
Undiscovered Biodiversity of the European Moss Flora: Neodicranella hamulosa (Aongstroemiaceae), a New Genus and Species from SW Portugal



{kind=link}
{kind=link}
{kind=link}
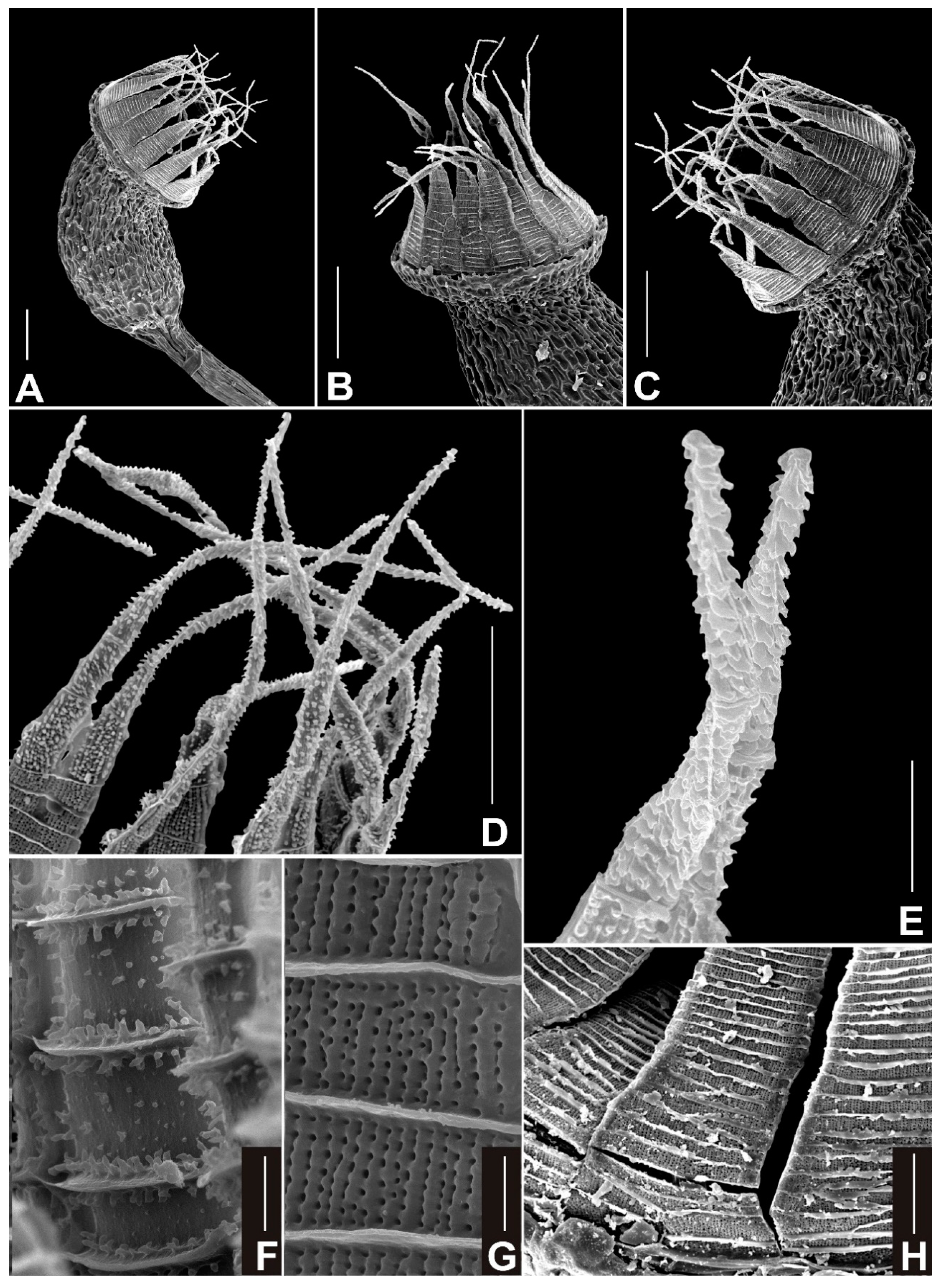{kind=link}
{kind=link}
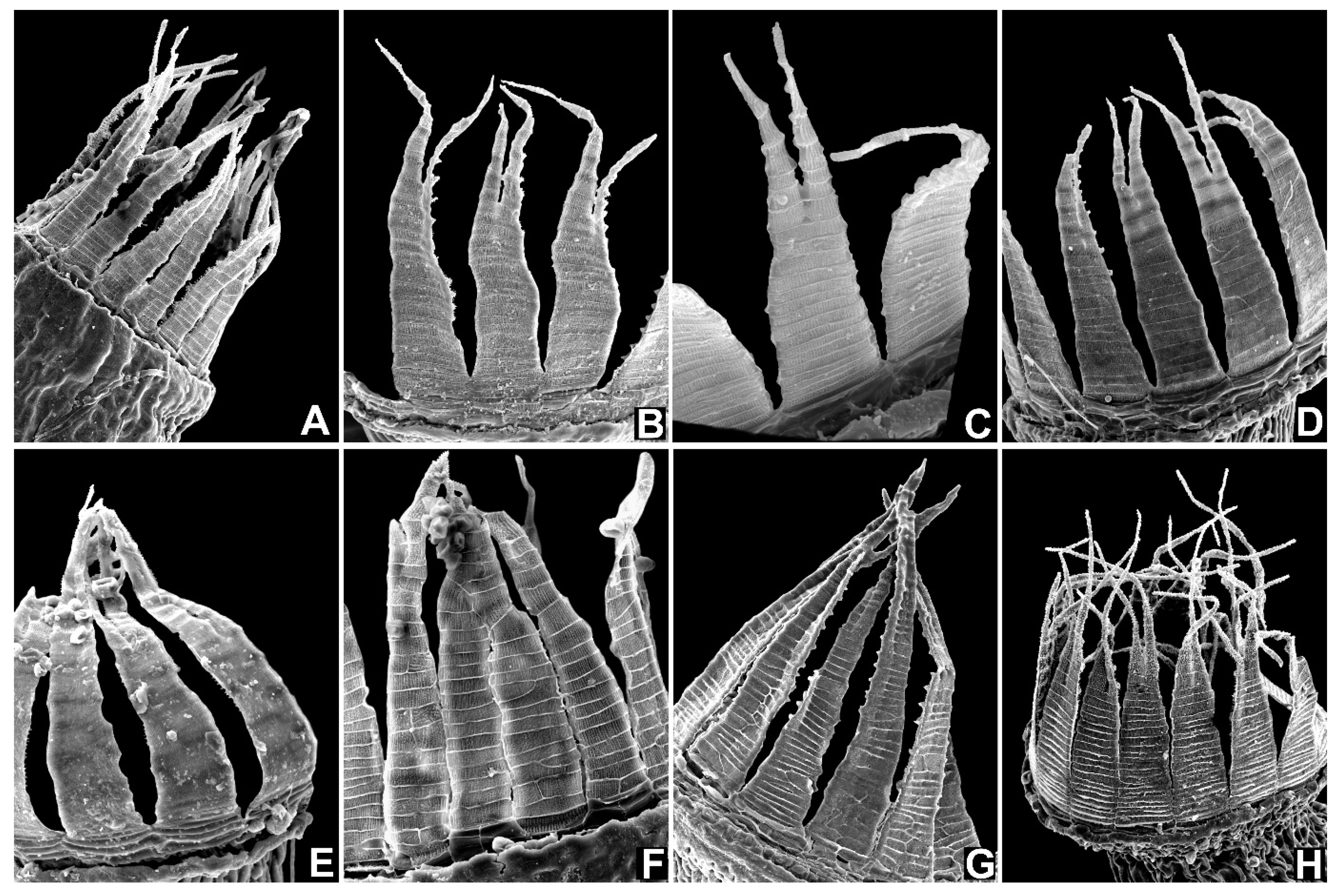{kind=link}
{kind=link}
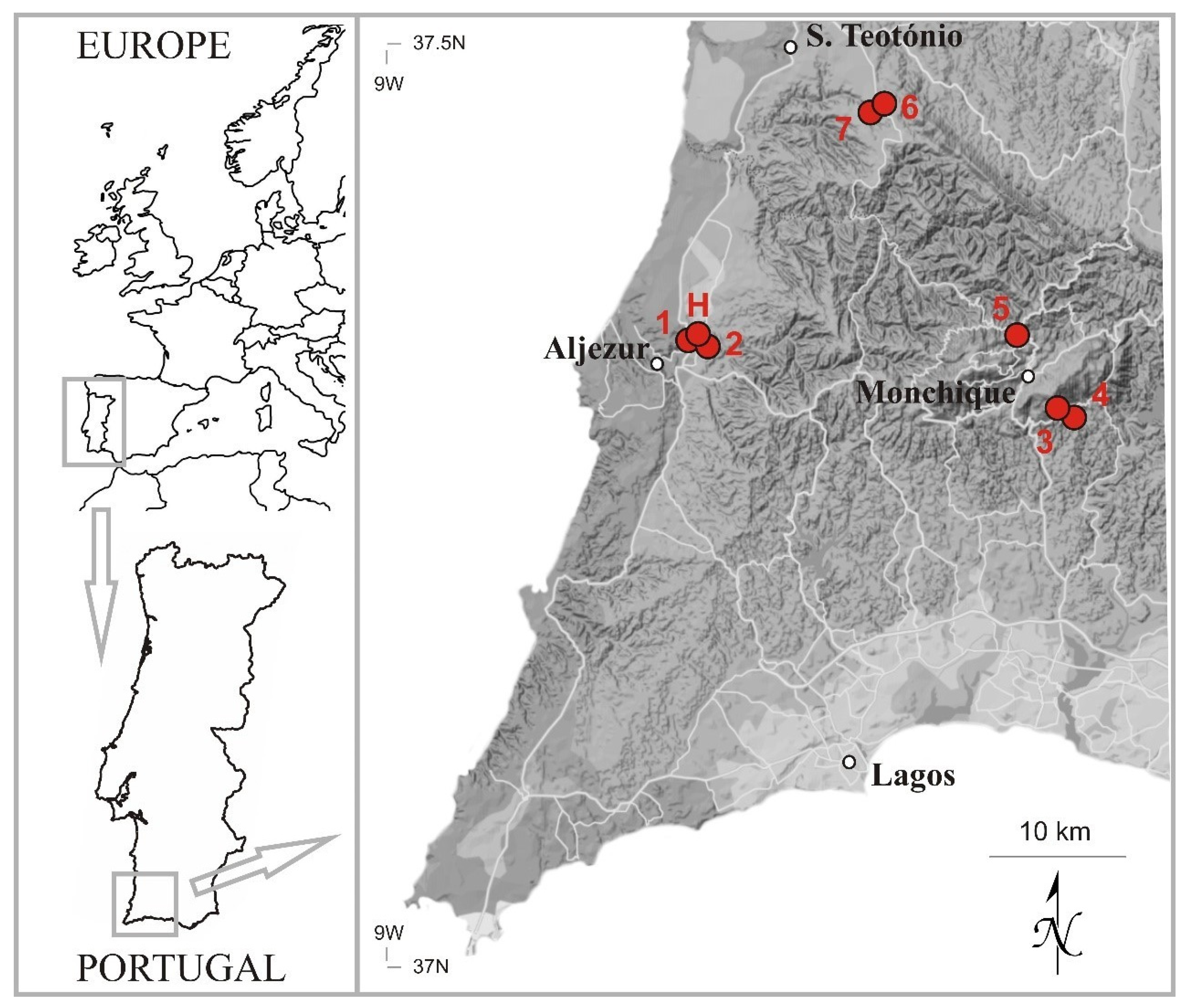{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphological Study
2.2. Molecular Phylogenetic Analysis
3. Results
3.1. Phylogenetic Results
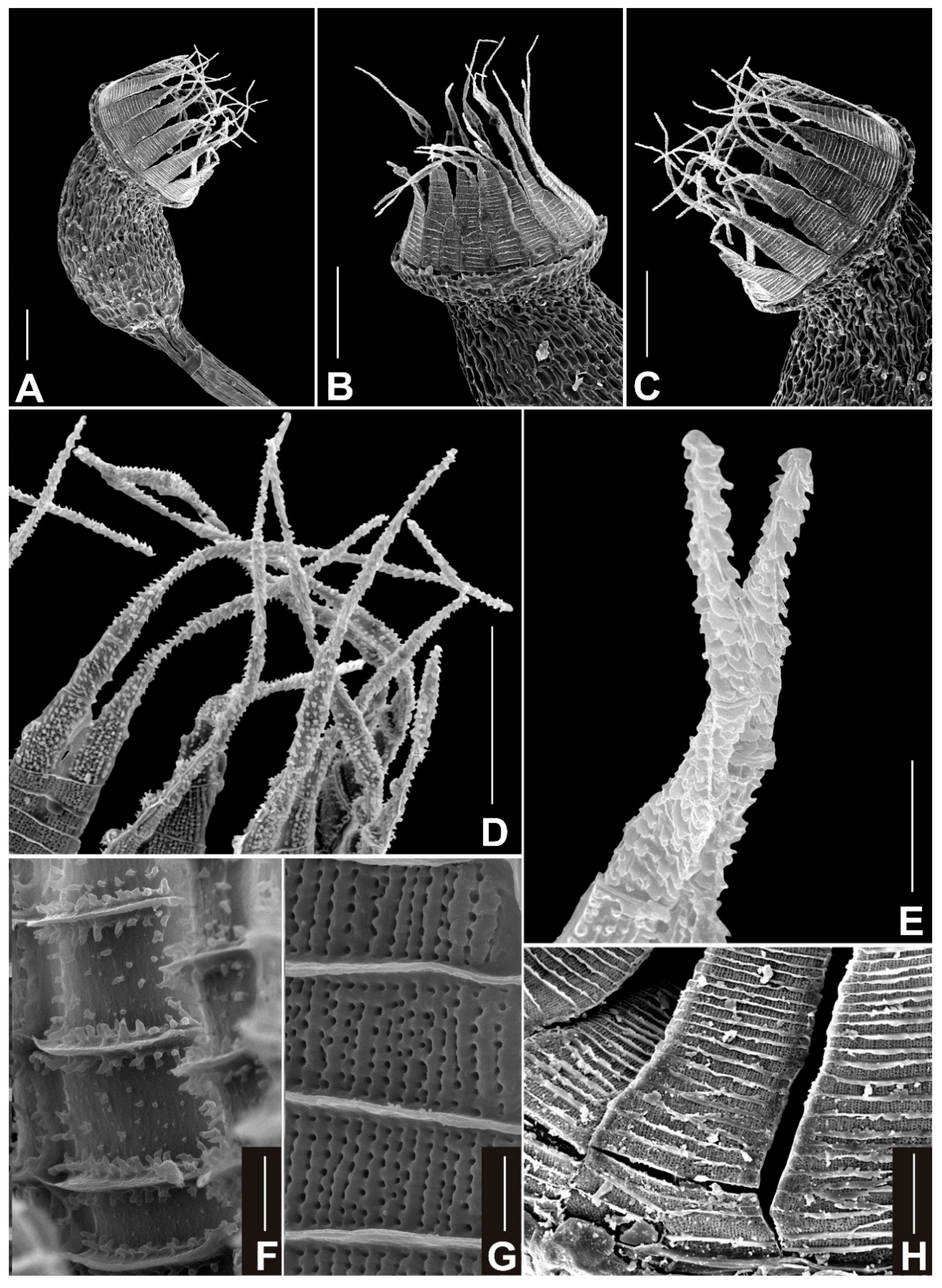
3.2. Taxonomy
4. Discussion
4.1. Taxonomical Notes
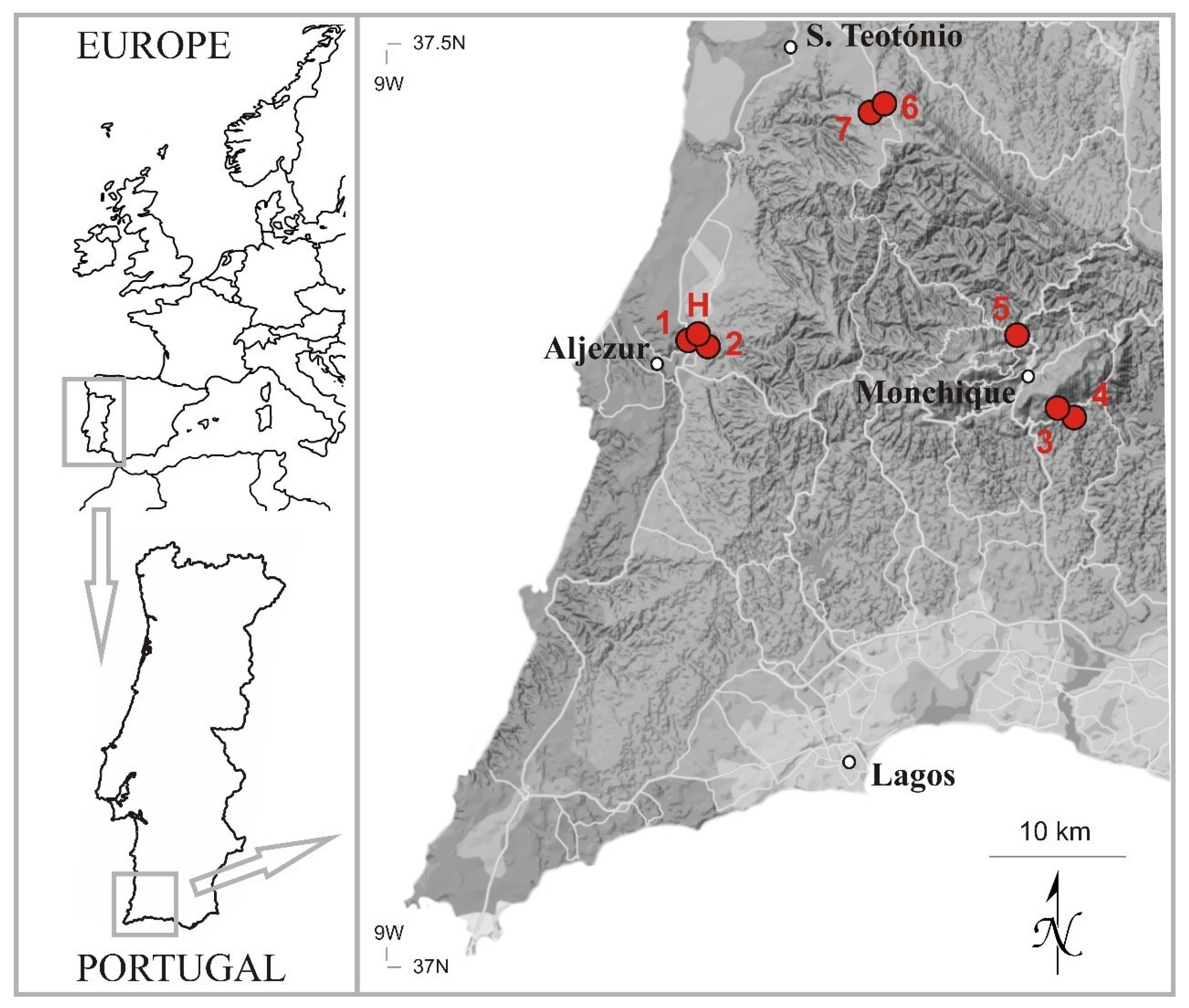
4.2. Habitat and Conservation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
References
- Frey, W.; Stech, M. Division Bryophyta Schimp. (Musci, Mosses). In Syllabus of Plant Families, Part 3: Bryophytes and Seedless Vascular Plants; Frey, W., Ed.; Gebrüder Borntraeger: Stuttgart, Germany, 2009; pp. 116–257. ISBN 9783443010638. [Google Scholar]
- Crosby, M.R.; Magill, R.E.; Allen, B.; He, S. A Checklist of the Mosses; Missouri Botanical Garden: St. Louis, MO, USA, 2000; pp. 1–320. [Google Scholar]
- Gradstein, S.R.; Churchill, S.P.; Salazar-Allen, N. Memoirs of the New York Botanical Garden. In Guide to the Bryophytes of Tropical America; New York Botanical Garden Press: New York, NY, USA, 2001; pp. 1–577. [Google Scholar]
- Larraín, J.; Suárez, G.; Bednarek-Ochyra, H.; Ochyra, R. The rediscovery of Dicranella circinata (Dicranellaceae, Bryophyta), with comments on other southern South American species of Dicranella. Nova Hedwig. 2010, 91, 361–376. [Google Scholar] [CrossRef]
- Carmo, D.M.; Peralta, D.F. Sinopse do gênero Dicranella (Müll.Hal.) Schimp. (Dicranellaceae, Bryophyta) para o Brasil com lectotipificações e citações de novas ocorrências. Pesqui. Botânica 2020, 74, 249–282. [Google Scholar]
- Ochyra, R.; Bednarek-Ochyra, H.; van Rooy, J. Dicranella hookeri, an addition to the moss flora of continental Africa. Herzogia 2013, 26, 169–174. [Google Scholar] [CrossRef]
- Bonfim Santos, M.; Fedosov, V.; Hartman, T.; Fedorova, A.; Siebel, H.; Stech, M. Phylogenetic inferences reveal deep polyphyly of Aongstroemiaceae and Dicranellaceae within the haplolepideous mosses (Dicranidae, Bryophyta). Taxon 2021, 70, 246–262. [Google Scholar] [CrossRef]
- Allen, B. Moss Flora of Central America. Part 1. Sphagnaceae-Calymperacea; Missouri Botanical Garden: St. Louis, MO, USA, 1994; pp. 1–242. ISBN 9780915279265. [Google Scholar]
- Crum, H.A. Dicranella. In Flora of North America north of Mexico. 27. Bryophytes: Mosses (1); Zander, R.H., Ed.; Oxford University Press: Oxford, UK, 2007; pp. 386–393. ISBN 9780195318234. [Google Scholar]
- Ochyra, R.; Żarnowiec, J.; Bednarek-Ochyra, H. Census Catalogue of Polish Mosses; Institute of Botany, Polish Academy of Sciences: Krakow, Poland, 2003; pp. 1–372. ISBN 83-85444-84-X. [Google Scholar]
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- Puche, F. Dicranella. In Flora Briofítica Ibérica; Guerra, J., Cano, M.J., Ros, R.M., Eds.; Universidade Murcia: Murcia, Spain, 2015; Volume 2, pp. 90–101. ISBN 9788460821984. [Google Scholar]
- Hodgetts, N.G.; Lockhart, N. Checklist and Country Status of European Bryophytes—Update 2020; Irish Wildlife Manuals, No.123; National Parks and Wildlife Service: Dublin, Ireland, 2020; pp. 1–224.
- Sérgio, C.; Garcia, C.A.; Sim-Sim, M.; Vieira, C.; Hespanhol, H.; Stow, S. Atlas and Red Data Book of Threatened Bryophytes of Portugal; Documenta: Lisbon, Portugal, 2013; pp. 1–464. ISBN 9789898618573. [Google Scholar]
- Ros, R.M.; Mazimpaka, V.; Abou-Salama, U.; Aleffi, M.; Blockeel, T.L.; Brugués, M.; Cros, R.M.; Dia, M.G.; Dirkse, G.M.; Draper, I.; et al. Mosses of the Mediterranean, an annotated checklist. Cryptogam. Bryol. 2013, 34, 99–283. [Google Scholar] [CrossRef]
- Rivas-Martínez, S.; Penas, Á.; Díaz-González, T.E.; del Río, S.; Cantó, P.; Herrero, L.; Pinto Gomes, C.; Costa, J.C. Biogeography of Spain and Portugal. Preliminary typological synopsis. Int. J. Geobot. Res. 2014, 4, 1–64. [Google Scholar] [CrossRef]
- Fedosov, V.E.; Fedorova, A.V.; Troitsky, A.V.; Bobrova, V.K.; Ignatov, M.S. On the systematic position of Hymenoloma (Bryophyta). Arctoa 2016, 25, 119–130. [Google Scholar] [CrossRef]
- Fedosov, V.E.; Fedorova, A.V.; Fedosov, A.E.; Ignatov, M.S. Phylogenetic inference and peristome evolution in haplolepideous mosses, focusing on Pseudoditrichaceae and Ditrichaceae s.l. Bot. J. Linn. Soc. 2016, 181, 139–155. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Simmons, M.P.; Ochoterena, H. Gaps as characters in sequence-based phylogenetic analyses. Syst. Biol. 2000, 49, 349–381. [Google Scholar] [CrossRef] [Green Version]
- Müller, K. SeqState. Appl. Bioinform. 2005, 4, 65–69. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. Partition-Finder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Teslenko, M.; Van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010. [Google Scholar] [CrossRef] [Green Version]
- Cox, C.J.; Goffinet, B.; Wickett, N.J.; Boles, S.B.; Shaw, A.J. Moss diversity: A molecular phylogenetic analysis of genera. Phytotaxa 2010, 9, 175–195. [Google Scholar] [CrossRef]
- Smith, A.J.E. The Moss Flora of Britain; Cambridge University Press: Cambridge, UK, 2004; pp. 1–1026. ISBN 9780521546720. [Google Scholar]
- Magill, R.E. Flora of Southern Africa. Bryophyta. Part 1, Fascicle 1 Sphagnaceae-Grimmiaceae; Botanical Research Institute: Pretoria, South Africa, 1981; pp. 120–124. ISBN 0621069515. [Google Scholar]
- Chien, G.; Crosby, M.R. Moss Flora of China. English Version, Volume 1. Sphagnaceae-Leucobryaceae; Science Press: Beijing, China, 1999; pp. 129–143. ISBN 9780915279685. [Google Scholar]
- Ochyra, R.; Lewis Smith, R.I.; Bednarek-Ochyra, H. The Illustrated Moss Flora of Antarctica; Cambridge University Press: Cambridge, UK, 2008; pp. 1–685. ISBN 987-0-521-81-402-7. [Google Scholar]
- Eddy, A. A Handbook of Malesian Mosses. Volume 1. Sphagnales to Dicranales; British Museum Natural History: London, UK, 1988; pp. 1–204. ISBN 0565010387. [Google Scholar]
- Sainsbury, G.O.K. A Handbook of the New Zealand Mosses; Royal Society of New Zealand Bulletin: Wellington, New Zealand, 1955; pp. 1–490.
- Catchside, D.G. Mosses of South Australia; D.J. Woolman: Adelaide, Australia, 1980; pp. 1–364. ISBN 0724357203. [Google Scholar]
- Malato-Beliz, J. A Serra de Monchique Flora e Vegetação; Serviço Nacional de Parques: Lisbon, Portugal, 1982; pp. 1–92.
- González-Clavijo, E.J.; Valadares, V. A Estrutura do Complexo de Monchique; Actas do VI Congresso Nacional de Geologia Ciências da Terra (UNL) No. especial V; Ciências da Terra (UNL): Lisbon, Portugal, 2003; pp. 37–40. [Google Scholar]
- Rock, N.M.S. Petrology and Petrogenesis of the Monchique Alkaline Complex. J. Petrol. 1978, 19, 171–214. [Google Scholar] [CrossRef]
- Sérgio, C.; Figueira, R.; Menezes, R. Modelling the distribution of Sematophyllum substrumulosum (Hampe) E. Britton as a signal of climate changes in Europe. In Bryophyte Ecology and Climate Change; Tuba, Z., Slack, N.G., Stark, L.R., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 427–439. ISBN 9780521757775. [Google Scholar]
- EEA. European Forest Types. Categories and Types for Sustainable Forest Management Reporting and Policy; EEA Technical Report No. 9/2006; EEA: Copenhagen, Denmark, 2007; Available online: http://www.eea.europa.eu/publications/technical_report_2006_9 (accessed on 6 September 2021).
- Mabberley, D.J.; Placito, P.J. Algarve Plants and Landscapes: Passing Tradition and Ecological Change; Oxford University Press: Oxford, UK, 1993; pp. 1–336. ISBN 0198587023. [Google Scholar]
- De Rios, R.S.; Benito-Garzón, M.; Sainz-Ollero, H. Present and future extension of the Iberian submediterranean territories as determined from the distribution of marcescent oaks. Plant Ecol. 2009, 204, 189–205. [Google Scholar]
- Sousa, P.M.; Trigo, R.M.; Pereira, M.G.; Bedia, J.; Gutiérrez, J.M. Different approaches to model future burnt area in the Iberian Peninsula. Agric. For. Meterol. 2015, 202, 11–25. [Google Scholar] [CrossRef]
- Hewitt, G.M. Post-Glacial re-colonization of European biota. Bot. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Gómez, A.; Lunt, D.H. Refugia within refugia: Patterns of phylogeographic concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia; Weiss, S., Ferrand, N., Eds.; Springer: Amsterdam, The Netherlands, 2006; pp. 155–188. [Google Scholar] [CrossRef]
- Olalde, M.; Herrán, A.; Espinel, S.; Goicoechea, P.G. White oaks phylogeography in the Iberian Peninsula. For. Ecol. Manag. 2002, 156, 89–102. [Google Scholar] [CrossRef] [Green Version]
- Patiño, J.; Vanderpoorten, A. Bryophyte biogeography. Crit. Rev. Plant Sci. 2018, 2–3, 175–209. [Google Scholar] [CrossRef]
- Albertos, B.; Garilleti, R.; Heras, P.; Infante, M. On the mend of bryophyte conservation in Spain: Preparing a proposal for the inclusion of bryophytes in national species protection catalogues. Mediterr. Bot. 2018, 39, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Ignatov, M.S.; Porley, R.D.; Ochyra, R.; Kuznetsova, O.I.; Ignatova, E.A. Coscinodon monchiquensis R.D.Porley, Ochyra & Ignatova (Grimmiaceae), a new species from the Algarve, southern Portugal. J. Bryol. 2018, 40, 125–136. [Google Scholar]
- Infante, M.; Muñoz Puelles, L.; Albertos, B.; Garilleti, R.; Heras, P. View on bryophyte conservation in Peninsular and Balearic Spain: Analysis of Red Lists and legal protection. Cryptogam. Bryol. 2017, 38, 19–51. [Google Scholar] [CrossRef]
- Vila-Viçosa, C.; Gonçalves, J.; Honrado, J.; Lomba, Â.; Almeida, R.R.; Vázquez, F.M.; Garcia, C. Late Quaternary range shifts of marcescent oaks unveil the dynamics of a major biogeographic transition in southern Europe. Sci. Rep. 2020, 10, 21598. [Google Scholar] [CrossRef]
- Vila-Viçosa, C.; Vázquez, F.M.; Mendes, P.; Del Rio, S.; Musarella, C.; Cano-Ortiz, A.; Meireles, C. Syntaxonomic update on the relict groves of Mirbeck’s oak (Quercus canariensis Willd. and Q. marianica C. Vicioso) in southern Iberia. Plant Biosyst. 2015, 149, 512–526. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Porley, R.D.; Fedosov, V.; Plášek, V.; Fedorova, A. Undiscovered Biodiversity of the European Moss Flora: Neodicranella hamulosa (Aongstroemiaceae), a New Genus and Species from SW Portugal. Plants 2021, 10, 2289. https://doi.org/10.3390/plants10112289
Porley RD, Fedosov V, Plášek V, Fedorova A. Undiscovered Biodiversity of the European Moss Flora: Neodicranella hamulosa (Aongstroemiaceae), a New Genus and Species from SW Portugal. Plants. 2021; 10(11):2289. https://doi.org/10.3390/plants10112289
Chicago/Turabian StylePorley, Ronald D., Vladimir Fedosov, Vítězslav Plášek, and Alina Fedorova. 2021. "Undiscovered Biodiversity of the European Moss Flora: Neodicranella hamulosa (Aongstroemiaceae), a New Genus and Species from SW Portugal" Plants 10, no. 11: 2289. https://doi.org/10.3390/plants10112289
APA StylePorley, R. D., Fedosov, V., Plášek, V., & Fedorova, A. (2021). Undiscovered Biodiversity of the European Moss Flora: Neodicranella hamulosa (Aongstroemiaceae), a New Genus and Species from SW Portugal. Plants, 10(11), 2289. https://doi.org/10.3390/plants10112289