
Leaf Vein Morphological Variation in Four Endangered Neotropical Magnolia Species along an Elevation Gradient in the Mexican Tropical Montane Cloud Forests

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
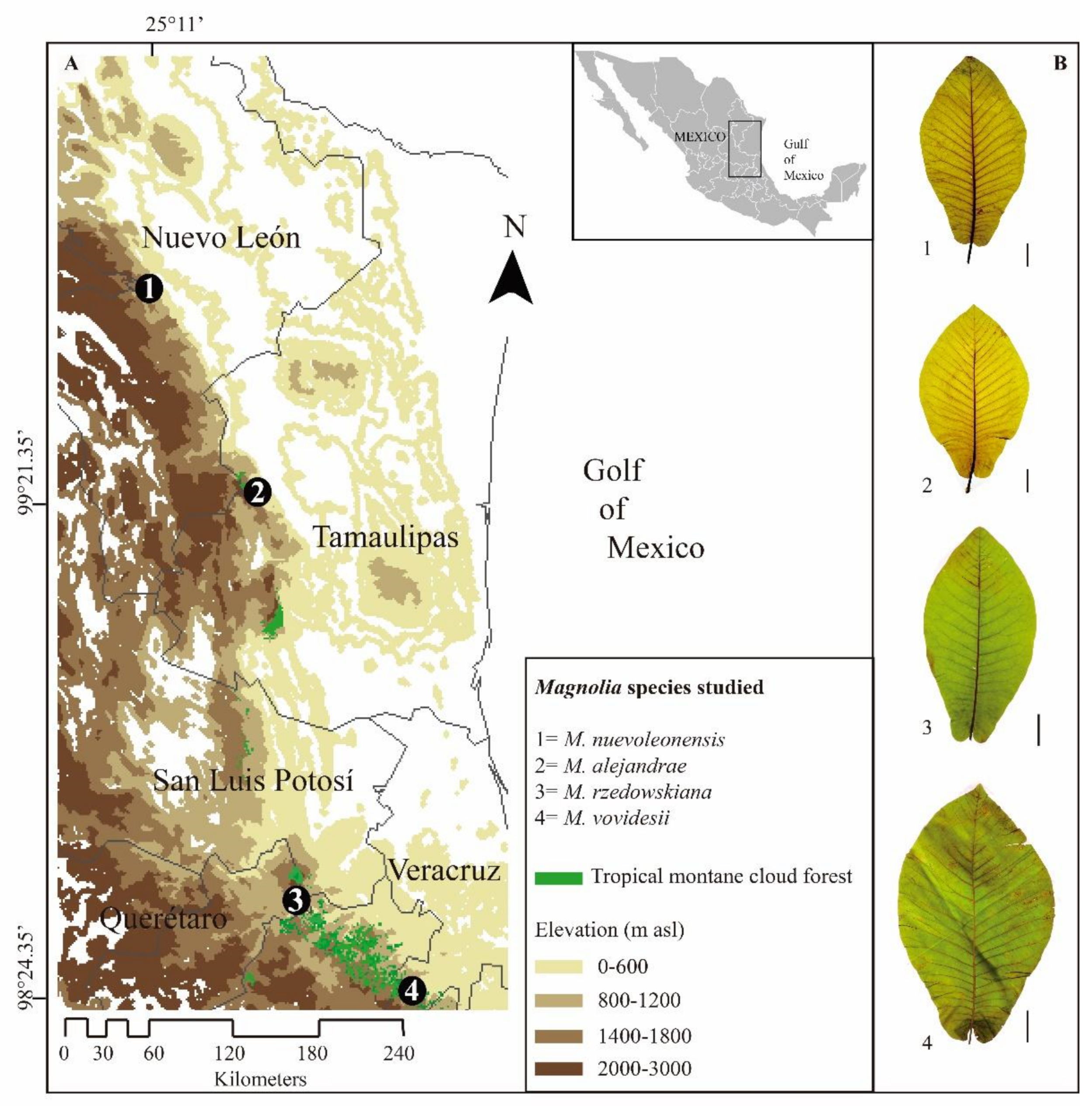
2.1. Study Forests
- La Trinidad locality shows a natural distribution (<4 km2) of endangered Magnolia nuevoleonensis (15–20 m tall), with obovate leaves from 25–35 (–40) × 15–23 (–25) cm [15]. The high-canopy (≤25 m) is composed of Quercus rysophylla Weath., Q. pinnativenulosa C.H. Mull., Q. sartorii Liebm., Taxus globosa Schltdl., Meliosma alba (Schltdl.) Walp., Tilia mexicana Schltdl., and Carpinus tropicalis subsp. mexicana Furlow. The study site is affected by deforestation and grazing [16].
- The Los San Pedros locality is characterized by the presence of critically endangered M. alejandrae (8–16 m tall), with obovate leaves from (27–) 30–45 (–55) × 12–20 cm [17]. The study site is dominated by Quercus rysophylla, Q. affinis Scheidw., Meliosma alba, Liquidambar styraciflua L., Carpinus tropicalis (Donn. Sm.) Lundell subsp. mexicana Furlow, Tilia mexicana, Ostrya virginiana subsp. guatemalensis (H.J.P. Winkl.) A.E. Murray, Carya ovata var. mexicana (Engelm.) W.E. Manning, and Pinus pseudostrobus Brongn. This is one of the less disturbed Magnolia forests in Mexico.
- La Yesca locality is characterized by the presence of endangered M. rzedowskiana (8–25 m tall) with ovate-oblong leaves from 25–40 (–50) × 15–25 (–27) cm [18]. The study site is dominated by Cupressus lusitanica Mill., Liquidambar styraciflua, and Pinus spp. The forest is highly disturbed, mainly by grazing, preventing natural regeneration.
- El Batda locality is characterized by the presence of the endangered M. vovidesii (8–25 m tall) with ovate-oblong leaves from 25–60 × 16–30 (35) cm [19]. The tree species composition is dominated by Liquidambar styraciflua, Pinus patula Schltdl. & Cham., P. greggii Engelm. ex Parl, Befaria aestuans L., Quercus meavei Valencia-A., Sabás & Soto, Q. delgadoana S. Valencia, Nixon & L. M. Kelly, and Q. trinitatis Trel. The mid-canopy is composed of M. schiedeana Schltdl., Podocarpus reichei J. Buchholz & N.E. Gray, Clethra mexicana DC., Dicksonia sellowiana Hook. var. arachneosa Sodiro, Alsophila firma (Baker) D.S. Conant, Cyathea fulva (M. Martens & Galeotti) Fée, and Cyathea bicrenata Liebm. The threatened Magnolia species is affected by logging and grazing and is often used in traditional medicine for heart diseases [2].

2.2. Leaf Sampling Design
2.3. Leaf-Clearing and Digitalization Methods
2.4. Leaf Vein Measurement
2.5. Climate Data
2.6. Associating Climatic Factors, Leaf Vein Traits, and Altitude
3. Results
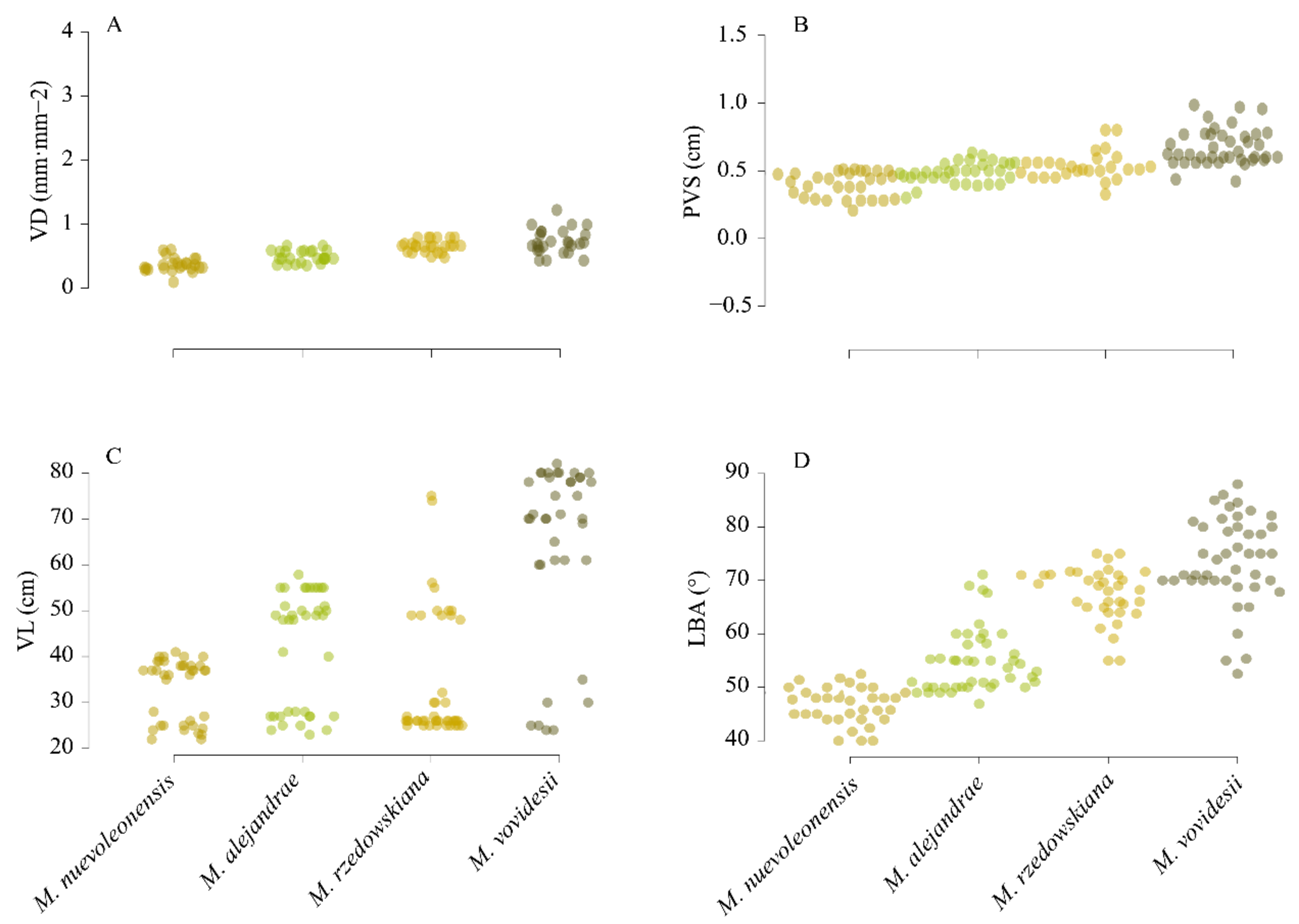
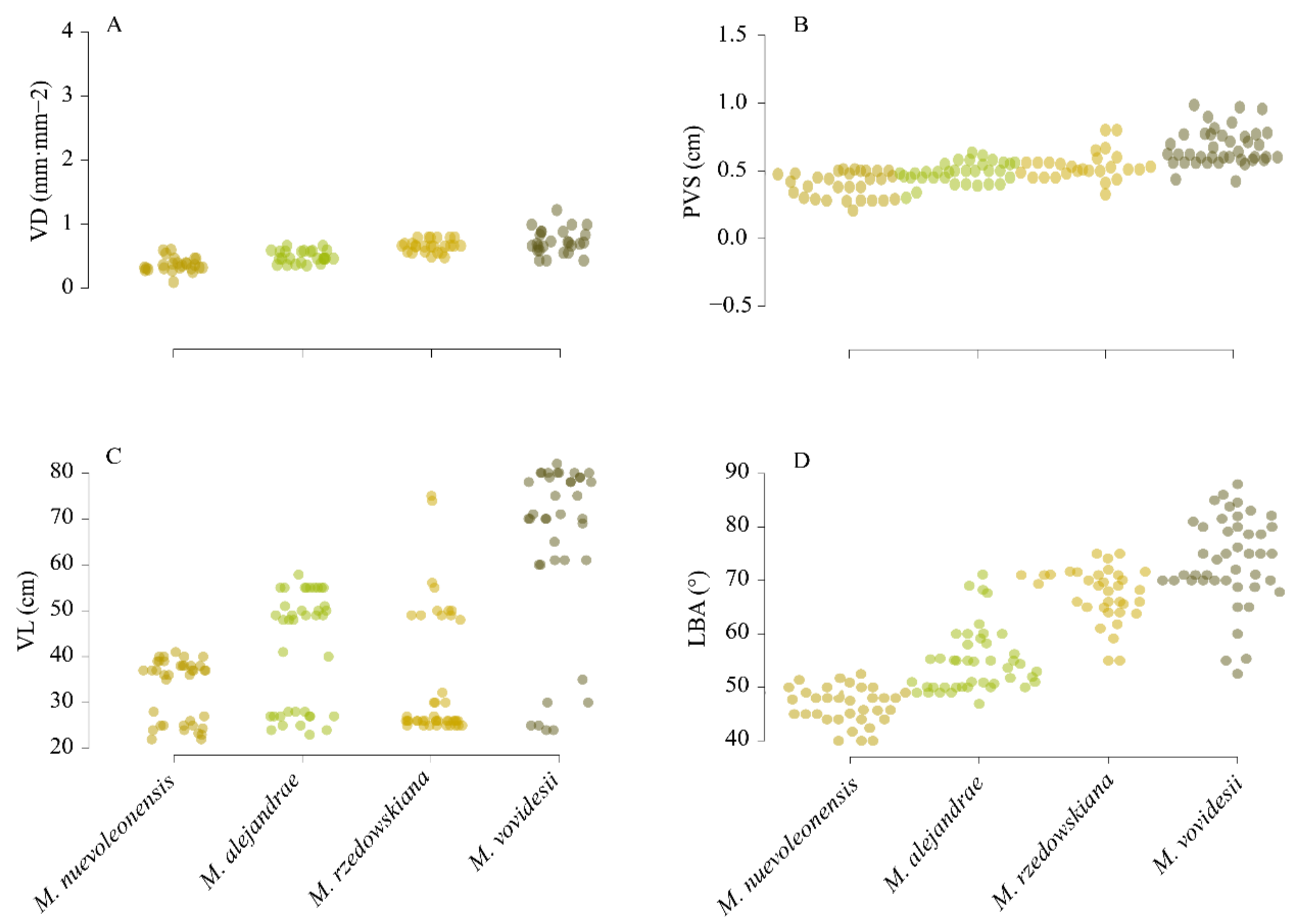
3.1. Leaf Vein Traits
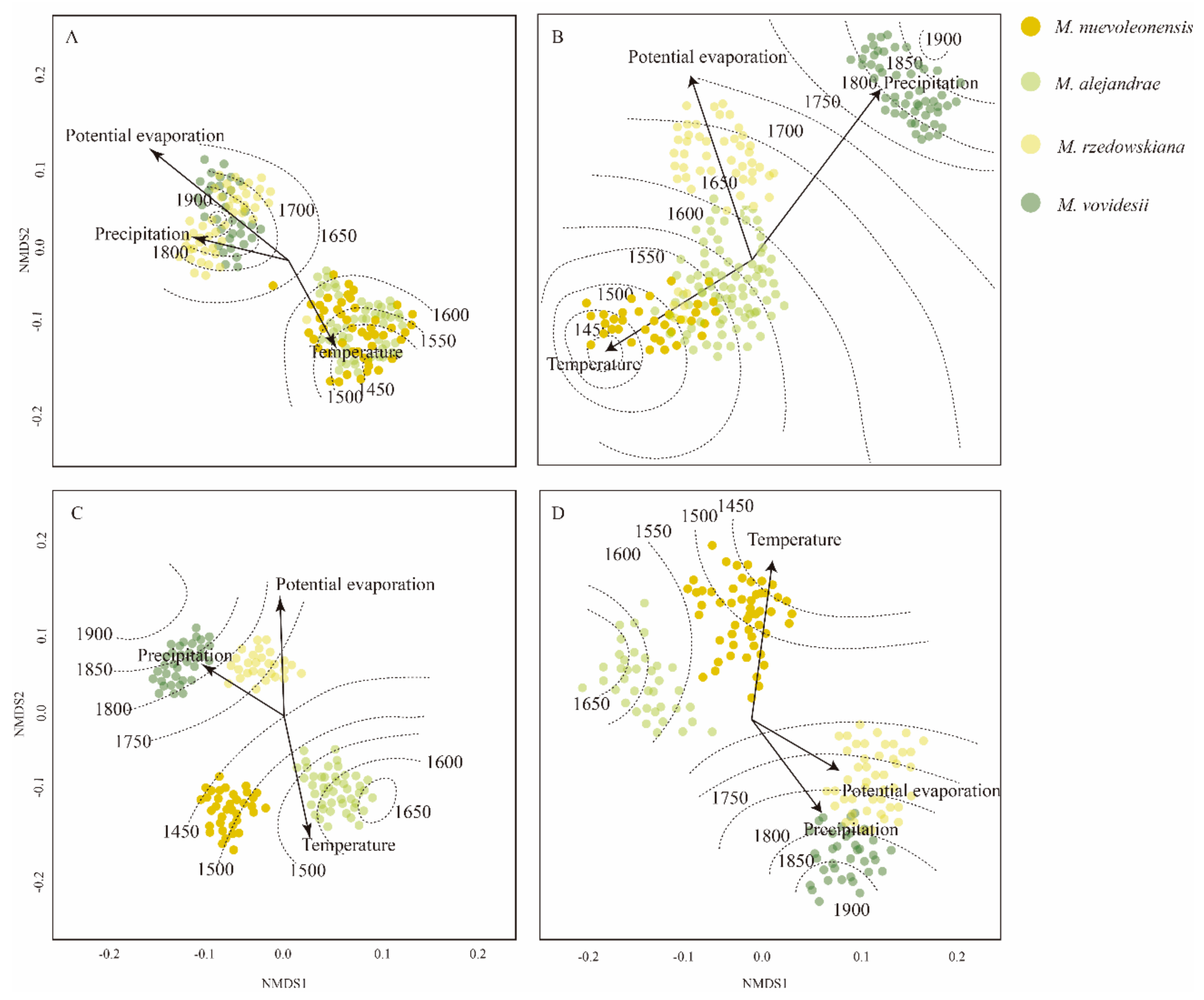
3.2. Leaf Vein Traits and Climatic Factors
4. Discussions and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams-Linera, G. Leaf demography and leaf traits of temperate-deciduous and tropical evergreen-broadleaved trees in a Mexican montane cloud forest. Plant Ecol. 2000, 149, 233–244. [Google Scholar] [CrossRef]
- Rodríguez-Ramírez, E.C.; Vázquez-García, J.A.; García-González, I.; Alcántara-Ayala, O.; Luna-Vega, I. Drought effects on the plasticity in vessel traits of two endemic Magnolia species in the tropical montane cloud forests of eastern Mexico. J. Plant Ecol. 2020, 13, 331–340. [Google Scholar] [CrossRef]
- Blonder, B.; Enquist, B.J. Inferring climate from angiosperm leaf venation networks. New Phytol. 2014, 204, 116–126. [Google Scholar] [CrossRef]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Global climatic drivers of leaf size. Science 2017, 357, 917–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doughty, C.E.; Santos-Andrade, P.E.; Shenkin, A.; Goldsmith, G.R.; Bentley, L.P.; Blonder, B.; Díaz, S.; Salinas, N.; Enquist, B.J.; Martin, R.E.; et al. Tropical forest leaves may darken in response to climate change. Nat. Ecol. Evol. 2018, 2, 1918–1924. [Google Scholar] [CrossRef]
- Liancourt, P.; Boldgiv, B.; Song, D.S.; Spence, L.A.; Helliker, B.R.; Petraitis, P.S.; Casper, B.B. Leaf-trait plasticity and species vulnerability to climate change in a Mongolian steppe. Glob. Chang. Biol. 2015, 21, 3489–3498. [Google Scholar] [CrossRef]
- Ackerly, D.D.; Knight, C.A.; Weiss, S.B.; Barton, K.; Starmer, K.P. Leaf size, specific leaf area and microhabitat distribution of chaparral woody plants: Contrasting patterns in species level and community level analyses. Oecologia 2002, 130, 449–457. [Google Scholar] [CrossRef]
- Sevik, H.; Cetin, M.; Ozel, H.B.; Erbek, A.; Zeren Cetin, I. The effect of climate on leaf micromorphological characteristics in some broad-leaved species. Environ. Dev. Sustain. 2021, 23, 6395–6407. [Google Scholar] [CrossRef]
- Guerin, G.R.; Wen, H.; Lowe, A.J. Leaf morphology shift linked to climate change. Biol. Lett. 2012, 8, 882–886. [Google Scholar] [CrossRef]
- Helmer, E.H.; Gerson, E.A.; Scott Baggett, L.; Bird, B.J.; Ruzycki, T.S.; Voggesser, S.M. Neotropical cloud forests and páramo to contract and dry from declines in cloud immersion and frost. PLoS ONE 2019, 14, e0213155. [Google Scholar] [CrossRef] [Green Version]
- Pound, M.J.; Haywood, A.M.; Salzmann, U.; Riding, J.B.; Lunt, D.J.; Hunter, S.J. A Tortonian (Late Miocene, 11.61–7.25 Ma) global vegetation reconstruction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 300, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Hampe, A. Climatic relicts: Past, present, future. Annu. Rev. Ecol. Evol. Syst. 2011, 42. [Google Scholar] [CrossRef] [Green Version]
- Eller, C.B.; Lima, A.L.; Oliveira, R.S. Cloud forest trees with higher foliar water uptake capacity and anisohydric behavior are more vulnerable to drought and climate change. New Phytol. 2016, 211, 489–501. [Google Scholar] [CrossRef] [Green Version]
- Jiménez-García, D.; Peterson, A.T. Climate change impact on endangered cloud forest tree species in Mexico. Rev. Mex. Biodivers. 2019. [Google Scholar] [CrossRef]
- Vázquez-García, J.A.; Domínguez-Yescas, R.; Velazco-Macías, C.; Shalisko, V.; Merino-Santi, R.E. Magnolia nuevoleonensis sp. nov. (Magnoliaceae) from northeastern Mexico and a key to species of section Macrophylla. Nord. J. Bot. 2016, 34, 48–53. [Google Scholar] [CrossRef]
- Rivers, M.; Beech, E.; Murphy, L.; Oldfield, S. The Red List of Magnoliaceae—Revised and Extended, 1st ed.; Rivers, M., Beech, E., Murphy, L., Oldfield, S., Eds.; Botanic Gardens Conservation International: Richmond, UK, 2016; ISBN 9781905164646. [Google Scholar]
- García-Morales, L.J.; Iamonico, D.; García Jiménez, J. Nomenclatural remarks on Magnolia sect. Macrophylla (Magnoliaceae), with description of a new species from North America (Tamaulipas, Mexico). Phytotaxa 2017, 309, 238–244. [Google Scholar] [CrossRef]
- Vázquez-García, J.A.; Domínguez-Yescas, R.; Pedraza-Ruiz, R.; Sánchez-González, A.; Muñiz-Castro, M.Á. Magnolia rzedowskiana (Magnoliaceae), una especie nueva de la sección Macrophylla de la parte central de la Sierra Madre Oriental, México. Acta Botánica Mex. 2015, 112, 19–36. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ramírez, E.C.; Luna-Vega, I. Dendroecology as a research tool to investigate climate change resilience on Magnolia vovidesii, a threatened Mexican cloud forest tree species of eastern Mexico. In Latin American Dendroecology: Combining Tree-Ring Sciences and Ecology in a Megadiverse Territory; Pompa-García, M., Camarero, J.J., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 3–20. [Google Scholar]
- Ishii, H.R.; Cavaleri, M.A. Canopy ecophysiology: Exploring the terrestrial ecosystem frontier. Tree Physiol. 2017, 37, 1263–1268. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Okuhara, I. Comparison of climatic effects on radial growth of evergreen broad-leaved trees at their northern distribution limit and co-dominating deciduous broad-leaved trees and evergreen conifers. Ecol. Res. 2012, 27, 125–132. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, J.; Huang, Y.; Jia, Z.; Fang, Y. Leaf venation variation and phenotypic plasticity in response to environmental heterogeneity in Parrotia subaequalis (H. T. Chang) R. M. Hao et H. T. Wei, An endemic and endangered tree species from China. Forests 2018, 9, 247. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing (Version 3.4.3); R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011; ISBN 978-1-4419-7975-9. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; Mcglinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package (Version 2.4–1); University of Helsinski: Helsinski, Finland, 2016. [Google Scholar]
- Arlot, S.; Celisse, A. A survey of cross–validation procedures for model selection. Stat. Surv. 2010, 4, 40–79. [Google Scholar] [CrossRef]
- Givnish, T. Leaf and canopy adaptations in tropical forests. In Physiological Ecology of Plants of the Wet Tropics; Medina, E., Mooney, H.A., Vázquez-Yánes, C., Eds.; Springer: Dordrecht, The Netherlands, 1984; pp. 51–84. [Google Scholar]
- Rodríguez-Ramírez, E.C.; Valdez-Nieto, J.A.; Vázquez-García, J.A.; Dieringer, G.; Luna-Vega, I. Plastic responses of Magnolia schiedeana Schltdl., a relict–endangered Mexican cloud forest tree, to climatic events: Evidences from leaf venation and wood vessel anatomy. Forests 2020, 11, 737. [Google Scholar] [CrossRef]
- Gratani, L. Plant phenotypic plasticity in response to environmental factors. Adv. Bot. 2014, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Ramírez, E.C.; Martínez-Mejía, M.d.R.; Ghimire, B.; Luna-Vega, I. Response of leaf morphological traits of relict-endemic Symplocos species (S. coccinea and S. speciosa) to elevation and abiotic fluctuations. Plant Ecol. 2021, 222, 693–704. [Google Scholar] [CrossRef]
- Milla, R.; Reich, P.B. Multi-trait interactions, not phylogeny, fine-tune leaf size reduction with increasing altitude. Ann. Bot. 2011, 107, 455–465. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Ramírez, E.C.; García-Morales, L.J.; Alcántara-Ayala, O.; Vázquez-García, J.A.; Luna-Vega, I. Leaf Vein Morphological Variation in Four Endangered Neotropical Magnolia Species along an Elevation Gradient in the Mexican Tropical Montane Cloud Forests. Plants 2021, 10, 2595. https://doi.org/10.3390/plants10122595
Rodríguez-Ramírez EC, García-Morales LJ, Alcántara-Ayala O, Vázquez-García JA, Luna-Vega I. Leaf Vein Morphological Variation in Four Endangered Neotropical Magnolia Species along an Elevation Gradient in the Mexican Tropical Montane Cloud Forests. Plants. 2021; 10(12):2595. https://doi.org/10.3390/plants10122595
Chicago/Turabian StyleRodríguez-Ramírez, Ernesto C., Leccinum J. García-Morales, Othón Alcántara-Ayala, J. Antonio Vázquez-García, and Isolda Luna-Vega. 2021. "Leaf Vein Morphological Variation in Four Endangered Neotropical Magnolia Species along an Elevation Gradient in the Mexican Tropical Montane Cloud Forests" Plants 10, no. 12: 2595. https://doi.org/10.3390/plants10122595
APA StyleRodríguez-Ramírez, E. C., García-Morales, L. J., Alcántara-Ayala, O., Vázquez-García, J. A., & Luna-Vega, I. (2021). Leaf Vein Morphological Variation in Four Endangered Neotropical Magnolia Species along an Elevation Gradient in the Mexican Tropical Montane Cloud Forests. Plants, 10(12), 2595. https://doi.org/10.3390/plants10122595