
Impacts of Agriculture on the Environment and Soil Microbial Biodiversity

 and
and 
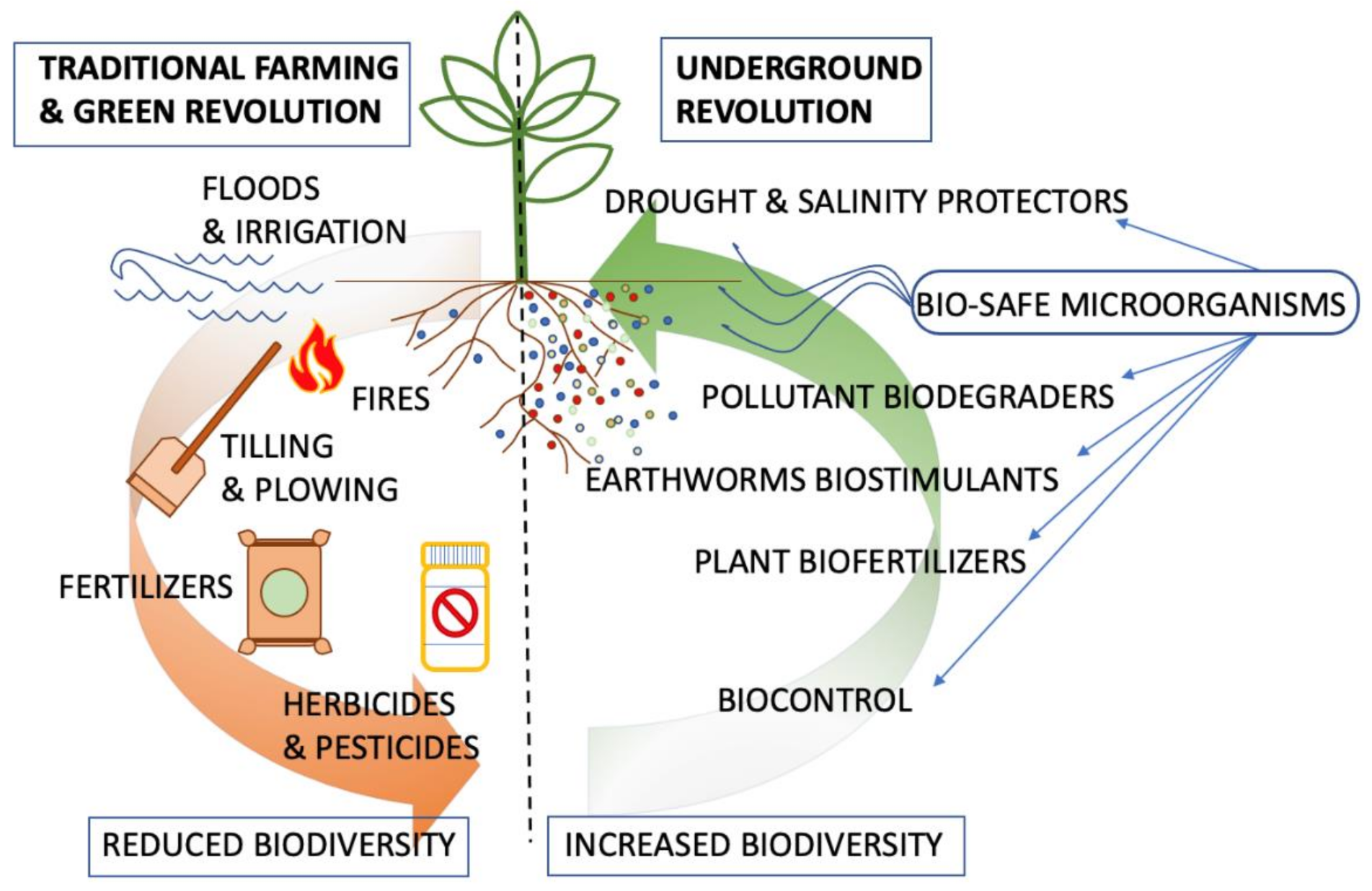
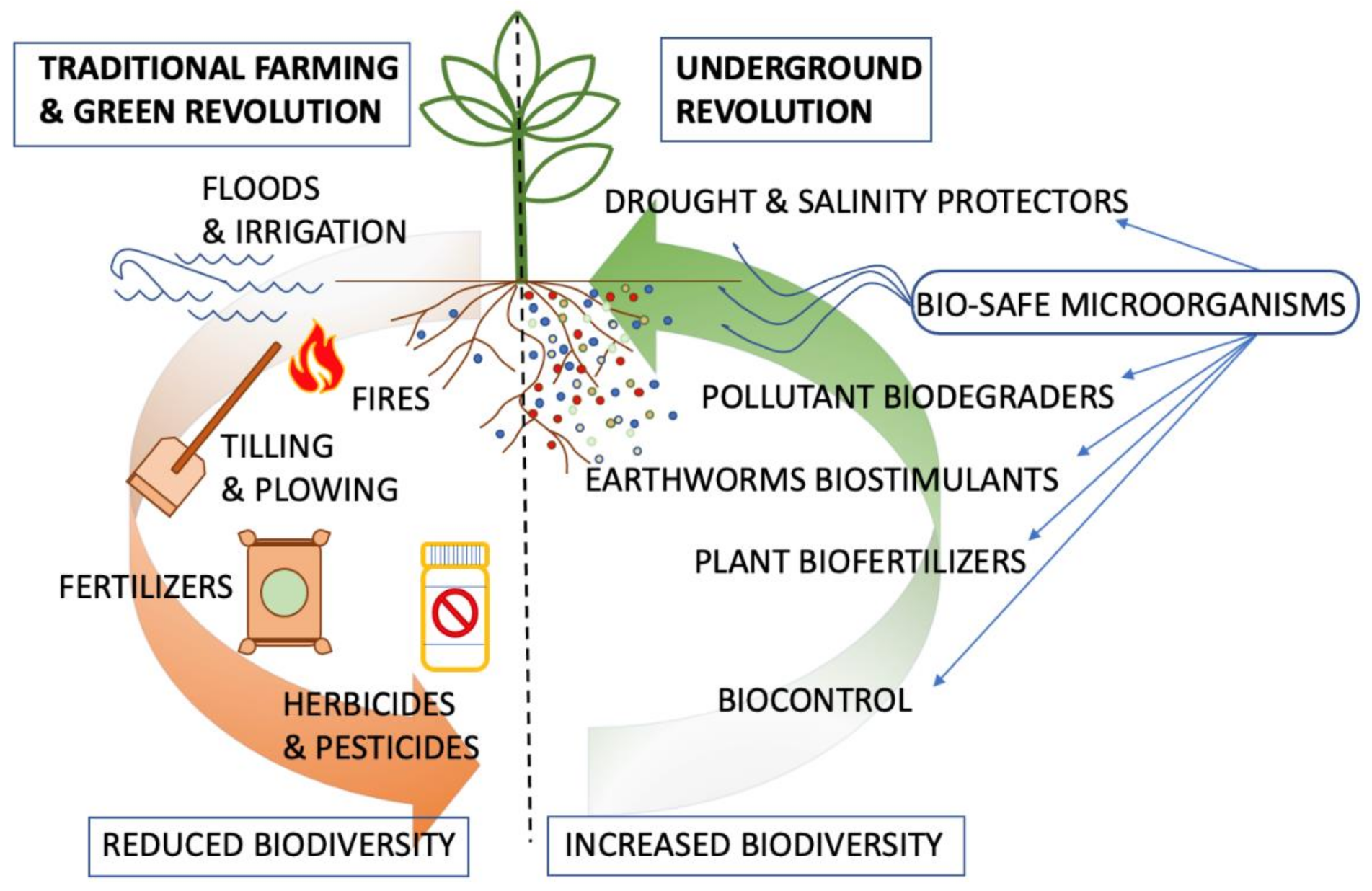{kind=link}
Abstract
:1. Introduction
2. The Impact of Traditional Agricultural Management and Techniques
2.1. Floods
2.2. Irrigation
2.3. Fires
2.4. Tilling and Plowing
2.5. Fertilizers
2.6. Herbicides and Other Pesticides
2.7. Other Aspects of the Green Revolution
3. Future Prospects and Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Johnson, C.N.; Balmford, A.; Brook, B.W.; Buettel, J.C.; Galetti, M.; Guangchun, L.; Wilmshurst, J.M. Biodiversity losses and conservation responses in the Anthropocene. Science 2017, 356, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, M.M.; Araki, H.; Karp, D.S.; Poveda, K.; Whitehead, S.R. The eco-evolutionary impacts of domestication and agricultural practices on wild species. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milla, R.; Osborne, C.P.; Turcotte, M.M.; Violle, C. Plant domestication through an ecological lens. Trends Ecol. Evol. 2015, 30, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Maddison, A. The World Economy. A Millenial Perspective; OECD Publishing: Paris, France, 2001; ISBN 92-64-18998-X. [Google Scholar]
- Nielsen, R.W. Growth of the world population in the past 12,000 years and its link to the economic growth. J. Econ. Bibliogr. 2016, 3, 1–12. [Google Scholar] [CrossRef]
- Ramankutty, N.; Evan, A.T.; Monfreda, C.; Foley, J.A. Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Glob. Biogeochem. Cycles 2008, 22. [Google Scholar] [CrossRef]
- Kehoe, L.; Romero-Muñoz, A.; Polaina, E.; Estes, L.; Kreft, H.; Kuemmerle, T. Biodiversity at risk under future cropland expansion and intensification. Nat. Ecol. Evol. 2017, 1, 1129–1135. [Google Scholar] [CrossRef]
- Best, J. Anthropogenic stresses on the world’s big rivers. Nat. Geosci. 2018, 12, 7–21. [Google Scholar] [CrossRef]
- Macklin, M.G.; Lewin, J. The rivers of civilization. Quat. Sci. Rev. 2015, 114, 228–244. [Google Scholar] [CrossRef]
- González Macé, O.; Steinauer, K.; Jousset, A.; Eisenhauer, N.; Scheu, S. Flood-induced changes in soil microbial functions as modified by plant diversity. PLoS One 2016, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Hefting, M.; Clement, J.-C.; Dowrick, D.; Cosandey, A.; Bernal, S.; Cimpian, C.; Tatur, A.; Burt, T.; Pinay, G. Water table elevation controls on soil nitrogen cycling in riparian wetlands along a European climatic gradient. Biogeochemistry 2004, 67, 113–134. [Google Scholar] [CrossRef]
- Lockaby, B.G.; Murphy, A.L.; Somers, G.L. Hydroperiod Influences on Nutrient Dynamics in Decomposing Litter of a Floodplain Forest. Soil Sci. Soc. Am. J. 1996, 60, 1267–1272. [Google Scholar] [CrossRef]
- Unger, I.M.; Kennedy, A.C.; Muzika, R.-M. Flooding effects on soil microbial communities. Appl. Soil Ecol. 2009, 42, 1–8. [Google Scholar] [CrossRef]
- Wagner, D.; Eisenhauer, N.; Cesarz, S. Plant species richness does not attenuate responses of soil microbial and nematode communities to a flood event. Soil Biol. Biochem. 2015, 89, 135–149. [Google Scholar] [CrossRef]
- Sorensen, P.O.; Germino, M.J.; Feris, K.P. Microbial community responses to 17 years of altered precipitation are seasonally dependent and coupled to co-varying effects of water content on vegetation and soil C. Soil Biol. Biochem. 2013, 64, 155–163. [Google Scholar] [CrossRef]
- Zhang, Y.; Dai, J.; Wang, R.; Zhang, J. Effects of long-term sewage irrigation on agricultural soil microbial structural and functional characterizations in Shandong, China. Eur. J. Soil Biol. 2008, 44, 84–91. [Google Scholar] [CrossRef]
- Whitlock, C.; Colombaroli, D.; Conedera, M.; Tinner, W. Land-use history as a guide for forest conservation and management. Conserv. Biol. 2017, 32, 84–97. [Google Scholar] [CrossRef] [Green Version]
- Potapov, P.; Hansen, M.C.; Laestadius, L.; Turubanova, S.; Yaroshenko, A.; Thies, C.; Smith, W.; Zhuravleva, I.; Komarova, A.; Minnemeyer, S.; et al. The last frontiers of wilderness: Tracking loss of intact forest landscapes from 2000 to 2013. Sci. Adv. 2017, 3, e1600821. [Google Scholar] [CrossRef] [Green Version]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Choromanska, U. Microbial activity and nitrogen mineralization in forest mineral soils following heating: Evaluation of post-fire effects. Soil Biol. Biochem. 2002, 34, 263–271. [Google Scholar] [CrossRef]
- Cobo-Díaz, J.F.; Fernández-González, A.J.; Villadas, P.J.; Robles, A.B.; Toro, N.; Fernández-López, M. Metagenomic assessment of the potential microbial nitrogen pathways in the rhizosphere of a Mediterranean forest after a wildfire. Microb. Ecol. 2015, 69, 895–904. [Google Scholar] [CrossRef]
- Prendergast-Miller, M.T.; de Menezes, A.B.; Macdonald, L.; Toscas, P.; Bissett, A.; Baker, G.; Farrell, M.; Richardson, A.E.; Wark, T.; Thrall, P.H. Wildfire impact: Natural experiment reveals differential short-term changes in soil microbial communities. Soil Biol. Biochem. 2017, 109, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.; Santalahti, M.E.; Pumpanen, J.; Köster, K.; Berninger, F.; Raffaello, T.; Asiegbu, F.O.; Heinonsalo, J. Bacterial community structure and function shift across a northern boreal forest fire chronosequence. Sci. Rep. 2016, 6, 32411. [Google Scholar] [CrossRef] [PubMed]
- Narváez-Reinaldo, J.J.; Barba, I.; González-López, J.; Tunnacliffe, A.; Manzanera, M. Rapid Method for Isolation of Desiccation-Tolerant Strains and Xeroprotectants. Appl. Environ. Microbiol. 2010, 76, 5254–5262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malhi, S.; Kutcher, H. Small grains stubble burning and tillage effects on soil organic C and N, and aggregation in northeastern Saskatchewan. Soil Tillage Res. 2007, 94, 353–361. [Google Scholar] [CrossRef]
- Romasanta, R.R.; Sander, B.O.; Gaihre, Y.K.; Alberto, M.C.; Gummert, M.; Quilty, J.; Nguyen, H.; Castalone, A.G.; Balingbing, C.; Sandro, J.; et al. How does burning of rice straw affect CH4 and N2O emissions? A comparative experiment of different on-field straw management practices. Agric. Ecosyst. Environ. 2017, 239, 143–153. [Google Scholar] [CrossRef]
- Raheem, A.; Sajid, M.; Iqbal, M.S.; Aslam, H.; Bilal, M.; Rafiq, F. Microbial inhabitants of agricultural land have potential to promote plant growth but they are liable to traditional practice of wheat (T. aestivum L) straw burning. Biocatal. Agric. Biotechnol. 2019, 18, 101060. [Google Scholar] [CrossRef]
- Jangid, K.; Williams, M.A.; Franzluebbers, A.; Sanderlin, J.S.; Reeves, J.H.; Jenkins, M.B.; Endale, D.M.; Coleman, D.; Whitman, W. Relative impacts of land-use, management intensity and fertilization upon soil microbial community structure in agricultural systems. Soil Biol. Biochem. 2008, 40, 2843–2853. [Google Scholar] [CrossRef]
- Lal, R. Enhancing ecosystem services with no-till. Renew. Agric. Food Syst. 2013, 28, 102–114. [Google Scholar] [CrossRef]
- Groenigen, J.W.; Lubbers, I.M.; Vos, H.M.J.; Brown, G.G.; De Deyn, G.B.; van Groenigen, K.J. Earthworms increase plant production: A meta-analysis. Sci. Rep. 2016, 4, 6365. [Google Scholar] [CrossRef] [Green Version]
- Scheu, S. Effects of earthworms on plant growth: Patterns and perspectives: The 7th international symposium on earthworm ecology · Cardiff · Wales · 2002. Pedobiologia 2003, 47, 846–856. [Google Scholar] [CrossRef]
- Blouin, M.; Hodson, M.; Delgado, E.A.; Baker, G.L.; Brussaard, L.; Butt, K.; Dai, J.; Dendooven, L.; Peres, G.; Tondoh, J.E.; et al. A review of earthworm impact on soil function and ecosystem services. Eur. J. Soil Sci. 2013, 64, 161–182. [Google Scholar] [CrossRef]
- Groenigen, J.W.V.; Groenigen, K.J.V.; Koopmans, G.F.; Stokkermans, L.; Vos, H.M.J.; Lubbers, I.M. How fertile are earthworm casts? A meta-analysis. Geoderma 2019, 338, 525–535. [Google Scholar] [CrossRef]
- Briones, M.J.I.; Schmidt, O. Conventional tillage decreases the abundance and biomass of earthworms and alters their community structure in a global meta-analysis. Glob. Change Biol. 2017, 23, 4396–4419. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, A.; Bedano, J.C. Earthworm and Enchytraeid Co-occurrence Pattern in Organic and Conventional Farming. Soil Sci. 2016, 181, 148–156. [Google Scholar] [CrossRef]
- Zuber, S.; Villamil, M.B. Meta-analysis approach to assess effect of tillage on microbial biomass and enzyme activities. Soil Biol. Biochem. 2016, 97, 176–187. [Google Scholar] [CrossRef] [Green Version]
- Álvaro-Fuentes, J.; Morell, F.; Madejón, E.; Lampurlanés, J.; Arrúe, J.; Cantero-Martínez, C. Soil biochemical properties in a semiarid Mediterranean agroecosystem as affected by long-term tillage and N fertilization. Soil Tillage Res. 2013, 129, 69–74. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Noya, Y.E.; Gómez-Acata, S.; Montoya-Ciriaco, N.; Rojas-Valdez, A.; Suárez-Arriaga, M.C.; Valenzuela-Encinas, C.; Jiménez-Bueno, N.; Verhulst, N.; Govaerts, B.; Dendooven, L. Relative impacts of tillage, residue management and crop-rotation on soil bacterial communities in a semi-arid agroecosystem. Soil Biol. Biochem. 2013, 65, 86–95. [Google Scholar] [CrossRef]
- Verzeaux, J.; Roger, D.; Lacoux, J.; Nivelle, E.; Adam, C.; Habbib, H.; Hirel, B.; Dubois, F.; Tetu, T. In Winter Wheat, No-Till Increases Mycorrhizal Colonization thus Reducing the Need for Nitrogen Fertilization. Agronomy 2016, 6, 38. [Google Scholar] [CrossRef] [Green Version]
- Clark, C.M.; Tilman, D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature 2008, 451, 712–715. [Google Scholar] [CrossRef]
- Dickson, T.L.; Gross, K.L. Plant community responses to long-term fertilization: Changes in functional group abundance drive changes in species richness. Oecologia 2013, 173, 1513–1520. [Google Scholar] [CrossRef]
- Suding, K.N.; Collins, S.; Gough, L.; Clark, C.; Cleland, E.E.; Gross, K.L.; Milchunas, D.G.; Pennings, S. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proc. Natl. Acad. Sci. USA 2005, 102, 4387–4392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMalach, N.; Zaady, E.; Weiner, J.; Kadmon, R. Size asymmetry of resource competition and the structure of plant communities. J. Ecol. 2016, 104, 899–910. [Google Scholar] [CrossRef]
- Rajaniemi, T.K. Explaining productivity-diversity relationships in plants. Oikos 2003, 101, 449–457. [Google Scholar] [CrossRef]
- Foster, B.L.; Gross, K.L. Species richness in a successional grassland: Effects of nitrogen enrichment and plant litter. Ecology 1998, 79, 2593–2602. [Google Scholar] [CrossRef]
- Midolo, G.; Alkemade, R.; Schipper, A.M.; Benítez-López, A.; Perring, M.P.; De Vries, W. Impacts of nitrogen addition on plant species richness and abundance: A global meta-analysis. Glob. Ecol. Biogeogr. 2018, 28, 398–413. [Google Scholar] [CrossRef] [Green Version]
- Fay, P.A.; Prober, S.M.; Harpole, W.S.; Knops, J.M.H.; Bakker, J.D.; Borer, E.T.; Lind, E.M.; MacDougall, A.S.; Seabloom, E.W.; Wragg, P.D.; et al. Grassland productivity limited by multiple nutrients. Nat. Plants 2015, 1, 15080. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Kumar, U.; Nayak, A.K.; Shahid, M.; Gupta, V.V.; Panneerselvam, P.; Mohanty, S.; Kaviraj, M.; Kumar, A.; Chatterjee, D.; Lal, B.; et al. Continuous application of inorganic and organic fertilizers over 47 years in paddy soil alters the bacterial community structure and its influence on rice production. Agric. Ecosyst. Environ. 2018, 262, 65–75. [Google Scholar] [CrossRef]
- Wang, L.; Yang, F.; Yaoyao, E.; Yuan, J.; Raza, W.; Huang, Q.; Shen, Q. Long-Term Application of Bioorganic Fertilizers Improved Soil Biochemical Properties and Microbial Communities of an Apple Orchard Soil. Front. Microbiol. 2016, 7, 1893. [Google Scholar] [CrossRef]
- Beffa, R.; Menne, H.; Köcher, H. Herbicide Resistance Action Committee (HRAC): Herbicide classification, resistance evolution, survey, and resistance mitigation activities. In Modern Crop Protection Compounds, Volume 3: Insecticides, Third, Completely Revised and Enlarged Edition; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2019; pp. 5–32. [Google Scholar] [CrossRef]
- García-Delgado, C.; Vicente, V.B.; Marín-Benito, J.; Igual, J.M.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S. Influence of different agricultural management practices on soil microbial community over dissipation time of two herbicides. Sci. Total Environ. 2018, 646, 1478–1488. [Google Scholar] [CrossRef]
- Wang, Y.; Hatt, J.K.; Tsementzi, D.; Rodriguez-R, L.M.; Perez, C.A.R.; Weigand, M.R.; Kizer, H.; Maresca, G.; Krishnan, R.; Poretsky, R.; et al. Quantifying the Importance of the Rare Biosphere for Microbial Community Response to Organic Pollutants in a Freshwater Ecosystem. Appl. Environ. Microbiol. 2017, 83, e03321-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallego, S.; Devers-Lamrani, M.; Rousidou, K.; Karpouzas, D.G.; Martin-Laurent, F. Assessment of the effects of oxamyl on the bacterial community of an agricultural soil exhibiting enhanced biodegradation. Sci. Total Environ. 2018, 651, 1189–1198. [Google Scholar] [CrossRef]
- Mulla, S.I.; Ameen, F.; Talwar, M.P.; Eqani, S.A.M.A.S.; Bharagava, R.N.; Saxena, G.; Tallur, P.N.; Ninnekar, H.Z. Organophosphate pesticides: Impact on environment, toxicity, and their degradation. In Bioremediation of Industrial Waste for Environmental Safety; Springer: Singapore, 2020; pp. 265–290. [Google Scholar] [CrossRef]
- Pingali, P.L. Green Revolution: Impacts, limits, and the path ahead. Proc. Natl. Acad. Sci. USA 2012, 109, 12302–12308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horlings, L.; Marsden, T. Towards the real green revolution? Exploring the conceptual dimensions of a new ecological modernisation of agriculture that could ‘feed the world’. Glob. Environ. Chang. 2011, 21, 441–452. [Google Scholar] [CrossRef]
- Manzanera, M.; Molina-Muñoz, M.L.; González-López, J. Biodiesel: An alternative fuel. Recent Pat. Biotechnol. 2008, 2, 25–34. [Google Scholar] [CrossRef]
- Tilman, D.; Fargione, J.; Wolff, B.; D’Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting Agriculturally Driven Global Environmental Change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [Green Version]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; van der Heijden, M.G.A. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef] [Green Version]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G. An Underground Revolution: Biodiversity and Soil Ecological Engineering for Agricultural Sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y.; Canaguier, R.; Svecova, E.B.; Cardarelli, M. Biostimulant action of a plant-derived protein hydrolysate produced through enzymatic hydrolysis. Front. Plant. Sci. 2014, 5, 448. [Google Scholar] [CrossRef] [Green Version]
- Vílchez, J.I.; Fontana, C.G.; Román-Naranjo, D.; González-López, J.; Manzanera, M. Plant Drought Tolerance Enhancement by Trehalose Production of Desiccation-Tolerant Microorganisms. Front. Microbiol. 2016, 7, 1577. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; De Pascale, S.; Cozzolino, E.; Giordano, M.; El-Nakhel, C.; Cuciniello, A.; Cenvinzo, V.; Colla, G.; Rouphael, Y. Protein Hydrolysate or Plant Extract-based Biostimulants Enhanced Yield and Quality Performances of Greenhouse Perennial Wall Rocket Grown in Different Seasons. Plants 2019, 8, 208. [Google Scholar] [CrossRef] [Green Version]
- Vílchez, J.I.; Navas, A.; Egonzález-López, J.; Arcos, S.C.; Emanzanera, M. Biosafety Test for Plant Growth-Promoting Bacteria: Proposed Environmental and Human Safety Index (EHSI) Protocol. Front. Microbiol. 2016, 6, 1514. [Google Scholar] [CrossRef] [PubMed]
- Bao, X.; Wang, Y.; Olsson, P.A. Arbuscular mycorrhiza under water—Carbon‒phosphorus exchange between rice and arbuscular mycorrhizal fungi under different flooding regimes. Soil Biol. Biochem. 2018, 129, 169–177. [Google Scholar] [CrossRef]
- Zheng, F.-L.; Liang, S.-M.; Chu, X.-N.; Yang, Y.-L.; Wu, Q.-S. Mycorrhizal fungi enhance flooding tolerance of peach through inducing proline accumulation and improving root architecture. Plant. Soil Environ. 2020, 66, 624–631. [Google Scholar] [CrossRef]
- Etesami, H.; Hosseini, H.M.; Alikhani, H.A. Bacterial biosynthesis of 1-aminocyclopropane-1-caboxylate (ACC) deaminase, a useful trait to elongation and endophytic colonization of the roots of rice under constant flooded conditions. Physiol. Mol. Biol. Plants 2014, 20, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Ravanbakhsh, M.; Sasidharan, R.; Voesenek, L.A.C.J.; Kowalchuk, G.A.; Jousset, A. ACC deaminase-producing rhizosphere bacteria modulate plant responses to flooding. J. Ecol. 2017, 105, 979–986. [Google Scholar] [CrossRef]
- Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Mattarozzi, M.; Di Zinno, J.; Montanini, B.; Manfredi, M.; Marengo, E.; Fornasier, F.; Ferrarini, A.; Careri, M.; Visioli, G. Biostimulants applied to maize seeds modulate the enzymatic activity and metaproteome of the rhizosphere. Appl. Soil Ecol. 2020, 148, 103480. [Google Scholar] [CrossRef]
- Turjaman, M.; Osaki, M. The Role of mycorrhizal fungi for supporting aerohydro culture in tropical peatland. In Tropical Peatland Eco-Management; Springer: Singapore, 2021; pp. 285–299. [Google Scholar] [CrossRef]
- Pizarro-Tobias, P.; Fernandez, M.; Niqui, J.L.; Solano, J.; Duque, E.; Ramos, J.; Roca, A. Restoration of a Mediterranean forest after a fire: Bioremediation and rhizoremediation field-scale trial. Microb. Biotechnol. 2014, 8, 77–92. [Google Scholar] [CrossRef]
- Vilchez, S.; Tunnacliffe, A.; Manzanera, M. Tolerance of plastic-encapsulated Pseudomonas putida KT2440 to chemical stress. Extremophiles 2007, 12, 297–299. [Google Scholar] [CrossRef]
- Vílchez, J.I.; Niehaus, K.; Dowling, D.N.; González-López, J.; Manzanera, M. Protection of Pepper Plants from Drought by Microbacterium sp. 3J1 by Modulation of the Plant’s Glutamine and α-ketoglutarate Content: A Comparative Metabolomics Approach. Front. Microbiol. 2018, 9, 284. [Google Scholar] [CrossRef] [PubMed]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Front. Plant Sci. 2019, 10, 845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangseekaew, P.; Barros-Rodríguez, A.; Pathom-Aree, W.; Manzanera, M. Deep-Sea Actinobacteria Mitigate Salinity Stress in Tomato Seedlings and Their Biosafety Testing. Plants 2021, 10, 1687. [Google Scholar] [CrossRef] [PubMed]
- Barros-Rodríguez, A.; Rangseekaew, P.; Lausadee, K.; Pathom-aree, W.; Manzanera, M. Regulatory risks associated with bacteria as biostimulants and biofertilizers in the frame of the European Regulation (EU) 2019/1009. Sci. Total Environ. 2020, 740, 140239. [Google Scholar] [CrossRef]
- Owen, D.; Williams, A.P.; Griffith, G.W.; Withers, P.J.A. Use of commercial bio-inoculants to increase agricultural production through improved phosphorus acquisition. Appl. Soil Ecol. 2015, 86, 41–54. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barros-Rodríguez, A.; Rangseekaew, P.; Lasudee, K.; Pathom-aree, W.; Manzanera, M. Impacts of Agriculture on the Environment and Soil Microbial Biodiversity. Plants 2021, 10, 2325. https://doi.org/10.3390/plants10112325
Barros-Rodríguez A, Rangseekaew P, Lasudee K, Pathom-aree W, Manzanera M. Impacts of Agriculture on the Environment and Soil Microbial Biodiversity. Plants. 2021; 10(11):2325. https://doi.org/10.3390/plants10112325
Chicago/Turabian StyleBarros-Rodríguez, Adoración, Pharada Rangseekaew, Krisana Lasudee, Wasu Pathom-aree, and Maximino Manzanera. 2021. "Impacts of Agriculture on the Environment and Soil Microbial Biodiversity" Plants 10, no. 11: 2325. https://doi.org/10.3390/plants10112325
APA StyleBarros-Rodríguez, A., Rangseekaew, P., Lasudee, K., Pathom-aree, W., & Manzanera, M. (2021). Impacts of Agriculture on the Environment and Soil Microbial Biodiversity. Plants, 10(11), 2325. https://doi.org/10.3390/plants10112325