizumi, K.; Mikami, Y.; Hashimoto, M.; Nara, T.; Hara, Y.; Aoki, T. Molecular cloning and characterization of ouabain-insensitive Na+-ATPase in the parasitic protist Trypanosoma cruzi. Biochim. Biophys. Acta 2006, 1758, 738–746. [Google Scholar] [CrossRef] [PubMed]
- Uji, T.; Hirata, R.; Mikami, K.; Mizuta, H.; Saga, N. Molecular characterization and expression analysis of sodium pump genes in the marine red alga Porphyra yezoensis. Mol. Biol. Rep. 2012, 39, 7973–7980. [Google Scholar] [CrossRef] [PubMed]
- Fraile-Escanciano, A.; Garciadeblas, B.; Rodriguez-Navarro, A.; Benito, B. Role of ENA ATPase in Na+ efflux at high pH in bryophytes. Plant Mol. Biol. 2009, 71, 599–608. [Google Scholar] [CrossRef] [PubMed]
- Shono, M.; Wada, M.; Hara, Y.; Fujii, T. Molecular cloning of Na+-ATPase cDNA from a marine alga, Heterosigma akashiwo. Biochim. Biophys. Acta 2001, 1511, 193–199. [Google Scholar] [CrossRef]
- Balnokin, Y.V.; Popova, L.G.; Pagis, L.Y.; Andreev, I.M. The Na+-translocating ATPase in the plasma membrane of the marine microalga Tetraselmis viridis catalyzes a Na+/H+ exchange. Planta 2004, 219, 332–337. [Google Scholar] [CrossRef]
- Pagis, L.Y.; Popova, L.G.; Andreev, I.M.; Balnokin, Y.V. Ion specificity of Na+-transporting systems in the plasma membrane of the halotolerant alga Tetraselmis (Platymonas) viridis. Russ. J. Plant Physiol. 2001, 48, 281–286. [Google Scholar] [CrossRef]
- Zakhozhii, I.G.; Matalin, D.A.; Popova, L.G.; Balnokin, Y.V. Responses of photosynthetic apparatus of the halotolerant microalga Dunaliella maritima to hyperosmotic salt shock. Russ. J. Plant Physiol. 2012, 59, 42–49. [Google Scholar] [CrossRef]
- Chen, H.; Lu, Y.; Jiang, J.-G. Comparative analysis on the key enzymes of the glycerol cycle metabolic pathway in Dunaliella salina under osmotic stressess. PLoS ONE 2012, 7, e37578. [Google Scholar] [CrossRef]
- Walker, N.A.; Sanders, D. Sodium-coupled solute transport in charophyte algae: A general mechanism for transport energization in plant cells? Planta 1991, 185, 443–445. [Google Scholar] [CrossRef]
- Chan, C.X.; Reyes-Prieto, A.; Bhattacharya, D. Red and green algal origin of diatom membrane transporters: Insights into environmental adaptation and cell evolution. PLoS ONE 2011, 6, e29138. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.X.; Zäuner, A.; Wheeler, G.; Grossman, A.R.; Prochnik, S.E.; Blouin, N.A.; Zhuang, Y.; Benning, C.; Berg, G.M.; Yarish, C.; et al. Analysis of Porphyra membrane transporters demonstrates gene transfer among photosynthetic eukaryotes and numerous sodium-coupled transport systems. Plant Physiol. 2012, 158, 2001–2012. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A.; Beardall, J. Energizing the plasmalemma of marine photosynthetic organisms: The role of primary active transport. J. Mar. Biol. Assoc. UK 2020, 100, 333–346. [Google Scholar] [CrossRef]
- Ambesi, A.; Miranda, M.; Petrov, V.V.; Slayman, C.W. Biogenesis and function of the yeast plasma membrane H+-ATPase. J. Exp. Biol. 2000, 203, 155–160. [Google Scholar] [CrossRef]
- Van Der Heyden, N.; Docampo, R. Proton and sodium pumps regulate the plasma membrane potential of different stages of Trypanosoma cruzi. Mol. Biochem. Parasitol. 2002, 120, 127–139. [Google Scholar] [CrossRef]
- Ruiz, A.; Ariño, J. Function and regulation of the Saccharomyces cerevisiae ENA sodium ATPase system. Eukaryot. Cell 2007, 6, 2175–2181. [Google Scholar] [CrossRef] [PubMed]
- Balnokin, Y.V. Ion homeostasis and osmoregulation in halotolerant microalgae. Russ. J. Plant Physiol. 1993, 40, 498–506. [Google Scholar]
- Katz, A.; Waridel, P.; Shevchenko, A.; Pick, U. Saltr-induced changes in the plasma membrane proteome of the halotolerant alga Dunaliella salina as revealed by Blue-Native gel electrophoresis and nanoLC-MC/MS analysis. Mol. Cell. Proteom. 2007, 6, 1459–1472. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, H.; Toyoshima, C. Homology modeling of the cation binding sites of Na+, K+-ATPase. Proc. Natl. Acad. Sci. USA 2002, 99, 15977–15982. [Google Scholar] [CrossRef] [PubMed]
- Palmgren, M.G.; Buch-Pedersen, M.J.; Møller, A.L. Mechanism of proton pumping by plant plasma membrane H+-ATPase. Ann. N. Y. Acad. Sci. 2003, 986, 188–197. [Google Scholar] [CrossRef]
- Bublitz, M.; Poulsen, H.; Morth, J.P.; Nissen, P. In and out of the cation pumps: P-type ATPase structure revisited. Curr. Opin. Struct. Biol. 2010, 20, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, H.K.; Anteli, A.; Fink, G.B.; Buckley, C.M.; Dorman, T.E.; LeVitre, J.; Davidow, L.S.; Mao, J.I.; Moir, D.T. The yeast secretory pathway is perturbed by mutations in PMR1, a member of a Ca2+-ATPase family. Cell 1989, 58, 133–145. [Google Scholar] [CrossRef]
- Haro, R.; Garciadeblas, B.; Rodriguez-Navarro, A. A novel P-type ATPase from yeast involved in sodium transport. FEBS Lett. 1991, 291, 189–191. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | DmHA1 | DmHA2 |
|---|---|---|
| Protein accession number (GenBank ID) | QEH60479.1 | AQM50087.1 |
| Number of amino acids | 1131 | 923 |
| Subunit size (kDa) | 123.4 | 100 |
| pI | 5.35 | 5.62 |
| GRAVY index * | 0.030 | 0.166 |
| Subcellular localization | plasma membrane | plasma membrane |
| The proteins with highest homology scores ** | P54211.1 (100% identity) *** P54210.1 (75% identity) *** | ABB88698.1 (99% identity) *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matalin, D.A.; Khramov, D.E.; Shuvalov, A.V.; Volkov, V.S.; Balnokin, Y.V.; Popova, L.G. Cloning and Characterization of Two Putative P-Type ATPases from the Marine Microalga Dunaliella maritima Similar to Plant H+-ATPases and Their Gene Expression Analysis under Conditions of Hyperosmotic Salt Shock. Plants 2021, 10, 2667. https://doi.org/10.3390/plants10122667
Matalin DA, Khramov DE, Shuvalov AV, Volkov VS, Balnokin YV, Popova LG. Cloning and Characterization of Two Putative P-Type ATPases from the Marine Microalga Dunaliella maritima Similar to Plant H+-ATPases and Their Gene Expression Analysis under Conditions of Hyperosmotic Salt Shock. Plants. 2021; 10(12):2667. https://doi.org/10.3390/plants10122667
Chicago/Turabian StyleMatalin, Dmitrii A., Dmitrii E. Khramov, Alexey V. Shuvalov, Vadim S. Volkov, Yurii V. Balnokin, and Larissa G. Popova. 2021. "Cloning and Characterization of Two Putative P-Type ATPases from the Marine Microalga Dunaliella maritima Similar to Plant H+-ATPases and Their Gene Expression Analysis under Conditions of Hyperosmotic Salt Shock" Plants 10, no. 12: 2667. https://doi.org/10.3390/plants10122667
APA StyleMatalin, D. A., Khramov, D. E., Shuvalov, A. V., Volkov, V. S., Balnokin, Y. V., & Popova, L. G. (2021). Cloning and Characterization of Two Putative P-Type ATPases from the Marine Microalga Dunaliella maritima Similar to Plant H+-ATPases and Their Gene Expression Analysis under Conditions of Hyperosmotic Salt Shock. Plants, 10(12), 2667. https://doi.org/10.3390/plants10122667