
Co-Application of Biochar and Arbuscular mycorrhizal Fungi Improves Salinity Tolerance, Growth and Lipid Metabolism of Maize (Zea mays L.) in an Alkaline Soil


 ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Effect of AMF and Biochar on Fatty Acids Composition of Maize Leaves
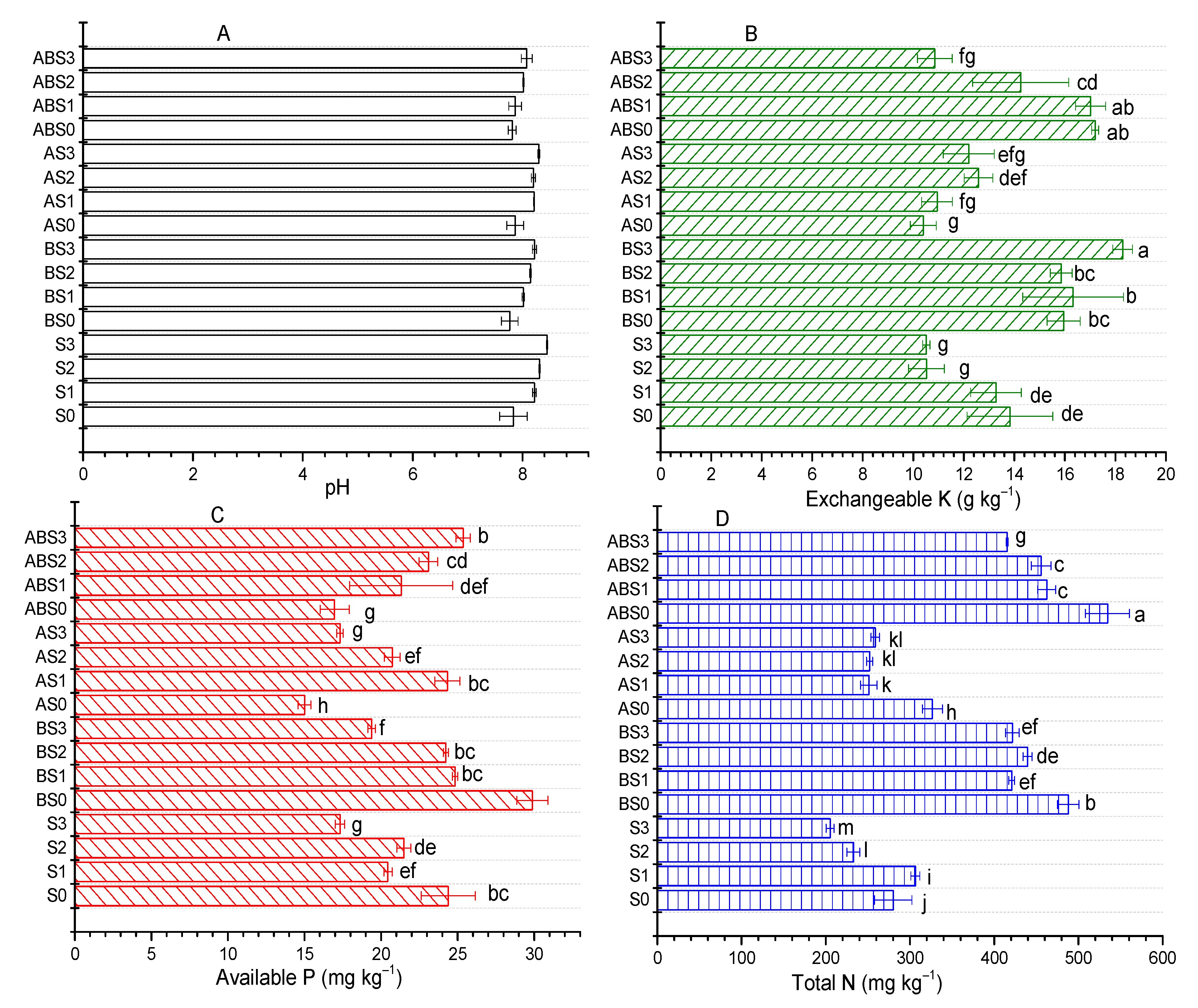
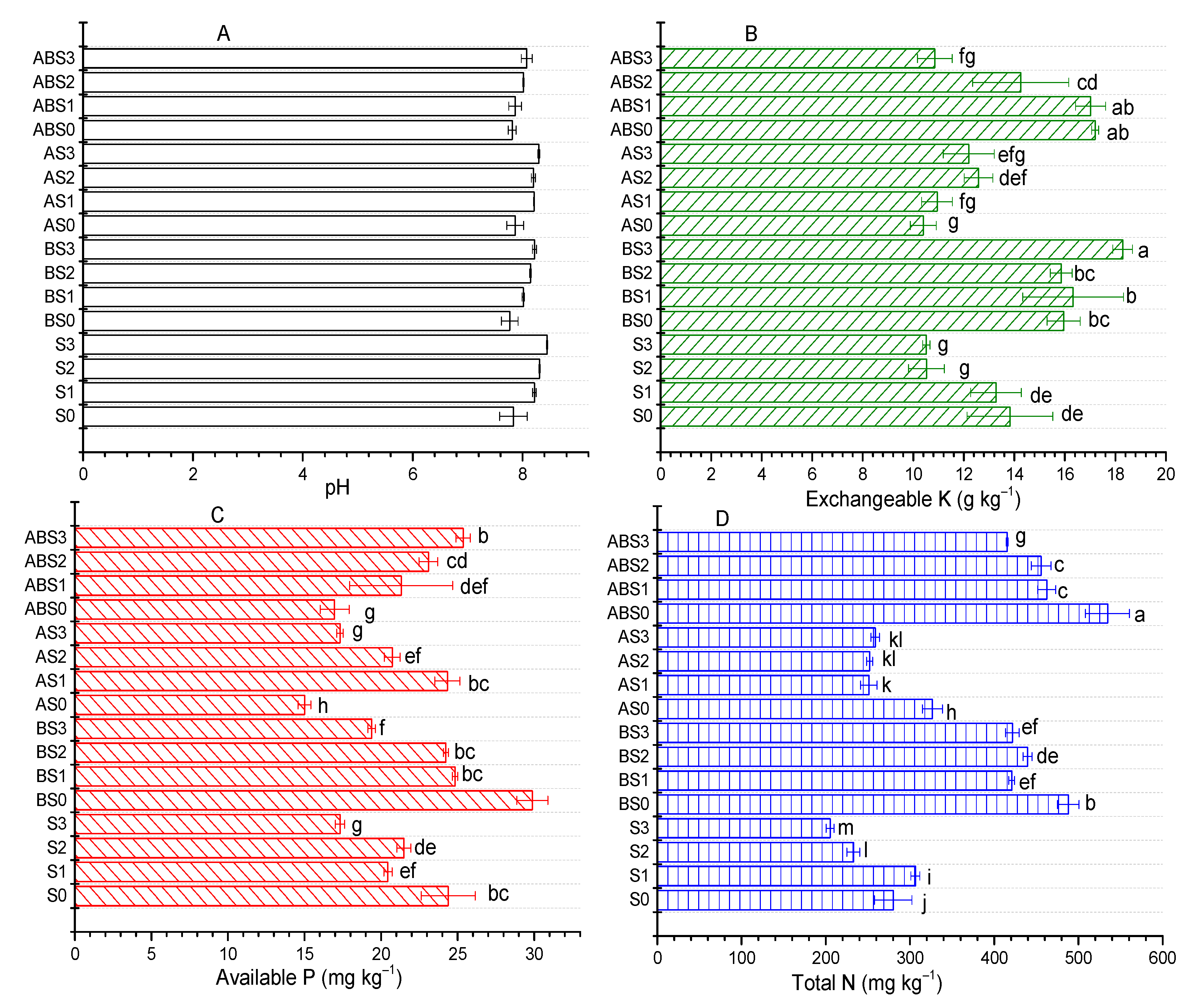
2.2. Influence of AMF and Biochar on Soil Nutrient Content
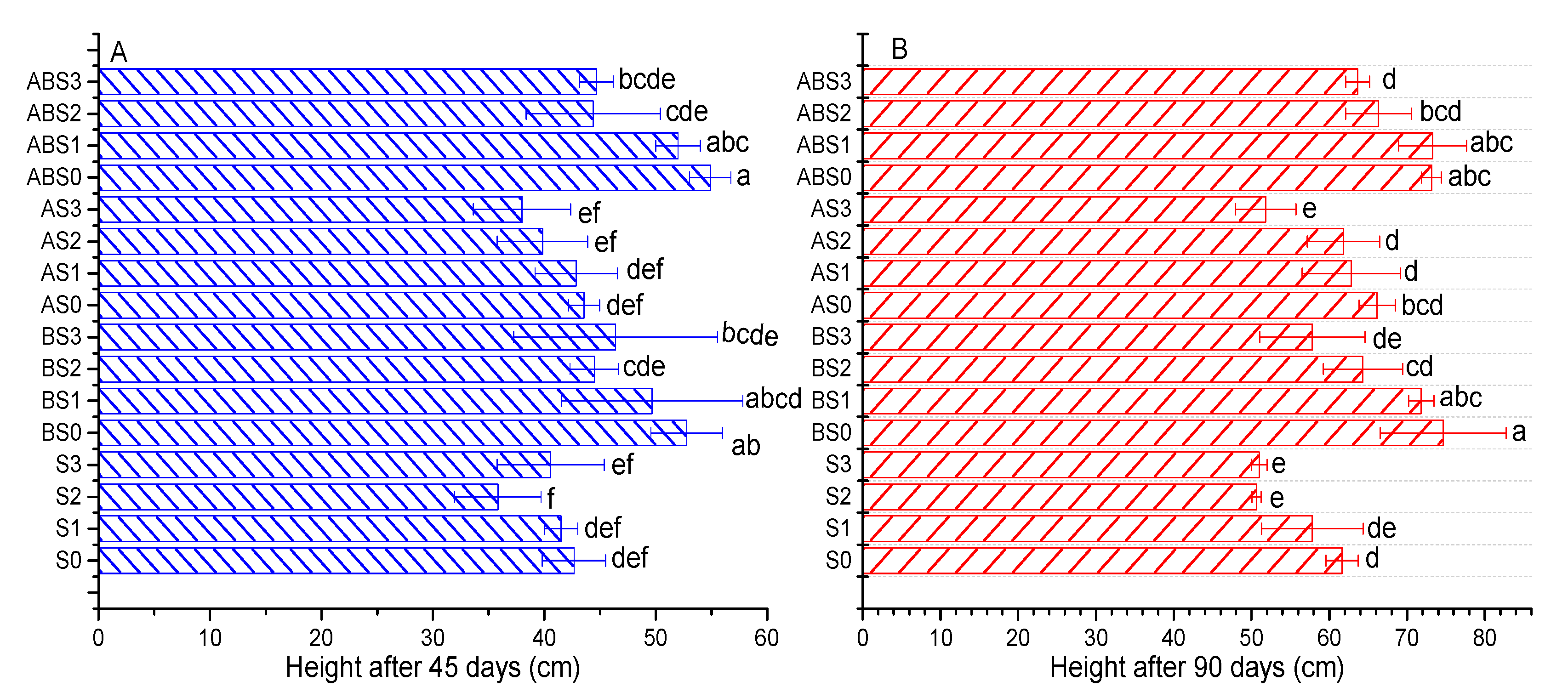
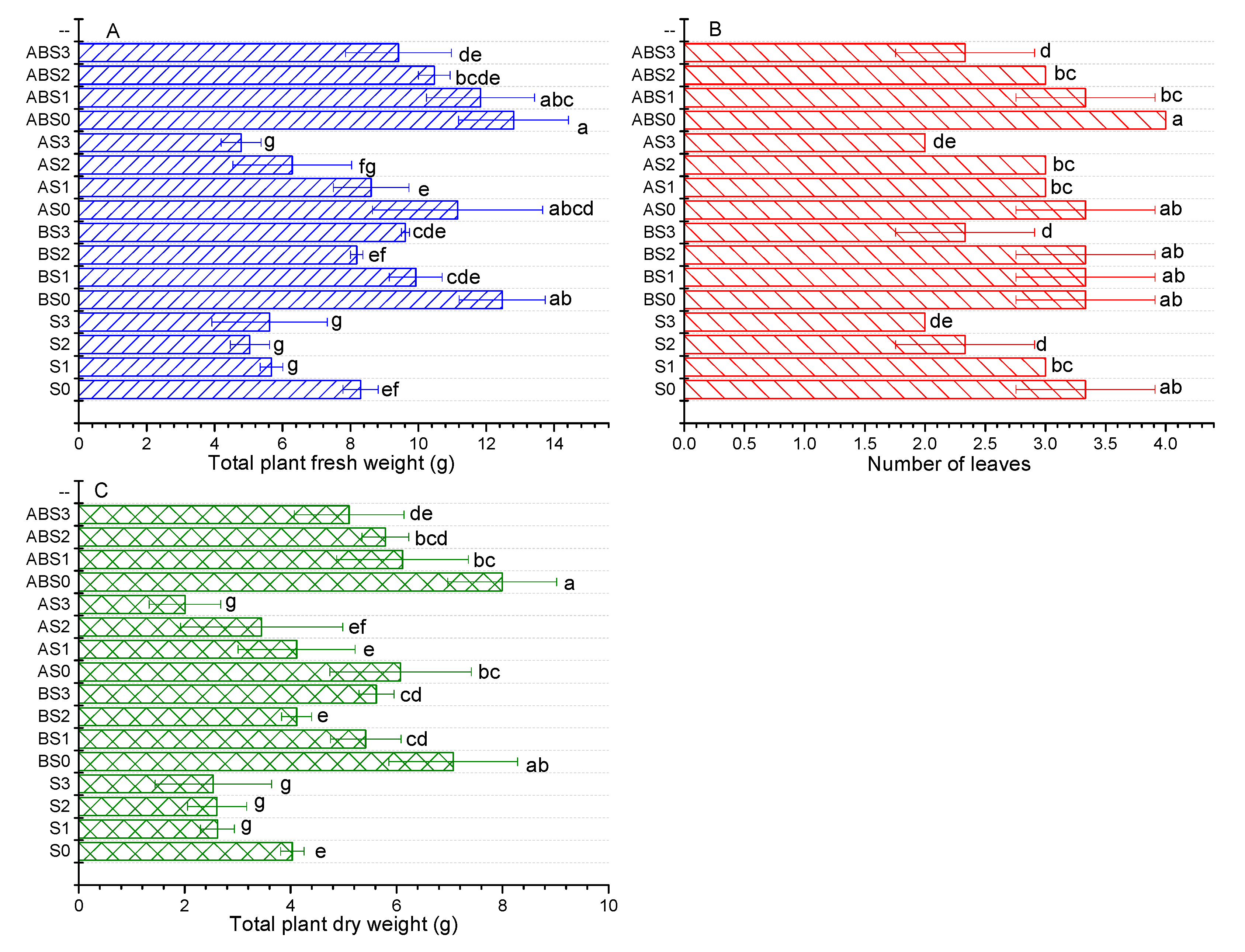
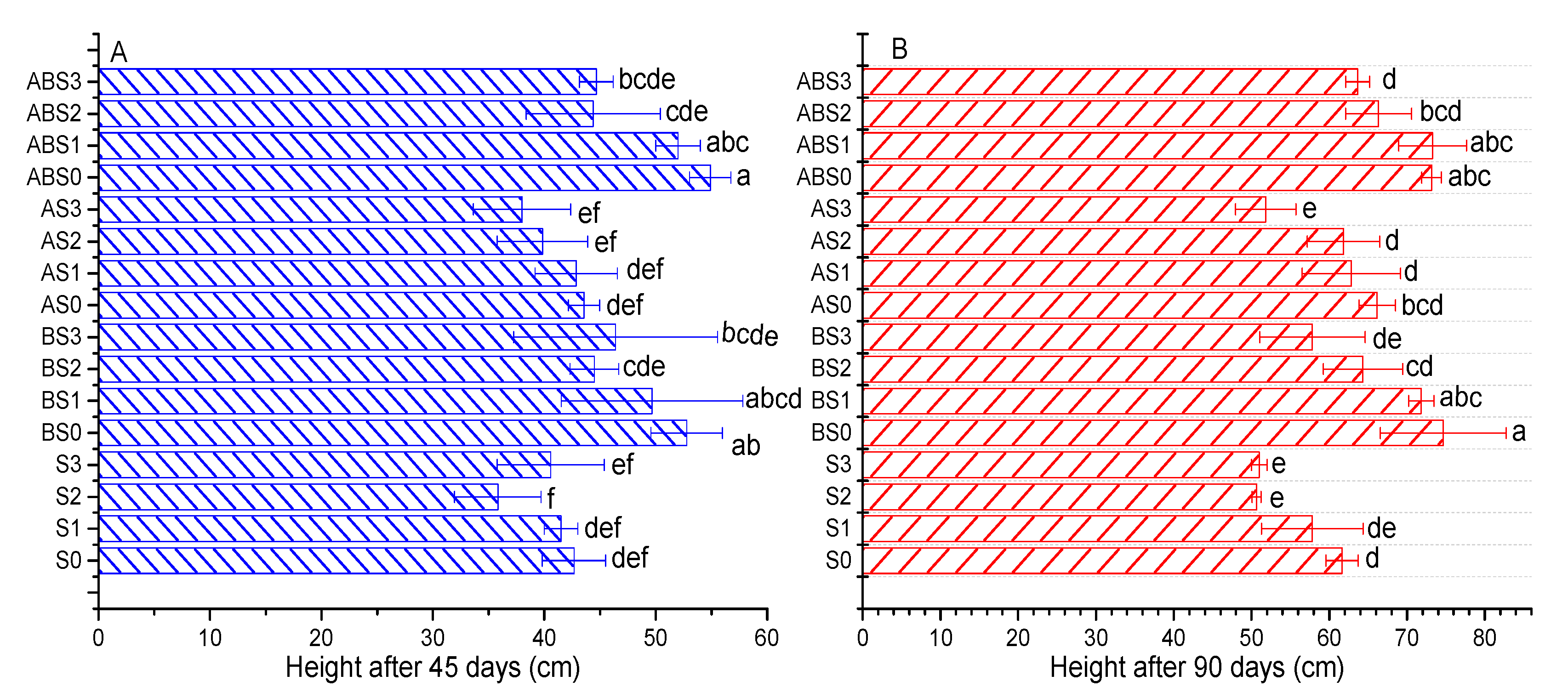
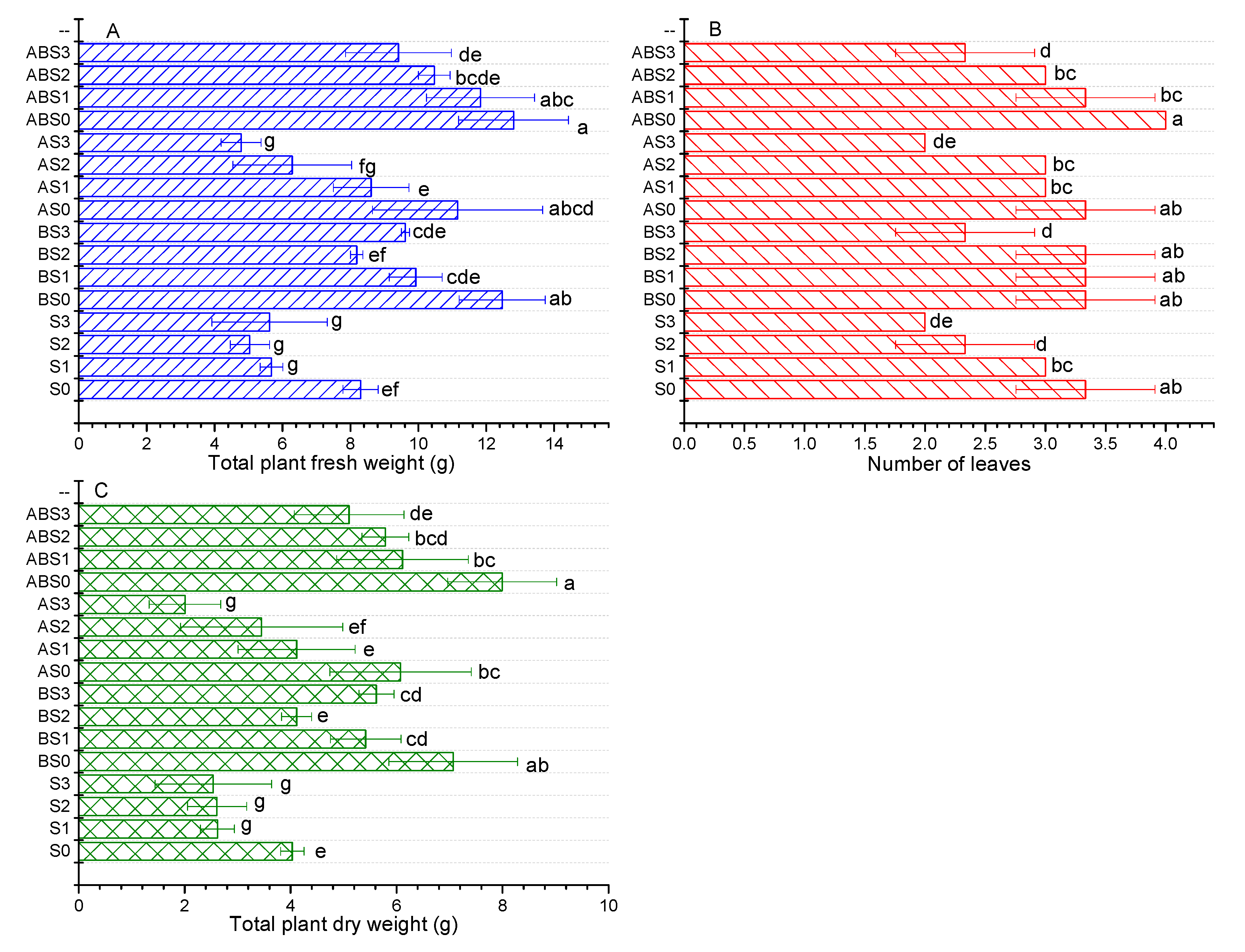
2.3. The Effects of Salinity, Biochar, and AMF on Plant Growth Parameters
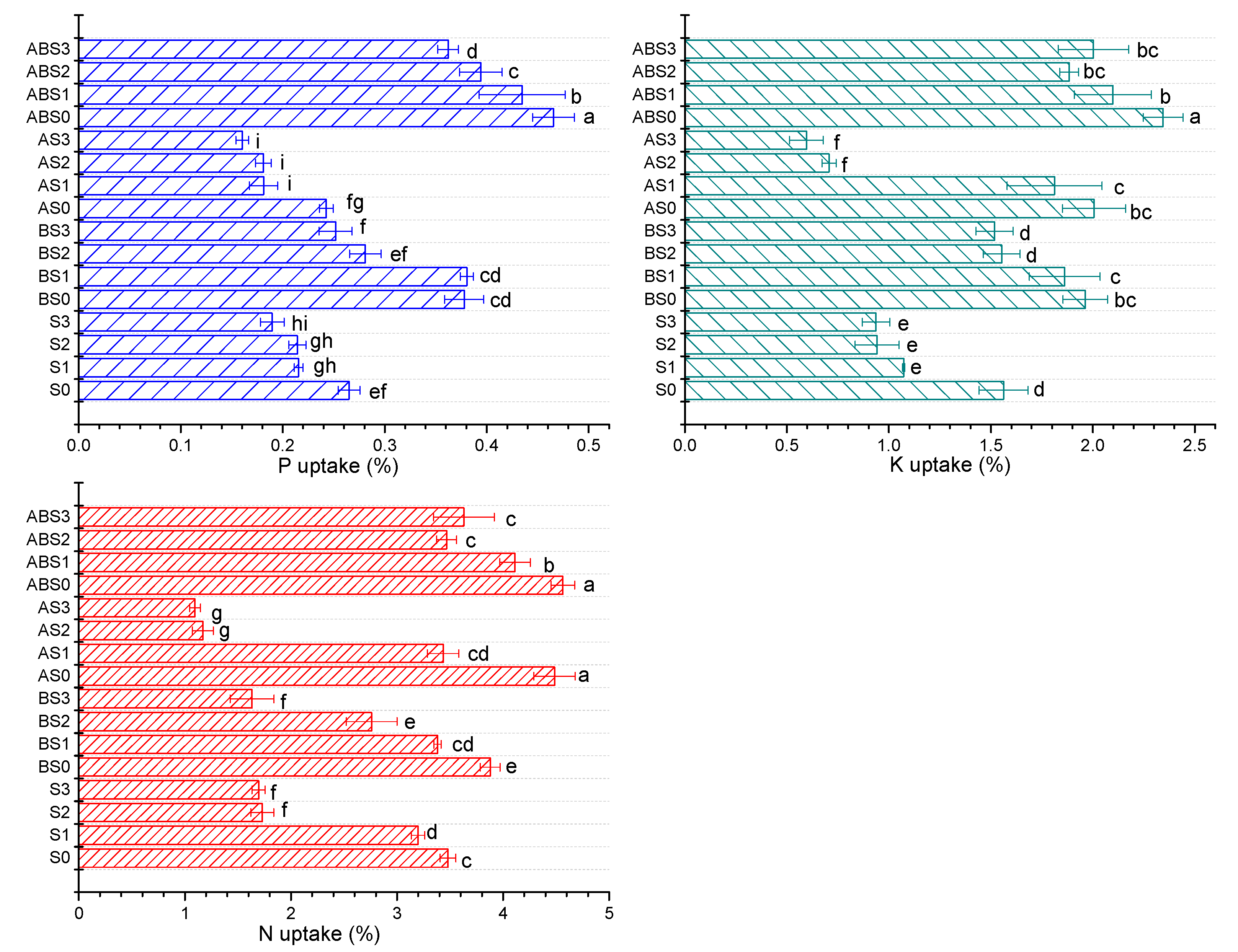
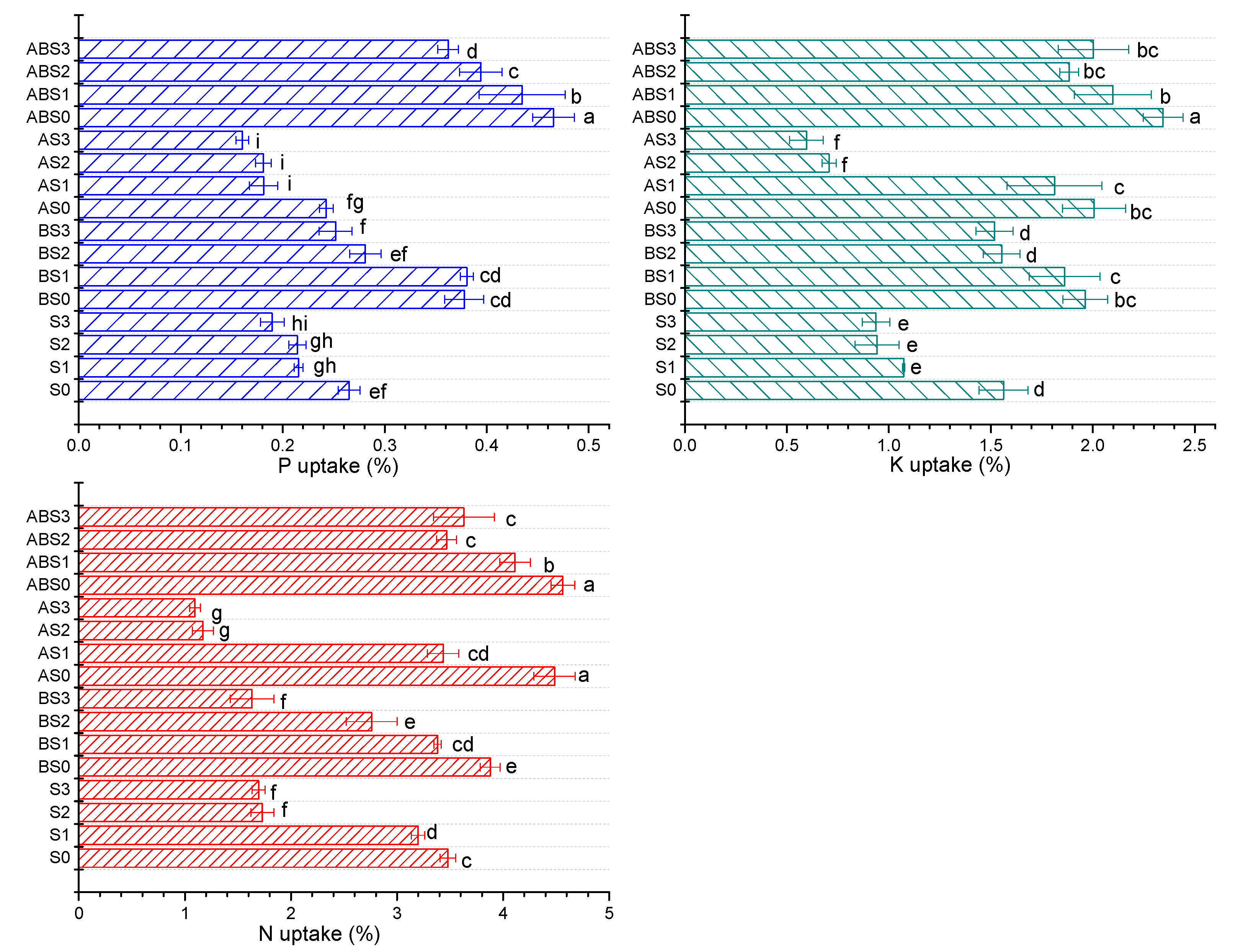
2.4. The Effects of Salinity, Biochar, and AMF on the Nutrient Uptake Ability of the Maize Plant during Growth
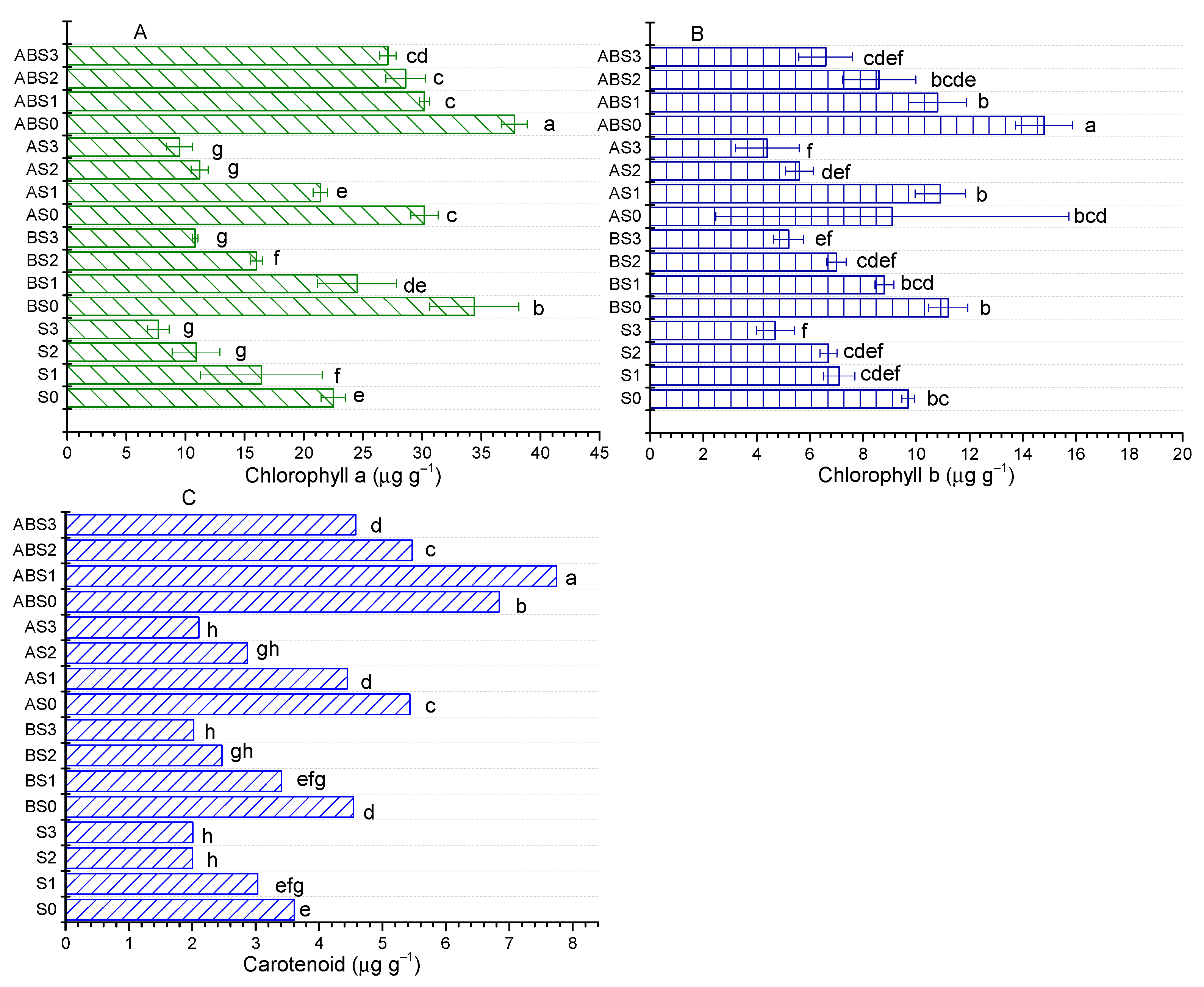
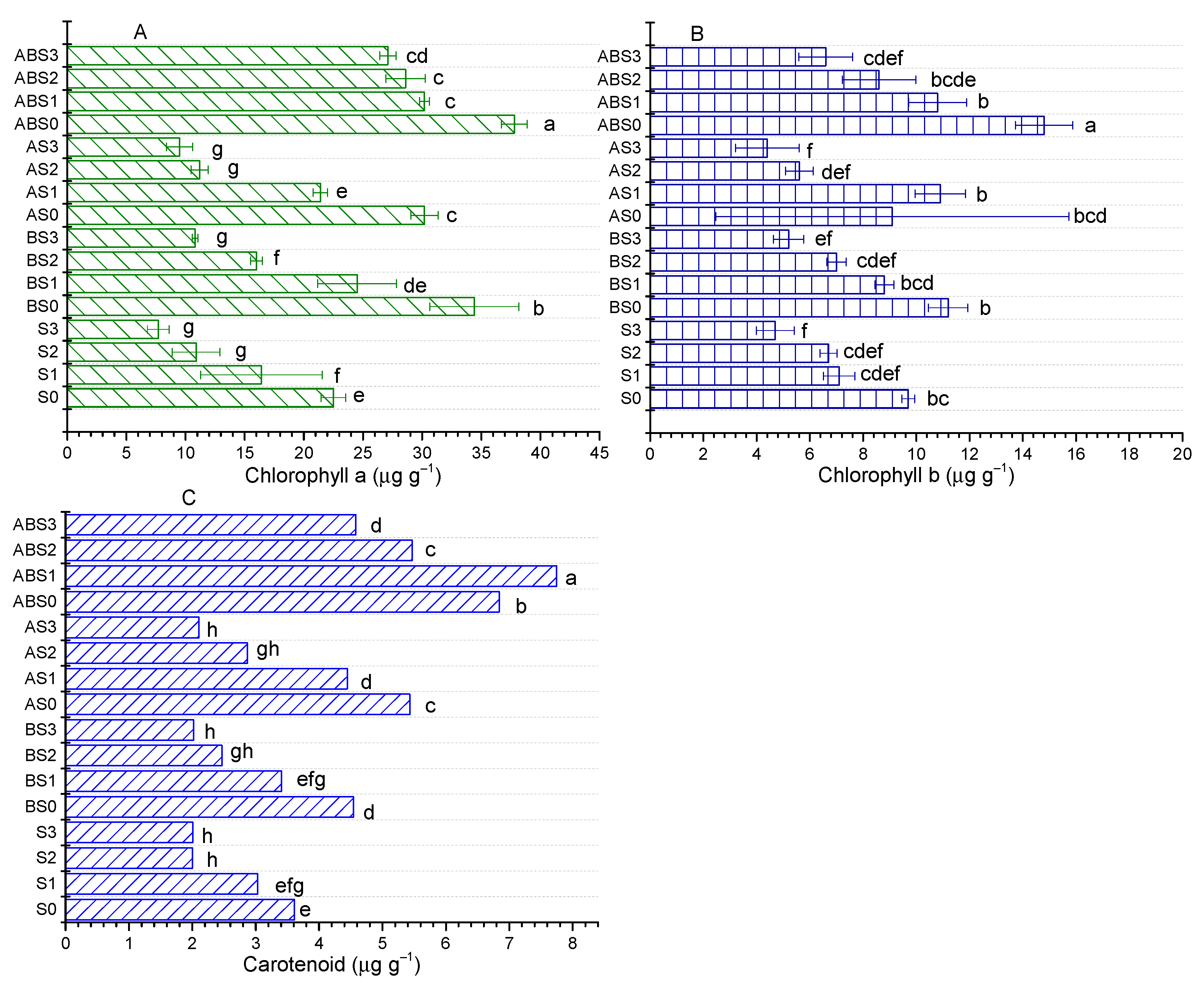
2.5. Influence of AMF and Biochar on Photosynthetic Pigments in Plant
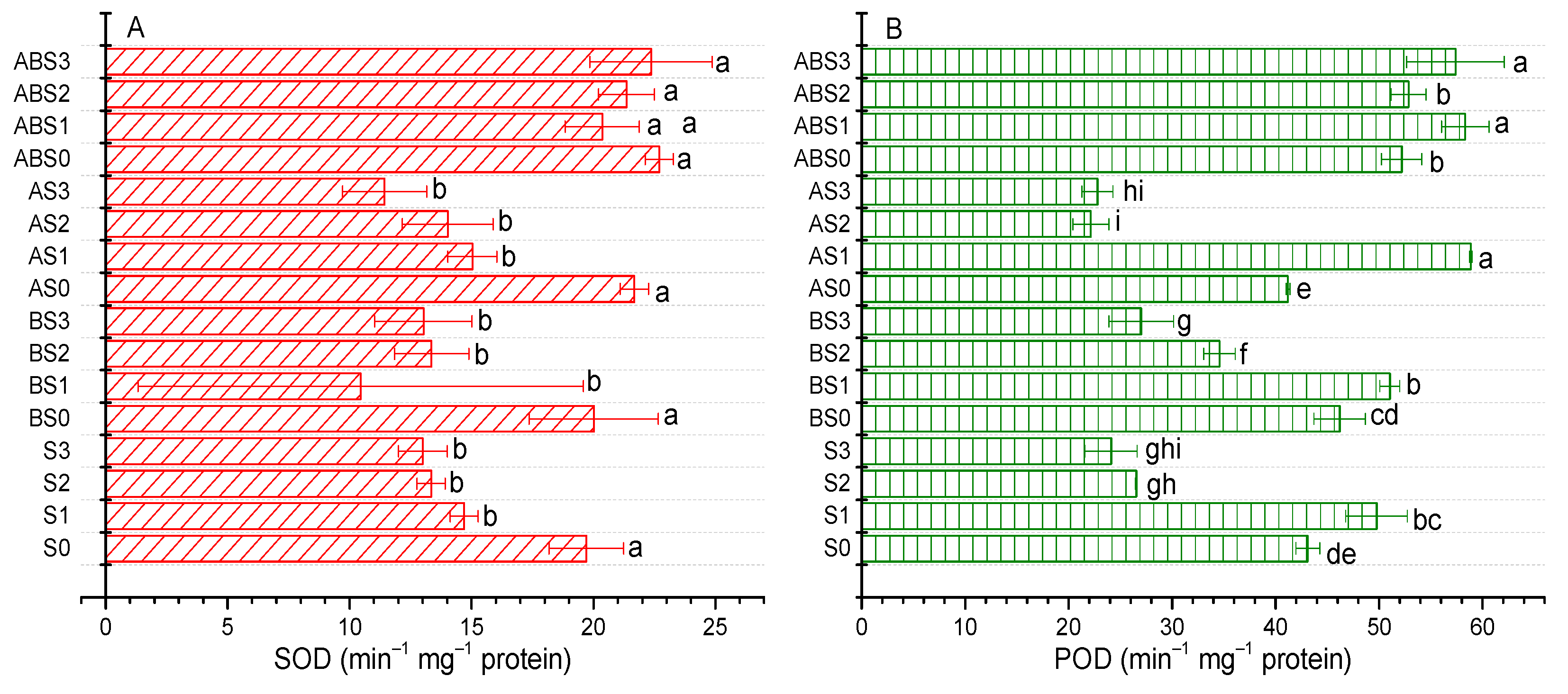
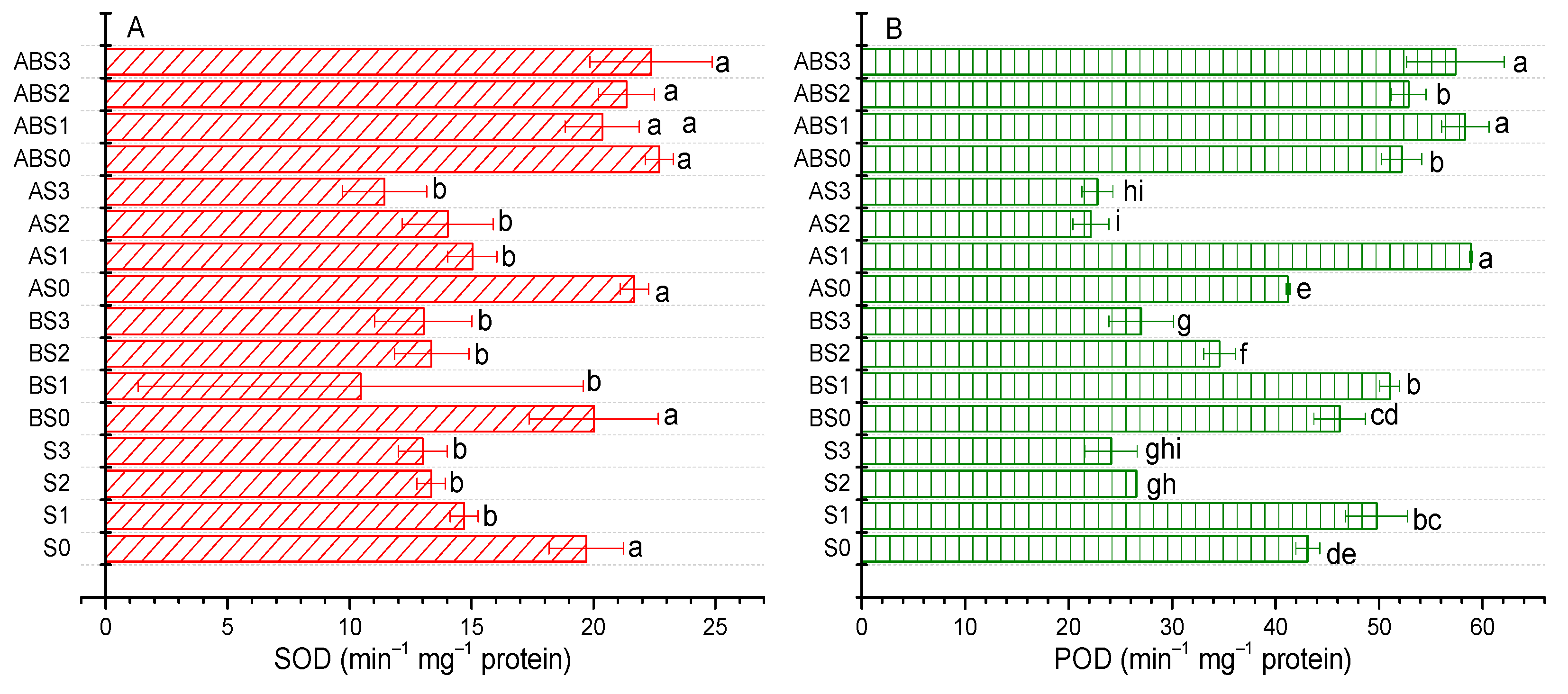
2.6. Impact of AMF and Biochar on Antioxidant Enzyme Activities
3. Discussion
4. Materials and Methods
4.1. Experimental Materials
4.2. Arbuscular Mycorrhizal Fungi (AMF) Inoculum
4.3. Experimental Treatments and Design
4.4. Measurement of Plant Growth Parameters
4.5. Lipid Extraction and Analysis
4.6. Antioxidant Enzyme Activities
4.7. Photosynthetic Pigments
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shi, R.Y.; Hong, Z.N.; Li, J.Y.; Jiang, J.; Baquy, M.A.A.; Xu, R.K.; Qian, W. Mechanisms for Increasing the pH Buffering Capacity of an Acidic Ultisol by Crop Residue-Derived Biochars. J. Agric. Food Chem. 2017, 65, 8111–8119. [Google Scholar] [CrossRef]
- Shi, R.Y.; Hong, Z.N.; Li, J.Y.; Jiang, J.; Kamran, M.A.; Xu, R.K.; Qian, W. Peanut straw biochar increases the resistance of two Ultisols derived from different parent materials to acidification: A mechanism study. J. Environ. Manag. 2018, 210, 171–179. [Google Scholar] [CrossRef]
- Abrishamkesh, S.; Gorji, M.; Asadi, H.; Bagheri-Marandi, G.H.; Pourbabaee, A.A. Effects of rice husk biochar application on the properties of alkaline soil and lentil growth. Plant Soil Environ. 2015, 62, 475–482. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.-Z.; Wang, X.-H.; Du, Z.-L.; Liu, X.-R.; Wang, Y.-D.; Zhang, Q.-Z.; Wang, X.-H.; Du, Z.-L.; Liu, X.-R.; Wang, Y.-D. Impact of biochar on nitrate accumulation in an alkaline soil. Soil Res. 2013, 51, 521–528. [Google Scholar] [CrossRef]
- Baigorri, R.; Francisco, S.S.; Urrutia, Ó.; García-Mina, J.M. Biochar-Ca and Biochar-Al/-Fe-Mediated Phosphate Exchange Capacity are Main Drivers of the Different Biochar Effects on Plants in Acidic and Alkaline Soils. Agronomy 2020, 10, 968. [Google Scholar] [CrossRef]
- Mete, F.Z.; Mia, S.; Dijkstra, F.A.; Abuyusuf, M.; Hossain, A.S.M.I. Synergistic Effects of Biochar and NPK Fertilizer on Soybean Yield in an Alkaline Soil. Pedosphere 2015, 25, 713–719. [Google Scholar] [CrossRef]
- Naveed, M.; Sajid, H.; Mustafa, A.; Niamat, B.; Ahmad, Z.; Yaseen, M.; Kamran, M.; Rafique, M.; Ahmar, S.; Chen, J.T. Alleviation of salinity-induced oxidative stress, improvement in growth, physiology and mineral nutrition of canola (Brassica napus L.) through calcium-fortified composted animal manure. Sustainability 2020, 12, 846. [Google Scholar] [CrossRef] [Green Version]
- Niamat, B.; Naveed, M.; Ahmad, Z.; Yaseen, M.; Ditta, A.; Mustafa, A.; Rafique, M.; Bibi, R.; Sun, N.; Xu, M. Calcium-Enriched Animal Manure Alleviates the Adverse Effects of Salt Stress on Growth, Physiology and Nutrients Homeostasis of Zea mays L. Plants 2019, 8, 480. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signaling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [Green Version]
- Farhangi-Abriz, S.; Torabian, S. Antioxidant enzyme and osmotic adjustment changes in bean seedlings as affected by biochar under salt stress. Ecotoxicol. Environ. Saf. 2017, 137, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Lu, H.; Chu, L.; Shao, H.; Shi, W. Biochar applied with appropriate rates can reduce N leaching, keep N retention and not increase NH3 volatilization in a coastal saline soil. Sci. Total Environ. 2017, 575, 820–825. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, X.; Ma, B.; Chang, S.X.; Gong, J. Biochar addition affected the dynamics of ammonia oxidizers and nitrification in microcosms of a coastal alkaline soil. Biol. Fertil. Soils 2013, 50, 321–332. [Google Scholar] [CrossRef]
- Cui, Q.; Xia, J.; Yang, H.; Liu, J.; Shao, P. Biochar and effective microorganisms promote Sesbania cannabina growth and soil quality in the coastal saline-alkali soil of the Yellow River Delta, China. Sci. Total Environ. 2021, 756, 143801. [Google Scholar] [CrossRef]
- Latef, A.A.H.A.; Chaoxing, H. Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Sci. Hortic. 2011, 127. [Google Scholar] [CrossRef]
- Hashem, A.; Kumar, A.; Al-Dbass, A.M.; Alqarawi, A.A.; Al-Arjani, A.B.F.; Singh, G.; Farooq, M.; Abd_Allah, E.F. Arbuscular mycorrhizal fungi and biochar improves drought tolerance in chickpea. Saudi J. Biol. Sci. 2019, 26, 614–624. [Google Scholar] [CrossRef]
- Yusif, S.A.; Dare, M.O. Effect of Biochar Application and Arbuscular Mycorrhizal Inoculation on Root Colonization and Soil Chemical Properties. Int. Ann. Sci. 2016, 1, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Qadir, M.; Ghafoor, A.; Murtaza, G. Amelioration strategies for saline soils: A review. Land Degrad. Dev. 2000, 11, 501–521. [Google Scholar] [CrossRef]
- Chen, M.; Alim, N.; Zhang, Y.; Xu, N.; Cao, X. Contrasting effects of biochar nanoparticles on the retention and transport of phosphorus in acidic and alkaline soils. Environ. Pollut. 2018, 239, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Abrar, M.M.; Saqib, M.; Abbas, G.; Atiq-ur-Rahman, M.; Mustafa, A.; Shah, S.A.A.; Mehmood, K.; Maitlo, A.A.; Sun, N.; Xu, M. Evaluating the contribution of growth, physiological, and ionic components towards salinity and drought stress tolerance in Jatropha curcas. Plants 2020, 9, 1574. [Google Scholar] [CrossRef]
- Saifullah; Dahlawi, S.; Naeem, A.; Rengel, Z.; Naidu, R. Biochar application for the remediation of salt-affected soils: Challenges and opportunities. Sci. Total Environ. 2018, 625, 320–335. [Google Scholar] [CrossRef]
- Lakhdar, A.; Rabhi, M.; Ghnaya, T.; Montemurro, F.; Jedidi, N.; Abdelly, C. Effectiveness of compost use in salt-affected soil. J. Hazard. Mater. 2009, 171, 29–37. [Google Scholar] [CrossRef]
- Umar, W.; Ayub, M.A.; ur Rehman, M.Z.; Ahmad, H.R.; Farooqi, Z.U.R.; Shahzad, A.; Rehman, U.; Mustafa, A.; Nadeem, M. Nitrogen and Phosphorus Use Efficiency in Agroecosystems. In Resources Use Efficiency in Agriculture; GATEWAY EAST Springer: Singapore, 2020; pp. 213–257. [Google Scholar]
- Gajewska, E.; Skłodowska, M.; Słaba, M.; Mazur, J. Effect of nickel on antioxidative enzyme activities, proline and chlorophyll contents in wheat shoots. Biol. Plant. 2006, 50. [Google Scholar] [CrossRef]
- Sabir, A.; Naveed, M.; Bashir, M.A.; Hussain, A.; Mustafa, A.; Zahir, Z.A.; Kamran, M.; Ditta, A.; Núñez-Delgado, A.; Saeed, Q.; et al. Cadmium mediated phytotoxic impacts in Brassica napus: Managing growth, physiological and oxidative disturbances through combined use of biochar and Enterobacter sp. MN17. J. Environ. Manag. 2020, 265, 110522. [Google Scholar] [CrossRef] [PubMed]
- Kamran, M.; Malik, Z.; Parveen, A.; Zong, Y.; Abbasi, G.H.; Rafiq, M.T.; Shaaban, M.; Mustafa, A.; Bashir, S.; Rafay, M.; et al. Biochar alleviates Cd phytotoxicity by minimizing bioavailability and oxidative stress in pak choi (Brassica chinensis L.) cultivated in Cd-polluted soil. J. Environ. Manag. 2019, 250, 109500. [Google Scholar] [CrossRef] [PubMed]
- Upchurch, R.G. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotechnol. Lett. 2008, 30, 967–977. [Google Scholar] [CrossRef]
- Kachroo, P.; Shanklin, J.; Shah, J.; Whittle, E.J.; Klessig, D.F. A fatty acid desaturase modulates the activation of defense signaling pathways in plants. Proc. Natl. Acad. Sci. USA 2001, 98, 9448–9453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blée, E. Impact of phyto-oxylipins in plant defense. Trends Plant Sci. 2002, 7, 315–322. [Google Scholar] [CrossRef]
- Wilson, R.A.; Calvo, A.M.; Chang, P.K.; Keller, N.P. Characterization of the Aspergillus parasiticus Δ 12-desaturase gene: A role for lipid metabolism in the Aspergillus-seed interaction. Microbiology 2004, 150. [Google Scholar] [CrossRef]
- Xue, H.Q.; Upchurch, R.G.; Kwanyuen, P. Ergosterol as a quantifiable biomass marker for Diaporthe phaseolorum and Cercospora kikuchii. Plant Dis. 2006, 90. [Google Scholar] [CrossRef]
- Dakhma, W.S.; Zarrouk, M.; Cherif, A. Effects of drought-stress on lipids in rape leaves. Phytochemistry 1995, 40, 1383–1386. [Google Scholar] [CrossRef]
- Pál, M.; Leskó, K.; Janda, T.; Páldi, E.; Szalai, G. Cadmium-induced changes in the membrane lipid composition of maize plants. Cereal Res. Commun. 2007, 35, 1631–1642. [Google Scholar] [CrossRef]
- Trinidad, J.L.; Pabuayon, I.C.M.; Kohli, A. Harnessing protein posttranslational modifications for plant improvement. Adv. Crop. Improv. Tech. 2020, 385–401. [Google Scholar] [CrossRef]
- Walkley, A.; Black, D.R. An examination of the digestion method for determination soil organic matter and proposed modification of the chronic acid titration method. Soil Sci. 1935, 37, 29–38. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis Prentice Hall of India. Pvt. Ltd. New Delhi 1967, 151–154, 183–192. [Google Scholar]
- Olsen, S.R.; Sommers, L.E. 1982 Phosphorus; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1973; pp. 403–430. [Google Scholar]
- Pansu, M.; Gautheyrou, J. Handbook of Soil Analysis. Mineralogical, Organic and Inorganic Methods; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2006; p. 993. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total Carbon, Organic Carbon, and Organic Matter. In Methods of Soil Analysis Part 3—Chemical Methods; SSSA Book Series: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Summer, M.E. Methods of Soil Analysis, Parts 2 and 3 Chemical Analysis; Soil Science Society of America Inc.: Madison, WI, USA, 1996. [Google Scholar]
- A.O.A.C. (Association of Official Agricultural Chemists). Official Methods of Analysis, 10th ed.; A.O.A.C.: Washington, WA, USA, 1965; pp. 308–764. [Google Scholar]
- Gerdemann, J.W.; Nicolson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Giovannetti, M.; Mosse, B. An Evaluation of Techniques for Measuring Vesicular Arbuscular Mycorrhizal Infection in Roots. New Phytol. 1980, 84, 489–500. [Google Scholar] [CrossRef]
- Xu, Y.; Siegenthaler, P.A. Low Temperature Treatments Induce an Increase in the Relative Content of Both Linolenic and Δ3-trans-Hexadecenoic Acids in Thylakoid Membrane Phosphatidylglycerol of Squash Cotyledons. Plant Cell Physiol. 1997, 38, 611–618. [Google Scholar] [CrossRef] [Green Version]
- Beauchamp, C.O.; Fridovich, I. Isozymes of superoxide dismutase from wheat germ. BBA Protein Struct. 1973, 317, 50–64. [Google Scholar] [CrossRef]
- Maehly, A.C.; Chance, B. The assay of catalases and peroxidases. Methods Biochem. Anal. 1954, 1, 357–424. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper Enzymes in Isolated Chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.Y.; Haque, M.M.; Molla, A.H.; Rahman, M.M.; Alam, M.Z. Antioxidant compounds and minerals in tomatoes by Trichoderma-enriched biofertilizer and their relationship with the soil environments. J. Integr. Agric. 2017, 16, 691–703. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Fatty Acids (mol %) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| C14:0 | C16:0 | C16:1 | C18:0 | C18:1 | C18:2 | C18:3 | C20:0 | C22:0 | |
| S0 | 1.84 ± 0.138a | 12.7 ± 0.013a | 0.786 ± 0.035bcd | 11.5 ± 0.611a | 10.8 ± 0.017b | 27.4 ± 0.141b | 28.4 ± 0.107b | 2.48 ± 0.103i | 2.29 ± 0.049g |
| S1 | 1.75 ± 0.151a | 12.1 ± 0.216a | 0.701 ± 0.011cdef | 11.2 ± 0.064a | 11.0 ± 1.07b | 26.9 ± 0.06b | 28.1 ± 0.017b | 3.09 ± 0.005g | 3.02 ± 0.002e |
| S2 | 1.81 ± 0.053a | 11.9 ± 0.63a | 0.540 ± 0.059ghi | 11.5 ± 0.236a | 8.92 ± 0.016c | 21.8 ± 0.948cd | 28.5 ± 1.18b | 3.79 ± 0.077de | 3.85 ± 0.15c |
| S3 | 1.90 ± 0.073a | 12.4 ± 0.199a | 0.440 ± 0.033ij | 11.0 ± 1.13a | 7.18 ± 0.556d | 20.3 ± 0.425e | 28.3 ± 0.587b | 3.95 ± 0.055c | 4.3 ± 0.167b |
| BS0 | 1.93 ± 0.056a | 11.5 ± 1.51a | 0.840 ± 0.081bc | 11.7 ± 1.03a | 10.7 ± 0.022b | 27.5 ± 0.578b | 28.8 ± 1.71b | 2.46 ± 0.005i | 2.32 ± 0.006g |
| BS1 | 1.84 ± 0.188a | 11.8 ± 2.18a | 0.640 ± 0.076defgh | 11.0 ± 1.0a | 11.1 ± 1.03b | 27.2 ± 0.647b | 28.0 ± 1.07b | 3.70 ± 0.154e | 2.79 ± 0.056f |
| BS2 | 1.84 ± 0.286a | 12.0 ± 2.81a | 0.560 ± 0.044fghi | 12.2 ± 1.1a | 8.06 ± 1.001cd | 22.2 ± 1.53c | 28.9 ± 1.04b | 4.63 ± 0.017b | 3.54 ± 0.006d |
| BS3 | 1.78 ± 0.161a | 11.7 ± 0.793a | 0.510 ± 0.055hi | 11.7 ± 0.519a | 6.92 ± 1.70d | 20.8 ±0.054de | 28.0 ± 2.31b | 5.66 ± 0.111a | 5.61 ± 0.053a |
| AS0 | 1.83 ± 0.031a | 12.6 ± 0.574a | 0.710 ± 0.071cdef | 11.0 ± 0.049a | 10.8 ± 0.577b | 28.1 ± 0.021b | 28.8 ±0.941b | 2.49 ± 0.053i | 2.29 ± 0.037g |
| AS1 | 1.79 ± 0.172a | 12.2 ± 1.61a | 0.680 ± 0.108defg | 11.1 ± 1.29a | 10.8 ± 0.585b | 27.4 ± 0.584b | 28.7 ± 0.58b | 2.84 ± 0.063h | 2.62 ± 0.05f |
| AS2 | 1.74 ± 0.057a | 12.0 ± 1.47a | 0.630 ± 0.044efgh | 11.5 ± 1.1a | 8.88 ± 0.028c | 22.0 ± 1.217cd | 28.1 ± 0.988b | 2.71 ± 0.0005h | 2.78 ± 0.052f |
| AS3 | 1.69 ± 0.112a | 12.3 ± 1.14a | 0.330 ± 0.249j | 11.7 ± 1.77a | 6.72 ± 0.527d | 20.7 ± 0.051de | 28.3 ± 0.498b | 2.79 ± 0.059h | 2.67 ± 0.302f |
| ABS0 | 1.84 ± 0.063a | 11.9 ± 1.41a | 0.760 ± 0.039cde | 11.9 ± 1.24a | 10.6 ± 0.06b | 27.9 ± 0.554b | 28.2 ± 1.00b | 2.42 ± 0.025i | 2.26 ± 0.051g |
| ABS1 | 1.90 ± 0.074a | 12.4 ± 0.557a | 0.920 ± 0.03b | 12.0 ± 1.77a | 10.3 ± 0.052b | 27.9 ± 0.101b | 28.6 ± 1.01b | 3.53 ± 0.025f | 2.72 ± 0.296f |
| ABS2 | 1.94 ± 0.042a | 12.2 ± 0.641a | 1.16 ± 0.001a | 11.9 ± 1.24a | 14.4 ± 1.16a | 33.9 ± 1.52a | 28.9 ± 0.973b | 3.91 ± 0.051cd | 3.043 ± 0.058e |
| ABS3 | 1.91 ± 0.184a | 12.1 ± 1.15a | 1.15 ± 0.01a | 11.2 ± 1.02a | 14.5 ± 0.579a | 33.5 ± 0.57a | 31.7 ± 1.53a | 4.67 ± 0.194b | 3.56 ± 0.034d |
| Code | Description |
|---|---|
| S0 | 0 mM NaCl without soil amendment |
| S1 | 50 mM NaCl without soil amendment |
| S2 | 100 mM NaCl without soil amendment |
| S3 | 150 mM NaCl without soil amendment |
| BS0 | biochar without NaCl and AMF |
| BS1 | biochar and 50 mM NaCl without AMF |
| BS2 | biochar and 100 mM NaCl without AMF |
| BS3 | biochar and 150 mM NaCl without AMF |
| AS0 | AMF inoculation without NaCl and biochar |
| AS1 | AMF inoculation with 50 mM NaCl but without biochar |
| AS2 | AMF inoculation with 100 mM NaCl but without biochar |
| AS3 | AMF inoculation with 150 mM NaCl but without biochar |
| ABS0 | combined AMF inoculation and biochar but without NaCl |
| ABS1 | combined AMF inoculation, biochar and 50 mM NaCl |
| ABS2 | combined AMF inoculation, biochar and 100 mM NaCl |
| ABS3 | combined AMF inoculation, biochar and 150 mM NaCl |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndiate, N.I.; Saeed, Q.; Haider, F.U.; Liqun, C.; Nkoh, J.N.; Mustafa, A. Co-Application of Biochar and Arbuscular mycorrhizal Fungi Improves Salinity Tolerance, Growth and Lipid Metabolism of Maize (Zea mays L.) in an Alkaline Soil. Plants 2021, 10, 2490. https://doi.org/10.3390/plants10112490
Ndiate NI, Saeed Q, Haider FU, Liqun C, Nkoh JN, Mustafa A. Co-Application of Biochar and Arbuscular mycorrhizal Fungi Improves Salinity Tolerance, Growth and Lipid Metabolism of Maize (Zea mays L.) in an Alkaline Soil. Plants. 2021; 10(11):2490. https://doi.org/10.3390/plants10112490
Chicago/Turabian StyleNdiate, Ndiaye Ibra, Qudsia Saeed, Fasih Ullah Haider, Cai Liqun, Jackson Nkoh Nkoh, and Adnan Mustafa. 2021. "Co-Application of Biochar and Arbuscular mycorrhizal Fungi Improves Salinity Tolerance, Growth and Lipid Metabolism of Maize (Zea mays L.) in an Alkaline Soil" Plants 10, no. 11: 2490. https://doi.org/10.3390/plants10112490
APA StyleNdiate, N. I., Saeed, Q., Haider, F. U., Liqun, C., Nkoh, J. N., & Mustafa, A. (2021). Co-Application of Biochar and Arbuscular mycorrhizal Fungi Improves Salinity Tolerance, Growth and Lipid Metabolism of Maize (Zea mays L.) in an Alkaline Soil. Plants, 10(11), 2490. https://doi.org/10.3390/plants10112490